Island Biogeography: Patterns in Species Richness
Wallace, Darwin, Hooker
David Lack (1947, 1976) -- critical study of evolution and ecology of
Galápagos finches
Ernst Mayr (1942, 1963) -- followed Wallace’s footsteps in the East Indies
G. Evelyn Hutchinson (1958,1959, 1967) -- studied lakes
Prior to 1960, dominant idea was the static theory of islands: insular
community structure was fixed in ecological time
Robert MacArthur and E. O. Wilson (1963, 1967) -- equilibrium theory of
island biogeography; dynamic equilibrium
The major contribution to biogeography was the theoretical aspect of
modeling
Searched for general patterns in the distributions of diverse kinds of
species
General themes of equilibrium theory of island biogeography: (1) the
tendency for species numbers to increase with area; (2) tendency for
species numbers to decrease with isolation; immigration and extinction
are relatively frequent -- so numbers don’t necessarily change, but
species composition does.
Other important contributors to island biogeography:
Eugene G. Munroe (1948) -- developed the equilibrium theory before
MacArthur and Wilson, but ideas buried in doctoral thesis about Caribbean
butterflies and never published
Frank Preston (1962) -- One of his contributions was the idea that in any
region, only a few species are extremely common, and most are moderately or
very rare. He also pointed out that small, isolated islands have fewer species
that are more prone to extinction because of their rarity.
Island Patterns that were used in MacArthur and Wilson’s
theory
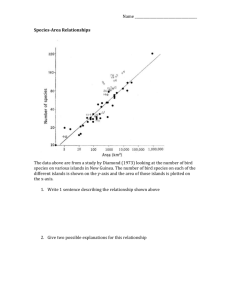
Species-Area Relationship
7-1
species numbers tends to increase with increasing area
this relationship described mathematically by Arrhenius (1920) as
S = cAz
where S = species number
A = island area
c = fitted constant that depends on the taxon and biogeographic region; but
most strongly on population density
z = fitted parameter that represents the slope when both S and A are plotted
on logarithmic scales; changes very little among taxa or within a given
taxon in different parts of the world, but is correlated with elevation
and proximity to the mainland
log (S) = log (c) + z log(A)
referred to as the “power model”
Species-isolation relationship
For a variety of taxa and ecosystems, species richness should decline as a
negative exponential (sigmoidal function) of isolation
Species Turnover
number of species increases until saturation; spp go extinct and are replaced
by others
Equilibrium Theory of Island Biogeography
Three basic characteristics:
1. species-area relationship
2. species-isolation relationship
3. species turnover
7-2
number of species inhabiting an island represents a dynamic equilibrium
between opposing rates of immigration and extinction, which are recurrent.
(Fig. 13.7) - simplest model
The simplest model doesn’t accommodate island size or isolation (Fig. 13.8) - in this model, island size affects only extinction; isolation affects the
immigration rate
Bottom line:
SLN > SLF ~ SSN > SSF
where L = large, S = small, N = near, F = far
Model also predicts this relationship among turnover rates:
TSN > TSF ~ TLN > TLF
Strengths and Weaknesses of Equilibrium Theory
Strength of the theory lies in the simplicity of the model -- specifically that
the model offered clear, testable predictions
Weakness also lies in the simplicity of the model -- and these were pointed
out by the authors
lack of information about the precise shape of the extinction and
immigration curves
7-3
artificial distinction between immigration and extinction
assumption that extinction and immigration curves have fairly regular
shapes for different faunas and different islands and for different
times on the same island
Concerns raised by others:
many insular biotas may not be in equilibrium
the model ignores interspecific interactions
immigration and extinction are treated as independent processes
it may be difficult to identify the source of an island biota
speciation on islands violate the model
area provides a coarse measure of an island’s ability to support a biota
Tests of the model
Estimates of turnover on landbridge islands
Jared Diamond (1969) - examined turnover of birds on California Channel
Islands
A follow-up study by Jones and Diamond (1976) where Santa Catalina island
censused several years in succession found year-to-year turnover.
Others looked at other island systems and found human influence to have
highest impact on species turnover.
In other words, the support is not yet compelling
7-4
Turnover on recently created anthropogenic islands
archipelagos created by flooding of mountainous areas in the tropics:
Barro Colorado island in Lake Gatun; islands in Lago Guri. Surveys of both
systems reveal that extinctions and immigrations appear to be recurrent.
Thomas Schoener (1983) reviewed 21 studies of turnover -- revealed that
turnover tends to be lower on larger islands and increases with generation
time of the organisms
= Support for the theory
Experimental defaunation
Wilson and Simberloff (1969, 1970) -- worked with mangrove islands in the
Florida Keys. Did initial survey of arthropods, hired an exterminator to
come in an kill all animals using methyl bromide (no harm to plants), and
then monitored the islands.
Recolonization was rapid. High rate of turnover noted, also.
= strong support for the theory
Other Patterns of Island Species Richness
The rescue effect
Study of insects and spiders growing on thistles plants growing in desert
shrubland of southeastern Arizona (Brown and Kodric-Brown 1977).
Proximity to a source of immigrants increases the immigration rate, and a
continual influx of individuals within a species tends to prevent extinction.
Rescue effect: high rates of immigration reduce the probability that a
species will temporarily be absent and hence recorded as a turnover.
Target area effect
larger islands may serve as a more effective target areas for potential
immigrants because they are more likely to be seen or encountered.
