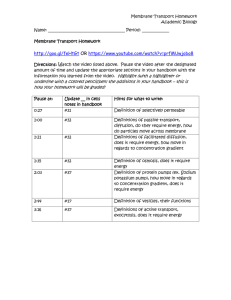
Wiley Manual
advertisement

CAPÍTULO 4 (8 na 6ª Edição Internacional) A ESTRUTURA E FUNÇÃO DA MEMBRANA PLASMÁTICA – PARTE 2 OBJECTIVOS Descrever as funções das membranas celulares. Enumerar os principais componentes moleculares das membranas biológicas e descrever a sua organização espacial. Relacionar as propriedades das membranas com a sua composição química Identificar a localização dos açucares na estrutura da membrana e relacionar com funções possíveis. Descrever diferentes tipos de proteínas presentes nas membranas e relacionar com as funções desempenhadas por essas proteínas. Compreender a importância da fluidez da membrana para a manutenção de todos os mecanismos moleculares da célula. Descrever os mecanismos utilizados pelas células para manter a fluidez das membranas. Descrever a assimetria da membrana e a natureza dinâmica da sua estrutura e função. Descrever os diferentes mecanismos utilizados pelas células para o transporte de materiais através da membrana, como a difusão simples e facilitada, proteínas canal e transporta activo. Enumerar as principais propriedades da membrana dos eritrócitos, como um exemplo em que a composição molecular se encontra bem determinada. LECTURE OUTLINE The Movement of Substances Across Cell Membranes: Diffusion and Osmosis I. Membrane has dual function – it retains dissolved materials of cell so they do not leak out into the environment & it must allow the necessary exchange of materials into & out of the cell A. Lipid bilayer is ideally suited to prevent loss of charged & polar solutes (ions, sugars, amino acids) 1. Must make special provisions for movement of nutrients, respiratory gases, hormones, wastes, etc. B. Membranes are selectively permeable barrier - how movement controlled? - two means for movement both of which lead to net flux of ions/compounds (influx - into cell; efflux - out of cell) 1. Passively by diffusion 2. Actively by an energy-coupled transport process C. Several different processes by which substances move across membranes 1. Simple diffusion through lipid bilayer 2. Simple diffusion through an aqueous, protein-lined channel 3. Facilitated diffusion via a protein transporter 4. Active transport via an energy-driven protein pump capable of moving substances against a concentration gradient II. Energetics of solute movement - depends on magnitude of concentration gradient A. Diffusion is a spontaneous process in which substance moves from region of high concentration to region of low concentration, eventually eliminating concentration difference between the 2 regions B. Depends on random thermal motion of solutes; an exergonic process driven by entropy increase III. When solute is a non-electrolyte, it will move down a concentration gradient (exergonic) A. When nonelectrolyte (uncharged solute) diffuses across membrane, the free-energy change depends on magnitude of concentration gradient B. The following formula describes the movement of nonelectrolyte into the cell: G = RT ln([CI]/[Co]) or G = 2.303 RT log10 ([CI]/[Co]) where: 1. G = free energy change 2. R = gas constant 3. T = absolute temperature 4. [CI]/[Co] = ratio of the concentration of solute on inside (i) & outside (o) surfaces of membrane C. At 25°C, G = 1.4 kcal/mole x log10 ([CI]/[Co]) 1. If the ratio of [CI]/[Co] is less than 1.0, then the log of the ratio is negative, G is negative & the net influx of solute is thermodynamically favored (exergonic) 2. If external solute concentration is 10 times the internal concentration, G = -1.4 kcal/mole 3. Thus, maintenance of a ten-fold concentration gradient represents storage of 1.4 kcal/mole 4. As solute moves into the cell. the concentration gradient decreases, the stored energy is dissipated & the G decreases, until, at equilibrium, G = 0 D. To calculate G for movement of a solute out of cell, the term for concentration ratios is [Co]/[Ci] IV. If solute is an electrolyte (a charged species), the overall charge (potential difference, voltage) difference is also important; bigger difference in charge, bigger difference in free energy A. If charges of electrolyte & compartment to which it is moving are opposite, attraction results; if they are the same, there is repulsion 1. Due to mutual repulsion of ions of like charges, it is thermodynamically unfavorable for an electrolyte to move across membrane into a compartment having a net charge of the same sign 2. If the electrolyte charge is opposite in sign to the compartment into which it is moving, the process is thermodynamically favored 