Linkage Mapping of Lr16 and Lr17 in Wheat
advertisement

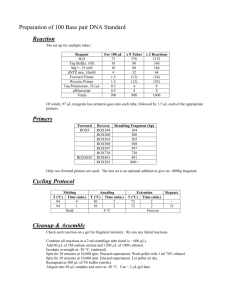
Linkage Mapping of Lr16 and Lr17 in Wheat Barbara Bremenkamp Kansas State University Honors Program Spring 2005 TABLE OF CONTENTS ABSTRACT ......................................................................................................... ii INTRODUCTION ...................................................................................................1 MATERIALS AND METHODS .............................................................................1 Plant Material ...............................................................................................1 Microsatellite Analysis ................................................................................2 Lr16 ..................................................................................................2 Lr17 ..................................................................................................2 High Resolution Agarose Gels.....................................................................3 Acrylamide Gels ..........................................................................................3 Capillary Fragment Analysis .......................................................................4 RESULTS Lr16 Lr17 ..........................................................................................................4 ..........................................................................................................4 ..........................................................................................................5 CONCLUSION ........................................................................................................6 REFERENCES ........................................................................................................8 TABLES AND FIGURES Table 1 Primer sets tested on Lr16 Table 2 Primer sets tested on Lr17 Table 3 Fragment sizes of tailed primers Figure 1 Lr16 Linkage Map Figure 2 Lr17 Linkage Map ABSTRACT Wheat leaf rust, caused by Puccinia triticina, is a constant problem for North American wheat producers. Each year, millions of acres of wheat are infected by this economically important fungus. The infected leaves senesce prematurely, affecting the ability of the wheat plant to complete kernel fill, in turn reducing yield. Lr16 and Lr17, leaf rust resistance genes in wheat, are present in many wheat varieties available today. In combination with other genes, resistance can be enhanced. Currently, no markers are available to follow Lr16 or Lr17 in breeding lines. The objective of this research is to identify molecular markers linked to Lr16 and Lr17. SSR markers were used on mapping populations of both Lr16 and Lr17. Capillary fragment analysis was performed on Lr17 primers. Six SSR markers were closely linked to Lr17. Xgwm614, 1.2 cM away, and Xwmc382, 4.4 cM, flank Lr17. DNA fragments of Lr16 were separated on high resolution agarose gels, however, none of the markers were closely linked to Lr16. We are currently running additional SSR primers on the Lr16 population using capillary fragment analysis. INTRODUCTION The most limiting disease in wheat production is leaf rust, which is caused by the fungal pathogen Puccinia triticina. This persistent pest decreases yield by reducing the number of kernels per spike, reducing test weight, and diminishing kernel quality. The amount of damage inflicted by leaf rusts varies with the degree of infection and plant resistance. However, up to 40 percent of yield can be lost with severe infection on susceptible plants (Bowden np). Although fungicides can be applied to fields, the most economical and effective way to control leaf rust is through plant resistance. Over 50 leaf rust resistance genes have been identified in wheat. Lr16 and Lr17 have been overcome by virulent rust races and are not effective resistance genes alone. However, in combination with other genes, resistance can be enhanced in wheat varieties that carry Lr16 or Lr17. Lr16 has been used extensively in spring wheat breeding populations in Canada. Lr17 is present in the Kansas hard red winter wheat cultivar Jagger. Jagger has been a popular variety throughout Kansas since its release in 1995. In 2003, 45.2% of all Kansas wheat acres were planted to Jagger (USDA-NASS). Although total acreage planted has declined, Jagger is still the most popular variety in Kansas taking 28.2% of the state’s acreage in 2005 (USDA-NASS). Lr16 has been mapped to the short arm of chromosome 2B while Lr17 has been mapped to the short arm of chromosome 2A. A closely linked molecular marker for Lr16 and Lr17 would be extremely beneficial for wheat breeders and allow them to follow these genes through breeding populations. MATERIALS