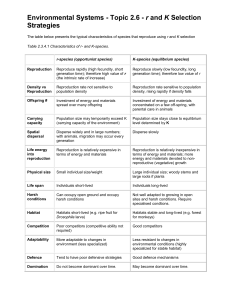
Lesson 6: Allocation and life history patterns
advertisement

Lesson 6: Allocation and life history patterns Principal of allocation Allocation to resource acquisition Allocation to survival Allocation to reproduction and reproductive structures • Plant strategies and life history patterns Seed dispersal and dormancy Classification of life history patterns r- and k-selected species (McArthur and Wilson) R-, C-, and S-selected species (Grime) Allocation and Strategy • Principle of Allocation: Individual plants have a limited amount of resources to spend on – growth, – maintenance (survival), – reproduction. • Strategy (or life history pattern) is a genetically inherited pattern of resource allocation to these basic functions that has evolved through the process of natural selection. Allocation: analogy with principles of economic theory “Plant growth has been considered analogous to a business operating under the principles of economic theory. Individuals that have the greatest profit (grow fastest) gain an advantage (usurp more space and increase their resource base). However, allocation to growth is typically associated with trade-offs (e.g., decreased reproduction). In other words, growth, reproduction, and survival represent competing demands for a finite supply of carbohydrates, fats or waxes, and proteins produced by synthetic processes in the plant. Thus, we could view the acquisition of resources as income, with expenditures partitioned between maintenance (e.g., storage and defenses), growth, and reproduction. Allocation can then be treated by a cost/benefit analysis, allowing the prediction of optimal allocation patterns. The ultimate measure of successful allocation, however, is reproductive success.” Barbour et al., 1999, p. 89. Three examples of plant strategies (a) Even allocation to reproduction, growth and maintenance throughout the growth period (probably no plant has this pattern). (b)Pattern of a typical annual. Most resources are allocated to growth in the early part of the growing season. Late in the growing season, nearly all the resources are allocated to reproduction. (c ) Pattern of a typical stress-tolerant tree or shrub. Most resources are allocated to maintenance , less to growth. They allocate resources to reproduction in growing seasons that are high in resource availability. A lot of tundra or desert plants have this sort of allocation pattern. Wilson 1983 cited in Barbour et al. 1999. Trade-offs in resource allocation 1. Allocating resources to the development of some plant modules (roots, shoots, seeds) restricts the ability of plant to develop others. 2. Allocation for the capture of some resources, (e.g. root production for capture of nutrients or water) will necessarily inhibit allocation to structures to capture other resources (e.g. leaves for carbon acquisition). Trade-offs associated with allocation in Poa pratensis (Kentucky bluegrass) Inflorescences in year 1 vs. Inflorescences in Year 2 Inflorescences in year 1 vs. Size of plant in Year 2 • High levels of reproduction in first year correspond to fewer flowers and smaller plants in second year. • Allocating more nutrients to reproductive organs in the first year results in less available nutrients in the second year. Law. 1979. The cost of reproduction effort in clonal plants: a benefit-cost model. Oikos 49: 199-208.) Problems associated with economic analogy in cost/benefit analysis of plant allocation 1. There are three currencies associated with plant growth (versus one in standard economic theory): carbon (CO2 acquisition using solar energy via photosynthesis and stored in organic molecules), water, and soil nutrients. 2. The net cost of biomass production is difficult to access. For example, reproductive cost of flowers may not consider that some flowers have photosynthetic parts that help offset the cost of flower making. 3. The costs of plasticity are difficult to assess. For example, nutrients are often resorbed from leaves and flowers before abscission, making the true costs difficult to measure. Justis Liebig Law of the minimum • The growth or distribution of a plant is dependent on the one environmental factor most critically in demand. Concept of limiting resources applied to competing species (Armstrong and McGeehee (1980) • m is the mortality rate (rate of population growth below which mortality of a species occurs). • R* is the level of resource R, below which a species cannot maintain itself. • For species A and B, B is able to maintain itself at a lower level of Resource R and hence is considered the better competitor. Population growth curves for Species A and B Tilman 1982 cited in Barbour 1999. An unusual aspect of plants with regard to competition for resources • The structures responsible for acquiring essential resources (water, energy, and nutrients) are in different parts of the