3. The greater the difference in charge (potential difference or voltage) between the 2 compartments, the greater the difference in free energy B. The tendency of an electrolyte to diffuse between 2 compartments depends upon 2 gradients: 1. A chemical gradient, determined by the concentration difference of the substance between the 2 compartments 2. Electric potential gradient, determined by the difference in charge C. Combination of the 2 gradients is electrochemical gradient & it can be very strong; the free energy change for diffusion of an electrolyte into cell is G = RT ln([CI]/[Co]) + zFEm where 1. z is the charge of the solute 2. F is the Faraday constant (23.06 kcal/V .equivalent, where an equivalent is the amount of the electrolyte having 1 mole of charge) 3. Em is the potential difference (in volts) between the 2 compartments D. Suppose a concentration gradient consists of Na+ ions present at tenfold higher concentration outside the cell than inside 1. Voltage across a membrane is typically ~-70 mV, thus the free energy change for the movement of a mole of Na+ ions into the cell under these conditions can be calculated 2. G = -1.4 kcal/mole + zFEm = -1.4kcal/mole + (1)(23.06 kcal/v x mole)(-0.07 V) = -3.1 kcal/mole 3. This compares to the G of –1.4 kcal/mole for a nonelectrolyte with a tenfold concentration difference across a membrane at 25°C 4. Thus under the conditions described, the concentration difference & the electric potential make similar contributions to the storage of free energy across a membrane E. Example: interplay between concentration & potential differences seen in K + ion diffusion out of cell 1. Efflux of K+ is favored by K+ concentration gradient ([K+] is higher inside cell) 2. Efflux is hindered by the electrical gradient that its diffusion creates (leaves higher "-" charge in cell III. Diffusion of substances through membrane A. 2 qualifications must be met before nonelectrolyte can diffuse passively across a membrane 1. Substances must go down gradient (must be present at higher concentration on one side of membrane than the other 2. Membrane must be permeable to the substance B. Membranes are permeable to a given solute in two ways 1. Solute can pass directly through bilayer or 2. Solute can traverse an aqueous channel (pore) that spans the membrane & prevents the solute from coming into contact with lipids of bliayer IV. Factors that determine the ability of molecules to pass directly through membrane A. Polarity of a solute – a measure of polarity or nonpolarity is its partition coefficient 1. A solute's partition coefficient is the ratio of its solubility in a nonpolar solvent (octanol, vegetable oil) to that in H2O under conditions where the nonpolar solvent & H2O are mixed together 2. Higher nonpolar solvent (e. g., oil) : water solubility ratio —> solute more able to pass bilayer (first clue that membrane has lipid layer) 3. In other words, greater lipid solubility leads to faster penetration of the membrane B. Size - smaller molecules pass through membrane faster - small inorganic substances penetrate rapidly; bigger polar molecules do not pass easily or at all 1. If two molecules have approximately equivalent partition coefficients, the smaller one tends to penetrate a lipid bilayer more rapidly than the larger one 2. Very small, uncharged (inorganic) molecules (O2, CO2 NO, H2O) penetrate very rapidly through membranes; these smaller molecules may slip between adjacent phospholipids 3. Larger polar molecules (sugars, amino acids, phosphorylated intermediates, etc.) can’t penetrate membrane 4. Thus, the lipid bilayer of cell membrane is an effective barrier that keeps these larger essential metabolites from diffusing out of the cell 5. Some of these molecules (sugars, amino acids) must enter cells from bloodstream, but cannot do so by simple diffusion; special mechanisms must be available to allow their penetration 6. The use of such mechanisms allows a cell to regulate the movement of substances across its surface barrier V. Diffusion of H2O through membranes – since H2O moves faster through membranes than solutes (dissolved ions, small polar organic solutes; essentially nonpenetrating), membranes called semipermeable A. Osmosis – ready movement of water through a semipermeable membrane from a region of lower solute (high water) concentration to a region of higher solute (low water) concentration 1. Water moves toward hypertonic (higher solute concentration, hyperosmotic) environments & away from hypotonic (lower solute concentration, hypoosmotic) environments 2. 