LATICIFERS OF FICUS CARICA AND THEIR POTENTIAL ROLE IN
advertisement

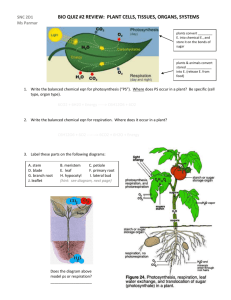
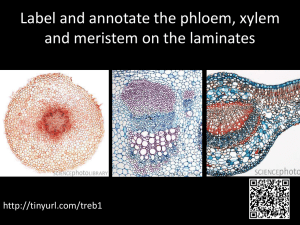
IAWA Kajii Journal et al. –35Laticifers (2), 2014:of109–115 Ficus 109 LATICIFERS OF FICUS CARICA AND THEIR POTENTIAL ROLE IN PLANT DEFENSE C. Kajii 1, T. Morita 2 and K. Kuroda 1, * 1 Graduate School of Agriculture, Kobe University, 1-1 Rokkodai, Nada-ku, Kobe 657-8501, Japan Technology Research Center, Hiroshima Prefectural Technology Research Institute, 2835 Mitsu, Akitsu-cho, Higashihiroshima 739-2402, Japan *Corresponding author; e-mail: kurodak@garnet.kobe-u.ac.jp 2Agricultural ABSTRACT The distribution and structure of laticifers in Ficus carica L. were investigated in a current-year branch and an 8-year-old trunk, using serial sections stained with safranin-fast green or nile blue. In the 8-year-old trunk, laticifers were found not only in the cortex, secondary phloem, and pith, but also in the secondary xylem. The laticifers in the phloem and xylem were of the branched, non-articulated type. In addition, horizontal laticifers extending from the phloem through the cambium to the xylem were found in some rays. Laticifers penetrating the cambial initial layer elongated in the cambial zone without cell division. Activation of latex production occurred after wounding of cortex and phloem, and the latex exuded from the cut site of laticifers into the surrounding wounded tissue and broken cells. The potential role of laticifers in defense of F. carica against certain pathogens is discussed. Keywords: Latex, fig tree, defense, Ceratocystis ficicola. INTRODUCTION Laticifers are defined as cells or series of connected cells containing a fluid called latex and forming systems that permeate various tissues of the plant body (Evert 2006). Latex is known to contain a variety of proteins and enzymes. In particular, the latex of the genus Ficus contains ficin (cysteine proteases), indicating appreciable proteolytic activity, which in turn has a role in defense against herbivores and pathogens (Williams et al. 1968; Konno 2011). We focused on the distribution of laticifers in the common edible fig tree (Ficus carica L.) that might prevent invasion by an ambrosia beetle, Euwallacea interjectus, the vector of a vascular wilt fungus, Ceratocystis ficicola (Kajii et al. 2013). Laticifers of the genus Ficus are mentioned frequently in the literature together with other Moraceae species (Vreede 1949; Metcalfe 1967; Davies et al. 1982; Rachmilevitz & Fahn 1982; Van Veenendaal & Den Outer 1990; Kang et al. 2000; Palhares et al. 2007). However, descriptions of the laticifers in the secondary phloem and secondary xylem of F. carica are fragmentary (Ogata et al. 2008). The purpose of the present study is to describe the distribution and structure of laticifers in the branch and trunk of F. carica, and to estimate the potential contribution of laticifers to blocking the activity of the pests. © International Association of Wood Anatomists, 2014 Published by Koninklijke Brill NV, Leiden DOI 10.1163/22941932-00000052 110 IAWA Journal 35 (2), 2014 MATERIALS AND METHODS An 8-year-old trunk of Ficus carica was harvested in the orchards of Hiroshima Prefecture. Current-year branches of F. carica were collected from the Hiroshima Prefectural Technology Research Institute after making horizontal incisions with a knife to induce latex excretion. All specimens were fixed with FAA (formalin:acetic acid:30% ethanol, 5:5: 90 v/v). After the specimens were washed under tap water overnight, transverse, tangential, and radial sections (20 µm thick) were cut from those specimens with a sliding microtome. Sections were stained with safranin-fast green (Johansen 1940) or nile blue (Conn 1977), or