plants one above ground and one below ground. Energy acquisition is done in the leaves through photosynthesis. Water and nutrients are acquired mainly through the roots. • Plants must balance allocating resources towards roots (water and nutrients) and shoots (energy, photosynthesis, and carbon gain). • Root:shoot ratios often give insights to the allocation strategies of plants. Example of very different aboveground and belowground environments for resource acquisition • • • Ranunculus adoneus, a snowbed plant in the Colorado alpine, can experience very hot aboveground conditions and simultaneous very cold soils conditions (from snow meltwater) Mismatch between the plants ability to produce phytosynthate in the leaves, and its ability to absorb the essential water and nutrients in the roots to support this production. So in this case, the plant may have to allocate more resources to belowground roots. Resource-ratio hypothesis (Huston and Smith 1987, Tilman 1988) Differences in the relative supply rates of limiting resources should lead to differences in the composition of plant communities: – Species allocation patterns: Species with allocation patterns focusing on shoots are assumed to be relatively effective competitors for light, and those allocating more heavily to roots are assumed to be good competitor for below-ground resources (water, nutrients). – Landscape implications: Various habitats within landscapes differ in their level of key resources, and hence will favor either root or shoot specialists depending on the local resource supply. – Succession implications: Resource supply ratios also vary systematically through successional series to first favor root specialists (because soil nutrition is more limiting than light in primary succession) and then shoot specialists because light is more limiting in later stages of succession. Examples of different allocation mechanisms for survival Life-history patterns Length of life Age at reproduction Differing growth rates Plant size Patterns of reproduction Monocarpic (semelparous) vs. Polycarpic (iteroparous) reproduction Seasonal timing of reproduction (early vs. late summer) Seed size Dispersal mechanisms Defense mechanisms Toleration of herbivory (increased nutrient uptake or photosynthesis) vs. chemical defenses Defensive plant structures (spines, hairs, tough leaves) Life-history patterns How could each of these factors affect a plants ability to survive? • • • • Length of life (annual, biennial, perennials) Age at reproduction Differing growth rates Plant size Allocation to maintenance: High root-shoot ratios • Courtesy of Niwot LTER web site Plants in highly stressful environments such as the alpine or arctic often have high root to shoot ratios. Other plants that allocate lots of resources to maintenance Sequoiadendron giganteum (Giant Sequoia) www.ginnyprior.com • Long-lived woody plants • Plants with a lot of protective chemical compounds and unpalatable leaves. Empetrum nigrum (Crowberry) www.borealforest.org/ shrubs/crowberry.jpg Plants with high reproductive strategies • Many annuals and biennials (ruderal species) in weedy environments; produce abundant early germinating seeds; palatable leaves; fast growing; need lots of light; low root to shoot ratios. • Desert annuals, plants able to take advantage of short moist periods with rapid growth, flowerin, and seed set. Can endure long periods of time in dormant state as seeds. Alliaria petiolata, garlic mustard www.uwgb.edu/.../allpet_ seedlings01_tnail.jpg Papaver escholtziii http://dplu-mscp.sdcounty.ca.gov/speciesinfo_6/bio13desert_scrubportal.htm Monocarpic species Frasera speciosa (green gentian, monument plant) Yucca whipplei (Yucca) www.laspilitas.com/ plants/pictures/a1077.jpg www-plb.ucdavis.edu/.../ Yucca_whipplei.jp Polycarpic vs. monocarpic reproduction • Polycarpic reproduction (plants that reproduce more than once) – Favored when adult mortality is low. – Potential to outproduce monocarpic species over a long lifetime. • Monocarpic reproduction (plants that reproduce once and then die) – Favored when adult mortality rates are high. – A huge burst of flowering allows the individual to gain a greater share of pollinators in a competitive system (e.g. Agave and Yucca). – Huge seed output can also be good strategy to coop space in open habitats. Many ways to allocate to reproduction: Each has its tradeoffs • Sexual reproduction – High investment in seed production and/or fertilization • High pollen production vs. large ovules (male vs. female fitness) • High pollen numbers vs. large pollen size (wind vs. animal dissemination) • Gamete production vs. pollinator or disperser reward (ovule number vs. nectar or food reward) – Investment in potential seedling success • Seed size (provisioning) vs. seed number • Dispersal vs. provisioning (e.g., wings, or bristles, downy appendages vs. fruit quality or size) • Vegetative