2 solutions with equal solute concentrations - isotonic (no net water movement; isoosmotic) B. Response of cells to nonisotonic environments 1. Animal cells in hypotonic environments take on water (swell) & eventually lyse (RBCs hemolyze) 2. Plant cells in hypotonic environment take in H2O; no lysis due to cell wall - internal pressure (turgor) builds up; important for support for nonwoody plants & nonwoody plant parts 3. Plant cells in hypertonic environment lose water (volume shrinks) - membrane pulls away from cell wall (plasmolysis); without water, plants wilt C. The above observations show that a cell's volume is controlled by the difference between the solute concentration inside the cell & that in the extracellular medium D. Swelling or shrinking of cells in slightly hypotonic or hypertonic media are usually temporary events; within a few minutes, cells recover & return to original volume 1. In hypotonic medium, cells recover as they rid themselves of ions (primarily K + & Cl-) 2. In hypertonic medium, cells recover as they gain ions (mostly Na+ & Cl-) from medium 3. Once [internal solute] (including a high concentration of dissolved proteins) equals [external solute], external & internal fluids are isotonic (no net movement of H2O into or out of cells) VI. Osmosis is important factor in multitude of bodily functions A. Digestive tract secretes several liters of fluid daily; it is reabsorbed osmotically by cells lining intestine; if it is not reabsorbed, as in cases of extreme diarrhea, can lead to rapid dehydration B. Plant cells are usually hypertonic compared to fluid environment (unlike animal cells which are isotonic); thus, water tends to enter cell causing internal (turgor) pressure to push against cell wall 1. In hypertonic medium, cell volume shrinks & membrane pulls away from cell wall (plasmolysis); loss of water via plasmolysis causes plants to lose their support & wilt 2. Turgor pressure provides support for nonwoody plant parts (leaves) & nonwoody plants; loss of water causes plants to lose support & wilt VII. Many cells are much more permeable to water than can be explained by simple diffusion through lipid bilayer A. Peter Agee & colleagues (Johns Hopkins Univ., early 1990s) – attempted to isolate & purify membrane proteins responsible for Rh antigen on surface of RBCs 1. While trying this they identified a protein they thought might be the long-sought water channel of the erythrocyte membrane 2. To test hypothesis, they engineered frog oocytes to incorporate the newly discovered protein into their plasma membranes & then placed the oocytes in a hypotonic medium 3. As predicted, the oocytes swelled due to the water influx & eventually burst 4. They had discovered a family of small integral proteins (aquaporins) that allow the passive movement of water from one side of plasma membrane to the other B. Each aquaporin subunit (in the 4-subunit protein) contains a central channel that is lined primarily by hydrophobic amino acid residues & is highly specific for water molecules 1. A billion or so water molecules can pass – in single file – through each channel every second 2. At the same time, H+ ions, which normally hop along a water molecule chain, are not able to penetrate these open pores C. The apparent mechanism by which these channels are able exclude protons has been suggested by a combination of X-ray crystallographic studies & computer-based simulations 1. X-ray crystallography revealed the protein structure; computer simulations put the protein structure into operation 2. Very near its narrowest point, aquaporin channel wall contains pair of precisely positioned "+" charges (residues N203 & N68) 3. These charges attract O atom of each H2O molecule as it speeds through constriction in protein 4. This interaction orients the central water molecule in a position that prevents it from maintaining H bonds that normally link it to its neighboring water molecules 5. This removes the bridge that would normally allow protons to move from one water molecule to the next D. Aquaporins are particularly prominent in cells, like those of a kidney tubule or plant root, where the passage of water plays a crucial role in the tissues' physiological activities 1. The hormone vasopressin, which stimulates water retention by the collecting ducts of the kidney, acts by way of one of these proteins (AQP2) 2. Some cases of the inherited disorder congenital nephrogenic diabetes insipidus have been traced to mutations in this aquaporin channel 3. Persons suffering from this disease excrete