kept without staining, and then were processed for light microscopy (Nikon, Eclipse 80i). Some of the thick sections (100 µm thick) including laticifers were embedded in Spurr media (Spurr Low-Viscosity Embedding Kit) after the presence of laticifers was checked under the light microscope. Then, 3-µm-thick sections were made with a rotary microtome. Sections were observed with a differential interference microscope. RESULTS AND DISCUSSION Laticifers were observed in the cortex (Fig. 1), secondary phloem, and pith (Fig. 2) of both the current-year branches and the 8-year-old trunk. In addition, they were found in the secondary xylem and cambium of the 8-year-old trunk. The laticifers in the phloem and pith were of the branched, non-articulated type (Fig. 3 & 4), as reported by Rachmilevitz and Fahn (1982). Laticifers in the secondary xylem extended in a horizontal direction within the rays (Fig. 5). Wheeler et al. (1989) described that non-articulated laticifers are not uncommon in the xylem rays of Apocynaceae, Asclepiadaceae, Euphorbiaceae, and Moraceae. In our material, laticifers found in the secondary xylem sometimes were branching and extending axially interspersed among the fibers (Fig. 6). The incidental presence of radial laticifers in Ficus is mentioned by Koek-Noorman et al. (1984) but not that of axial laticifers. Axial non-articulated laticifers have not yet been reported in the tissue of F. carica, although they are known to exist in the secondary xylem of Artocarpus species (Moraceae) (Topper & Koek-Noorman 1980). It is well documented that non-articulated, branched laticifers develop from single initials or primordia during development of the embryo, and then elongate as the plant grows (Mahlberg 1961, 1963; Cass 1985; Murugan & Inamdar 1987; Rudall 1987, 1994; → Figure 1–7. Laticifers in stem tissues of Ficus carica. – 1 & 2: Axial laticifers (L) in the cortex (1) and pith (2) of a current-year branch. Cross sections. Nile blue staining. – 3: An axial laticifer (L) in the secondary phloem. Tangential section. Safranin-fast green staining. – 4: A branching laticifer (L) in the secondary phloem. Tangential section. Nile blue staining. – 5: A long horizontal laticifer extending in the ray tissue of the xylem (L). Radial section. Safranin-fast green staining. – 6: Connection or branching of the horizontal and axial laticifer (L) in the secondary xylem. Radial section. Safranin-fast green staining. – 7: A horizontal laticifer (L) extending in the secondary phloem, cambium, and the secondary xylem. Cross section. Rectangular area (*) see Fig. 8. — Scale bars in 3, 4, 6 & 7 = 50 µm, in 2 = 100 µm, in 1 & 5 = 200 µm. Kajii et al. – Laticifers of Ficus 1 3 111 2 4 6 5 7 Abbreviations of the two plates: A = axial parenchyma; C = cortex, Ca = cambium; CZ = cambial zone; L = laticifer; Ph = phloem; Pi = pith; S = sieve tube; ; W = wound; X = xylem. 112 IAWA Journal 35 (2), 2014 8 9 8 10B 10A 10C Figure 8–10. Laticifers in stem tissues of Ficus carica. – 8: High magnification of a horizontal laticifer existing in the cambial zone. – 9: A laticifer in the secondary phloem (L) turned at a right angle and extending horizontally in the xylem. Radial section. – 10: Cortex of currentyear branches. After wounding, laticifers actively secreted latex (L) (A) and laticifers around the wound were stained purple (B). In non-treated areas, latex was not visible (C). Radial section (A) and cross section (B, C). Nile blue staining. — Scale bars in 8 = 10 µm, in 9 & 10A = 200 µm, in 10B & C = 500 µm. Kajii et al. – Laticifers of Ficus 113 Van Veenendaal & Den Outer 1990; Roy & De 1992; Da Cunha et al. 1998). It generally is presumed that the laticifers penetrate the rays from the pith (Evert 2006). It has also