reproduction (sprouting vs. seed production) Acacia seeds adapted for ant and bird dispersal • • ODowd & Gill 1986, cited in Barbour et al. 1999 Acacia seeds apparently adapted for dispersal by ants (above) and birds (below). The food reward in the appendage (eliaosome) on the seed is larger and usually more colorful (red, orange, or yellow vs. white) in the bird dispersed species. Example of poor reproductive resource allocation • Idiot fruit, Idiospermum australiensis. • Ancient 100 million year-old species. • Puts all reproductive resources into a large toxic fruit that now is impossible to disperse. • Toxic to all animals except an extinct species of bird. • Highly endemic to a very small area of the Daintree Rainforest in NE Australia. Life history patterns: Longevity vs. age at first reproduction and size of plant Longevity vs. age at first reproduction Size vs. longevity Loehle 1988, cited in Barbour et al. 1999 • Correlation between typical age of sexual maturity versus typical longevity for North American angiosperm trees and shrubs. Early investment in growth and maintenance has cost to early reproduction. • The longer an adult plant is likely to survive, the more likely it is to postpone reproduction in favor of production of woody and other maintenance costs. (greater woodiness of longer-lived plants). Trees with longer life spans also produce wood that is more likely to be better defended against decay and disease. Classification of Life History Attributes Why? • • • • Evolution leads to adaptations that maximize “fitness”. Life history classification schemes aid in our effort to understand and communicate information about plant adaptations. Plants growing in similar spatial and temporal microhabitats often have similar life history patterns. There have been many approaches to classify life-history patterns. Some focus on single aspects of life history such as reproduction, maintenance, or growth. r- and K-selected Life History Patterns (MacArthur and Wilson 1967) MacArthur and Wilson (1967) in their analysis of island biogeography, developed the concept of placing organisms within a spectrum between extremes of allocation to reproduction. • r- selected plants are those that maximize their intrinsic rate of reproductive increase. This is done through high seed production, and minimizing costs for maintenance. They generally grow in highly unpredictable climates or habitats, have low long term survivorship, are poor competitors, short development times, short life-span, strong reproductive focus with a monocarpic reproductive effort. • K-selected species tend to grow in more predictable climates. Mortality is density dependent, They have Type I or II survivorship. The population is near constant, near the carrying capacity. They are strongly affected by competition. They have a long development time, long life span, and generally a small seed bank. Allocation is to survivorship and delayed reproduction, which is typically polycarpic. Most plants are somewhere between these extremes Survivorship types Type I: characteristic of organisms with mortality concentrated in the later stages of life (e.g., annuals with seed dormancy). Type II: characteristic of organisms with constant mortality rates. Type III: characteristic of organisms with high juvenile mortality (e.g., most trees). Summary of r- and k-selected adaptations for different traits Pianka 1970, cited in Barbour Example of how variation in allocation to biomass affects on longevity of two desert ephemerals Camissonia boothii (Booths evening primrose) Onagraceae Clark & Burk 1980, cited in Barbour et al. 1999 Plantago ovata (Desert Indianwheat) Polygonaceae Photos courtesy of USDA Plants Database Comparison of life history patterns in two desert ephemerals • Early in the season, Camissonia allocates relatively more biomass to maintenance in the form of abovground and belowground biomass. • Plantago allocates more to reproductive structures. • Camissonia has over twice the life span because it has allocated more biomass to storage and vegetative tissues. • Plantago has higher short-term reproducutive output because of early, heavy commitment of resources to reproduction. • Comparatively, Plantago is more of an rselected species, and presumably will have high reproductive output even in years where there is no late season rainfall. Whereas, Camissonia should realize more reproduction in years with more late-season rainfall and an extended growing season. • Both plants can survive in the same habitat because of high year-to-year variability in the timing of rainfall. C-, S-, and R-, selected species (J.P. Grime 1977) Grime described a third strategy. He essentially split the k-selected species of McArthur and Wilson into two groups C- and S-selected species, and developed three groups, each specializing in one of the three main growth strategies: reproduction, growth