AND METHODS Plant material Thatcher isolines were obtained from the Cereal Disease Laboratory (USDA, St. Paul, MN). These cultivars were backcrossed for several generations to obtain homogeneity of a specific leaf rust resistance gene. An F2 mapping population was created by crossing Chinese Spring, which is susceptible to leaf rust, to both Thatcher isolines Lr16 and Lr17, resistant to specific leaf rust races. The F2 progeny were screened with the leaf rust isolates PRTUS-3 and PRTUS-1 to evaluate for resistance to Lr16 and Lr17, respectively. Leaf tissue was collected from three to four leaf stage seedling plants, frozen in liquid nitrogen, and stored at -80 oC until extraction. Genomic DNA was extracted using a modified CTAB extraction protocol (Hulbert et al., 1991). Microsatellite analysis Lr16 Microsatellite markers mapped to chromosome 2B were used to test for polymorphism in the parents. Forty primer sets were tested using Polymerase Chain Reaction (PCR). Each 25 L PCR reaction consisted of the following reagents: 250 ng genomic DNA, 10 pmoles of each primer set, 1x PCR Buffer (Sigma), 1.0 mM MgCl, 1x dNTP’s, 1.25 U Taq DNA polymerase (Sigma). Cycling conditions on an MJ Research PTC-225 included denature of 92°C for 3 minutes and 35 cycles of denature of 92°C for 1 minute, anneal of 50°C, 52°C, 55°C or 60°C (depending on primer set) for 2 minutes; extension of 72°C for 2 minutes. Lr17 PCR was also used to screen microsatellite markers located on 2AS for polymorphisms between Chinese Spring and Thatcher Lr17. The same PCR reaction listed above was used for fragment amplification for acrylamide gels. However, the reaction was modified for capillary fragment analysis. Each 25 L reaction consisted of the following reagents: 100 ng genomic DNA, 1x PCR Buffer (Sigma), 1.0mM MgCl, 1x dNTP’s, 1.25 U Taq DNA polymerase (Sigma), 5.0 picomoles of reverse primer, 1.0 picomoles of M13-tailed forward primer, and 5.0 picomoles of M13 primers labeled with one of the following dyes: 6-FAM, NED, PET, or VIC (Applied Biosystems). The M13tailed forward primers were constructed by adding 18 basepairs to the original forward primer sequence. An MJ Research PTC-225 thermal cycler was programmed for the following cycle: denature of 95°C for 5 minutes and 10 cycles of denature of 95°C for 45 seconds, anneal of 68°C for 5 minutes; extension of 72°C for 1 minute. Each cycle reduced the anneal temperature by 2°C with the final 10 cycles running a 50°C annealing temperature. A final elongation cycle of 72°C for 5 minutes completed the reaction. High-resolution agarose gels The amplified DNA fragments from Lr16 primer sets were separated on a 2.3% Metaphor agarose gel. Thirteen of the Lr16 primer sets showed polymorphism and were then tested using Bulked Segregate Analysis (BSA). Six of the primer sets tested by BSA showed polymorphism and were used on 94 individuals in the population. The highresolution gels were scored for presence or absence of bands or shifts in bands (depending on the primer set). This data was entered into Mapmaker Macintosh V2.0. for linkage analysis. Acrylamide gels DNA fragments from Lr17 primer sets were separated on 6% non-denaturing acrylamide gels. The gels were made with a 30% acrylamide stock solution, AccuGel 19:1 from National Diagnostics. The gel preparation protocol from National Diagnostics was modified by using no urea and increasing the amount of TEMED and 10% (w/v) Ammonium Persulfate to 80 L and 500 L L, respectively, per 100 mL of casting solution. The gels were run on a Dual Mega-Gel rig from CBS Scientific. The running buffer was 0.5x TBE. Gels were run at constant power (40W) for 600 to 700 volt-hours. The acrylamide gels were scored and data was entered into Mapmaker Macintosh V2.0. Capillary fragment analysis Capillary fragment analysis was performed on an ABI 3730. Samples were prepared by pooling 3 L of PCR product from four separate primer sets, each with a different dye. The DNA pool was vortexed and spun down. One L of the pooled DNA was added to a mixture of 6 L of Hi-Di formamide, 0.25 L of LIZ-500 size standard (Applied Biosystems), and 3 L of water. The samples were vortexed well and spun down. The 96-well plate was placed on an MJ Research PTC-225 thermal cycler for 5 minutes at 95°C and then on an ice slurry for 5 minutes before being submitted to the Kansas State University Sequencing facility. Raw data files from the ABI 3730 were imported into GeneMarker v2.1 for fragment analysis. The scoring data was entered into Mapmaker Macintosh V2.0. for linkage analysis. RESULTS Lr16 Polymorphisms were found in thirteen of the forty Lr16 primer sets tested on the parents. Bulked Segregate Analysis yielded nine primer sets showing polymorphism. Six of these primer sets were polymorphic in the