Small island effect
tendency of species richness of some insular faunas to remain relatively low
and independent of areas for the smallest islands
7-5
Insular biotas that are not in equilibrium
species numbers increasing or decreasing as a result of historical
perturbation -- approaching a new equilibrium, rather than being in
equilibrium
Pleistocene refugia
Habitat refugia
Ecologists have attempted to apply the equilibrium theory to the design of
nature reserves. Habitat fragmentation has a huge impact on species-area
curves, immigration and extinction.
Krakatau
Observation of Rakata have supported much of the equilibrium theory but
also highlighted limitations of the theory.
Other Limitations of Island Biogeography Theory:
MacArthur and Wilson anticipated that immigration and extinction curves
may not be simple if major succession changes occur. Some species are
strongly dependent on other pioneer species to create niches for them.
Periodic Disturbance
7-6
Equilibrium may never be attained if ecosystems are constantly disturbed
(hurricanes, fire, volcanic eruptions, etc).
Speciation
equilibrium model does not accommodate speciation.
7-7
Patterns in the Assembly and Evolution
of Insular Communities
Islands tend to be species-poor in comparison to mainland communities
Structure of insular communities is strongly influenced by evolution in situ
Harmonic biota - where island biotas are similar to source biota
Disharmonic biota - biotas have major differences
Selective nature of immigration
species vary in traits affecting their immigration potential
to the extent that interspecific differences are predictable, they will translate
into predictable, nonrandom patterns of species distributions and
community structure among islands.
Establishing Insular Populations
Good dispersers may be preadapted for successful colonization of islands
r- strategists vs. K-strategists
Selective nature of extinction
islands should favor communities of good dispersers that require few
resources.
some types of species have become locally extinct much more frequently than
7-8
others; differences related to ecological characteristics
Animals of larger size, carnivorous diet, or specialized habitat
requirements more susceptible to extinction than species that are
smaller, herbivorous, and generalized in habitat requirements
body size and demography influence the ability of introduced birds to
establish and maintain insular populations
Extinction of Hawaiian birds influenced by habitat preference
Patterns reflecting differential immigration
and extinction
Nestedness of insular communities
Islands of archipelagos may have decreasing subsets of taxa as isolation
increases
Darlington’s (1957) graphical model for nestedness
Distributions of particular species
Metapopulation theory -- an attempt to estimate the proportion of islands
(patches) that must be occupied to ensure the survival of the interacting
populations of a species - its metapopulation.
metapopulation models can be used to estimate the time to extinction
probability or frequency of immigration decreases with increasing
isolation
7-9
frequency of extinction decreases as island area increases
minimal area requirements to maintain populations should increase as
isolation increases
Patterns reflecting interspecific interactions
competition*, predation, parasitism, mutualism
Predicted patterns:
1. ecologically similar species should exhibit mutually exclusive
distributions, seldom if ever occurring together on the same island
2. conspecific populations on species-poor islands should exhibit
relatively high densities
3. ecological release should be exhibited; characterized by significantly
broader niches and shifts to other niches
Some patterns of insular community structure appear to be so regular that
Diamond termed them assembly rules
Evolutionary Trends on Islands
Flightlessness and reduced dispersal ability on islands
Birds:
derived flightlessness appears in eight orders of birds and most
archipelagoes
selective pressures associated with absence of predators and limited resources
on islands -- ecological release
moas, dodoes, solitaires, elephant birds
7-10
Insects:
beetles, lepidopterans, flies, bees, orthoptera, homoptera
some insects have lost power of flight and increased in size and have taken
over the rodent niche in New Zealand (the wetas)
selection pressure to remain in place
Snails:
small body size enables dispersal by wind; colonization by microsnails
followed by evolution of large body size
Plants:
colonizers were good dispersers (some via wind); reduction in dispersability
noted
herbaceous forms on mainland evolve to secondary woodiness (tree forms
common) -- represents ecological release
Evolution of Body Size on Islands
Gigantism:
Ecological release can promote gigantism.
Large size has a number of advantages:
1.
Larger individuals can exploit a greater diversity of resources.
2.
With sufficient resources, larger individuals can produce larger litters
or clutches.
3.
Larger individuals dominate in territorial disputes or in other
competitive interactions.
4.
Larger individuals have relatively greater energy and water reserves
7-11
that enable them to withstand famine and drought better than smaller
individuals.
Dwarfism:
Resource limitation can promote dwarfism.
Being small also has certain selective advantages:
1.
Smaller individuals need fewer resources.
2.
Smaller individuals tend to be more specialized and more efficient and
assimilating nutrients and energy.
3.
Smaller individuals can exploit small shelters and refugia from
predators -- reduce impact of stressful environments.
Insular mammals:
Island Rule sensu J. Bristol Foster (1964): Different groups of mammals
tend to exhibit different trends in insular body size.
General trend -- dwarfism in large mainland taxa and gigantism in small
mainland taxa
Pygmy mammoth: once widespread across Malaysia, the
Mediterranean, the California Channel and the Arctic Ocean.
Birds and reptiles:
Widely distributed on islands. Different families and orders exhibit different
trends
Gigantism common in iguanids, herbivorous lizards, whiptails, tiger snakes.
Dwarfism common in rattlesnakes, rails, ducks
7-12
7-13
The Taxon Cycle
E. O. Wilson (1959, 1961) proposed that insular species evolve through a
series of stages from newly arrived colonists (mainland forms) to highly
differentiated endemics, which ultimately go extinct.
Stage I -- initial colonization and establishment
Stage II -- differentiation of insular populations
Stage III -- differentiation and range contraction continues, taxon may
represent relictual endemic, highly specialized.
Stage IV -- ranges of relictual endemic undergo further contraction; extreme
specialization results in weak competitiveness to new species colonizing
island; extinction.
7-14
 0
0