been recorded that laticifers traverse the vascular cambium in some Asclepiadaceae, Euphorbiaceae, and Moraceae (Blaser 1945; Artschwager 1946; Vreede 1949; Rudall 1989). To understand the differentiating process of laticifers in Ficus, we observed the structure of the laticifers in the cambial region in detail with thin sections embedded in plastic (Fig. 7). Under the differential interference microscope, laticifers were observed extending horizontally in the secondary phloem and xylem traversing the cambial initial layer (Fig. 8, arrow). In addition, we found that some longitudinal laticifers in the secondary phloem turned at a right angle and then extended horizontally into the xylem (Fig. 9, arrow). These findings suggest that longitudinal laticifers in the phloem and horizontal laticifers in the secondary xylem are continually formed. For the few examples of laticifers that extend between the phloem and xylem and are changing their direction at a right angle, further discussion on the site of origin will be difficult. Tangential septa were not found in the horizontal laticifers in the cambial zone (Fig. 8, arrow). This shows that laticifers did not form by cell division in the cambial zone. That is, the cell wall of laticifers must be elongating in the cambial zone, probably with the repetitive addition of wall substances. Fujii and Sudo (1988) described that the laticifer walls in the matured ray tissue of Pimelodendron amboinicum (Euphorbiaceae) shows a loose texture and is composed of different layers. Although the process of laticifer wall formation in Ficus has not been observed, their data suggest that laticifers accumulate wall substances with the continual elongation at cambial zone. Laticifers were found more frequently in the secondary phloem than in the secondary xylem of specimens of both ages (current-year- and 8-year-old). In specimens wounded just before harvest by a thin blade, laticifers and latex that was exuded from the incision wound into adjacent broken cells (Fig. 10A, B) stained purple to pink with nile blue– much stronger so than specimens without incision wounds (Fig. 10C). This indicates that laticifers actively secreted latex in reaction to wounding. The latex may have a role in preventing pest invasion. This defense system, however, does not always function in the case of fig trees infected with a wilt pathogen, Ceratocystis ficicola, vectored by the ambrosia beetle, Euwallacea interjectus. As described in our previous report (Kajii et al. 2013) invasion of that vector beetle was successful from the necrotic area of the phloem where latex exudation had stopped, although the invasion had failed from the sound phloem. The function of laticifers in xylem is not clear because the activities of the beetle and pathogenic fungus were not prevented in the sapwood of fig trees (Kajii et al. 2013). Further observations will be necessary to assess whether the laticifers in phloem and xylem can be effective as defense system in fig trees, or whether certain pathogens and beetles have developed strategies to brake this defense system. Acknowledgement We wish to acknowledge valuable discussions with Dr. T. Fujii on the structure and distribution of laticifers in trees. 114 IAWA Journal 35 (2), 2014 REFERENCES Artschwager E. 1946. Contribution to the morphology and anatomy of Cryptostegia (Cryptostegia grandiflora). U.S. Dep. Agric. Tech. Bull. 915. Blaser HW. 1945. Anatomy of Cryptostegia grandiflora with special reference to the latex system. Amer. J. Bot. 32: 135–141. Cass DD. 1985. Origin and development of the non-articulated laticifers of Jatropha dioica. Phytomorphology 35: 133–140. Conn HJ. 1977. Biological stains. In: Lillie RD (ed.), A handbook on the nature and uses of the dyes in the biological laboratory. Ed. 9. Williams & Wilkins Co., Baltimore, USA. 692 pp. Da Cunha M, Costa CG, Machado RD & Miguens FC. 1998. Distribution and differentiation of the laticifer system in Chamaesyce thymifolia (L.) Millsp.