and maintenance. • Competitors (C ), plants that allocate most available resources to growth and that can capitalize on readily available resources. They have high efficiency for capturing resources that makes it difficult for other species to compete. • Stress-tolerators (S) are plants that live in habitats where resources are limited, or where survival depends on allocating most resources to maintenance and defense, (e.g., deserts, arctic tundra, alpine meadows, peat bogs, serpentine soils). • Ruderals (R) (weedy species) are typically found in temporary, frequently disturbed, habitats and are characterized by allocating most resources to reproduction, and an adaptive suite identical to that described for r-selected plants by MacArthur and Wilson. Examples of R-selected species Epilobium latifolium E. angustifolium • Soft, deciduous leaves,that are poorly defended against insects. • Produce many flowers. Both produce many light seeds that are easily dispersed by wind. • Generally short-lived plants that are replaced by other species during succession. Examples of S-selected species Ledum decumbens Picea mariana Rhizocarpon geographicum • Long-lived, slow-growing plants. Often woody. Many lichens. • Live in sites with low-levels of nutrients. • Relatively coarse, evergreen leaves or non deciduous structures. • High levels of tannins and other secondary metabolites. Highly defended against insect attack. • Tend to have years of high flowering and years of low flowering. Examples of C-selected species Vicia cracca (blue vetch) www.bruehlmeier.info/ Bilder/1201%20Vicia%20cr • • • • Poa pratensis turfgrassmanagement.psu.edu/ Graphics/kblue.jpg Allocate most available resources to growth. Highly competive root systems (offten rhizomatous species). Strong competitors for nitrogen. Or are nitrogen fixers. High efficiency for capturing resources (water, light, nutrients) that makes it difficult for other species to compete. Summary of C-, S-, and R-, selected characteristics Grimes triangular model of plant characteristics Allows for intermediate combinations of the R-, C-, and S-characteristics. The corners of the triangle represent the extremes of R, S, and C species. Interior positions in the triangle represent different combinations of the characteristics. (b) shows the position of several growth forms. Annuals (A), biennials (B), lichens (L), trees (T) and shrubs (S). (c )The position of bryophytes (BR) and perennials (P). Grime 1990 Summary • • • • Principle of Allocation: Individual plants have a limited amount resources to spend on growth, maintenance (survival), and reproduction and adopt strategies to allocate resources. Strategy (or life history pattern) is a genetically inherited pattern of resource allocation that has evolved through the process of natural selection. There are trade-offs associated with different allocation patterns. Tilmans Resource-ratio Hypothesis is a cost/benefit analysis of different patterns of plant allocation. It focuses on plant competition for limited resources (more on this later). Plants have adapted to survival in different habitats by differing allocation patterns in terms of: • Patterns of reproduction – • Life-history patterns – • Length of life, age at reproduction, differing growth rates, plant size, seed size Defense mechanisms – • Moncarpic (semelparous) vs. Polycarpic (iteroparous) reproduction, seasonal timing of reproduction (early vs. late summer) Toleration of herbivory (increased nutrient uptake or photosynthesis) vs. chemical defenses, also defensive plant structures (spines, hairs, tough leaves) Grimes plant strategy classification includes R-, C-, and S-selected species. It focuses on the role of environment in relation to plant life histories. Literature for Lesson 7 **Grime, J.P. Evidence for the existence of three primary strategies in plants and relevance to ecological and evolutionary theory. American Naturalist 111: 1169-1194. • Rejmanek, M. and D.M. Richardson. 1996. What attributes make some plant species more invasive? Ecology 77: 1655-1661. • Reichard, S.H. and C.W. Hamilton. 1997. Predicting invasions of woody plants introduced into North America. Conservation Biology 11: 193-203. **Bloom, A.J. F.S. Chapin, III., H.A. Mooney 1985. Resource limitation in plants – An Economic analogy. Ann. Rev. Ecol. Syst., 16: 393. **Schulze, E.D. and F.S. Chapin, III. 1987. Plant specialization to environments of different resource availabilities. Ecol. Studies, 61: 120.. • Chapin, F.S., III, E. Schulze, and H.A. Money. 1990. Storage ecology and economics of storage in plants. Ann. Rev. Ecol. Syst. 21:423-447. • Chapin, F.S., III. 1991. Integrated responses of plants to stress: a centralized system of physiological responses. BioScience 41: 29-36. • McGraw, J.B. 1985. Experimental ecology of Dryas octopetala ecotypes: relative response to competitors. New Phytologist, 100: 223-241.