population and scored. Table 1 gives the primer sets tested and results. Although polymorphisms were found in the primer sets tested, none were closely linked to Lr16 in our initial screeing. Screening the F2 progeny with leaf rust isolate PRTUS-3 only showed which plants were homozygous recessive (susceptible). We were unable to determine which plants were heterozygous or homozygous dominant from the resistant plants. Somers et. al. mapped Lr16 and listed Xwmc764 and Xwmc611 as closely linked markers (Somers 1109). We verified the presence of Lr16 in our mapping population by running four SSR markers. The closest marker was Xwmc611 2.5 cM away while Xwmc764 1.1 cM away from Xgwm611. Xwmc597 was mapped 103.6 cM away and Xwmc154 was not linked in our population. Somers showed these primers as only distantly linked to Lr16. Figure 1 shows the preliminary linkage map from our Lr16 mapping population. Lr17 The resistance reaction of the parents and progeny were scored with the resistance reaction scale from Wheat Rusts by McIntosh. Scores on this scale range from a fleck ‘;’, hypersensitive resistance reaction, to four, susceptible reaction. Chinese Spring exhibited a three to four reaction while Thatcher isoline Lr17 displayed a fleck. The progeny of the Chinese Spring, Thatcher Lr17 cross that exhibited a three or four were designated susceptible while a fleck or one reaction were designated resistant. Of the 101 progeny tested, 25 were susceptible and 76 were resistant. This ratio confirms the presence of a single, dominant resistance gene. Forty Lr17 primers were tested on the parents. Thirteen of the primers showed polymorphisms between the parents and in a sample of six individuals in the population. Table 2 gives a complete list of the Lr17 primers tested. Fragment separation on acrylamide gels yielded two primers that could be scored on the population. However, these primers were only loosely linked to Lr17. The remaining primers had just slight shifts. As a result, consistent scoring was impossible. When the primers that were unscoreable on acrylamide were run by capillary analysis, polymorphisms were evident. The ABI 3730 could detect as little as two base pair differences. Overall, 19 of the primers screened showed polymorphisms. Using the Somers consensus map, we focused on the primers located near the end of chromosome 2AS. Twelve of these primers were used on the entire population. Table 3 illustrates the fragment sizes. PRC products were then analyzed with the ABI 3730 and scored. Mapmaker Macintosh V2.0. linked six primers adjacent to Lr17. Xgwm614 was the most closely linked at 1.2 cM away. Xwmc382, flanking Lr17, is 4.4 cM away. Xgwm636 is 1.2 cM from Xgwm614, Xgwm667, and Xgwm407 are each another 1.2 cM away, respectively, and Xgwm512 is 3.8 cM from Xwmc407. Figure 2 illustrates the linkage distances for Lr17. It is possible for SSR markers anneal to sequences on more that one chromosome in the wheat genome. When this occurs, multiple fragments are often amplified and may show both polymorphic and monomorphic bands. This was the case for two Lr17 primers, Xgwm296 and Xgwm210. Each primer amplified three bands, two of which were polymorphic between Chinese Spring and Thatcher Lr17. However, after running the two primers on the entire population, it was evident that neither of the two polymorphic bands were linked to Lr17 this leads us to believe that the monomorphic band is located on chromosome 2B while the other bands are located else where in the genome. CONCLUSIONS Capillary fragment analysis is the most precise method of the three tested to determine fragment size and identify polymorphisms. This method makes more SSR markers available for linkage mapping. The identification of flanking markers Xgwm614 and Xwmc382 being closely linked to Lr17 and Xwmc764 and Xwmc661 linked to Lr16 will allow wheat breeders to follow the genes through their germplasm and potentially combine them with other resistance genes to enhance current levels of leaf rust resistance. The Lr16 and Lr17 mapping populations are currently being expanded to more accurately determine the linkage distance. Additional primers will be tested on the Lr16 mapping population. Expressed sequence tags located on chromosome 2 AS and 2BS will also be evaluated for polymorphisms. REFERENCES Bowden, R.L. “Wheat Leaf Rust.” Kansas State University. www.oznet.ksu.edu. April 28, 2005. Hulbert, S.H., and Bennetzen, J.L. 1991, Recombination at the Rp1 locus of maize. Mol. Gen. Genet. 226:377-382. McIntosh, R.A., Wellings, C.R., and Park, R.F. Wheat Rusts: An Atlas of Resistance Genes. Kluwer Academic Publishers, 1995. Somers, DJ. 2004, A high-density wheat microsatellite consensus map for bread wheat (Triticum aestivum L.). Theoretical & Applied Genetics.109:1105-1114.