(Euphorbiaceae). Acta Bot. Neerl. 47: 209–218. Davies FT, Lazarte JE & Joiner JN. 1982. Initiation and development of roots in juvenile and mature leaf bud cuttings of Ficus pumila L. Amer. J. Bot. 69: 804–811. Evert RF. 2006. Esau’s plant anatomy: Meristems, cells, and tissues of the plant body: their structure, function, and development. Ed. 3. John Wiley & Sons, New York. Fujii T & Sudo S. 1988. The cell wall structure of the latex tube in the rays of Pimelodendron amboinicum Hassk. (Euphorbiaceae). Mokuzai Gakkaishi 34: 195–199. Johansen DA. 1940. Plant microtechnique. McGraw Hill, New York. 523 pp. Kajii C, Morita T, Jikumaru S, Kajimura H, Yamaoka Y & Kuroda K. 2013. Xylem dysfunction in Ficus carica infected with wilt fungus Ceratocystis ficicola and the role of the vector beetle Euwallacea interjectus. IAWA J. 34: 301–312. Kang H, Kim YS & Chung GC. 2000. Characterization of natural rubber biosynthesis in Ficus benghalensis. Plant Physiol. & Biochem. 38: 979–987. Koek-Noorman J, Topper SMS & Ter Welle BJH. 1984. The systematic wood anatomy of the Moraceae (Utricales). III. Tribe Ficeae. IAWA Bull. n.s. 5: 330–334. Konno K. 2011. Plant latex and other exudates as plant defense systems: Roles of various defense chemicals and proteins contained therein. Phytochemistry 72: 1510–1530. Mahlberg PG. 1961. Embryogeny and histogenesis in Nerium oleander. II. Origin and development of the non-articulated laticifer. Amer. J. Bot. 48: 90–99. Mahlberg PG. 1963. Development of non-articulated laticifer in seedling axis of Nerium oleander. Bot. Gaz. 124: 224–231. Metcalfe CR. 1967. Distribution of latex in the plant kingdom. Econ. Bot. 21: 115–127. Murugan V & Inamdar JA. 1987. Studies in the laticifers of Vallaris solanacea (Roth) O.Ktze. Phytomorphology 37: 209–214. Ogata K, Fujii T, Abe H & Baas P. 2008. Identification of the timbers of southeast Asia and the western Pacific. Shiga-ken, Kaiseisha Press, Japan. 400 pp. Palhares D, De Paula JE, Rodrigues Pereira LA & Dos Santos Silveira CE. 2007. Comparative wood anatomy of stem, root and xylopodium of Brosimum gaudichaudii (Moraceae). IAWA J. 28: 83–94. Rachmilevitz T & Fahn A. 1982. Ultrastructure and development of the laticifers of Ficus carica L. Ann. Bot. 49: 13–22. Roy AT & De DN. 1992. Studies on differentiation of laticifers through light and electron microscopy in Calotropis gigantea (Linn.) R. Br. Ann. Bot. 70: 443–449. Rudall P. 1987. Laticifers in Euphorbiaceae – a conspectus. Bot. J. Linnean Soc. 94: 143–163. Rudall P. 1989. Laticifers in vascular cambium and wood of Croton spp. (Euphorbiaceae). IAWA Bull. n.s. 10: 379–383. Rudall P. 1994. Laticifers in Crotonoideae (Euphorbiaceae): homology and evolution. Ann. Missouri Bot. Gard. 81: 270–282. Kajii et al. – Laticifers of Ficus 115 Topper SMC & Koek-Noorman J. 1980.The occurrence of axial latex tubes in the secondary xylem of some species of Artocarpus J.R. & G. Forster (Moraceae). IAWA Bull. n.s. 1: 113–119. Van Veenendaal WLH & Den Outer RW. 1990. Distribution and development of the non-articulated branched laticifers of Morus nigra L. (Moraceae). Acta Bot. Neerl. 39: 285–296. Vreede MC. 1949. Topography of the laticiferous system in the genus Ficus. Ann. Bot. Gard. Buitenzorg 51: 125–149. Wheeler EA, Baas P & Gasson PE (eds.). 1989. IAWA list of microscopic features for hardwood identification. IAWA Bull. n.s. 10: 219–332. Williams DC, Sgarbieri VC &Whitaker JR. 1968. Proteolytic activity in the genus Ficus. Plant Physiol. 43: 1083–1088. Accepted: 12 February 2014