huge quantities of urine because their kidneys do not respond to vasopressin The Movement of Substances Across Cell Membranes: Diffusion of Ions I. Ions go through ion channels (integral proteins surrounding aqueous pores) since core of lipid bilayer is highly impermeable to charged substances like Na+, K+, Ca2+, & ClA. Most channels are very selective allowing only one ion through in a downhill direction B. Ion conductance (rapid movement across membranes) is critical in many cell activities like: 1. Nerve impulse formation & propagation, secretion of substance into extracellular space, muscle contraction, cell volume regulation, opening of plant leaf stomatal pores 2. Mutations in genes encoding ion channels can lead to many serious diseases C. Alan Hodgkin & Richard Keynes, Cambridge Univ. (1955) – proposed that membranes contain ion channels (openings in membranes permeable to specific ions); worked on squid giant nerve cells D. Bertil Hille (Univ. of Washington) & Clay Armstrong (Univ. of Penna.), late 1960s & 1970s – obtained evidence for the existence of ion channels E. Bert Sakmann & Erwin Neher, Max Planck Inst. in Germany (late 1970s & early 1980s) – proof; developed methods to monitor ionic current passing through a single ion channel 1. Used very fine micropipette-electrodes made of polished glass placed on outer cell surface & sealed to membrane by suction 2. Voltage across the membrane can be maintained (clamped) at any particular value 3. Current originating in the small patch of membrane surrounded by pipette can be measured 4. These landmark studies were first successful studies into activities of individual protein molecules II. Bewildering variety of ion channels identified, each with integral protein(s) surrounding aqueous pore; most are highly selective in allowing one particular type of ion to pass through pore A. Diffusion is always downhill (from higher to lower energy state; higher to lower concentration) 1. Ion channels are bidirectional allowing ion passage in either direction; net flux depends on electrochemical gradient B. Comparisons of amino acid sequences of different types of ion channels in diverse organisms (bacteria, plants, animals) show that they are all members of a small number of giant superfamilies 1. Members of given superfamily may have very different ion selectivities, but all are very similar in amino acid sequence & overall structure 2. Suggests they are derived from a single protein present in a common ancestor alive over a billion years ago C. Most ion channels can exist in either an open or a closed conformation; said to be gated 1. Opening & closing of gates is subject to complex physiological regulation 2. Can be induced by a variety of factors depending on the particular channel III. Three major categories of gated channels are distinguished A. Voltage-gated channels – conformational state depends on the difference in ionic charge on the 2 sides of membrane B. Ligand-gated channels – conformational state depends on binding of specific molecule (the ligand), which is usually not the solute that passes through the channel 1. Some ligand-gated channels are opened (or closed) after binding of molecule to outer surface of channel; others open (or close) after binding of ligand to inner surface of channel 2. Neurotransmitters (e.g., acetylcholine) act on outer surface of certain cation channels; cyclic nucleotides (e.g., cAMP) act on inner surface of certain calcium ion channels C. Mechano-gated channels – conformational state depends on mechanical forces (e.g., stretch tension) that are applied to the membrane 1. Members of one family of cation channels are opened by the movements of stereocilia on the hair cells of the inner ear in response to sound or motions of the head IV. Structure & function of K+ ion channels – Roderick MacKinnon et al., Rockefeller Univ. (1998) – first atomicresolution image of an ion channel protein, a bacterial K+ ion channel called KcsA A. Figuring out structure led directly to learning mechanism by which the channels select overwhelmingly K+ ions over Na+ ions while allowing incredibly rapid K+ ion conductance through membrane 1. The mechanisms of ion selectivity & conductance in this bacterial channel are thought to be virtually identical to those operating in the much larger mammalian channels 2. Evidently, the basic challenges in operating an ion channel were solved relatively early in evolution, although many refinements appeared over the next billion or 2 years B. KcsA channel consists of 4 subunits; each subunit contains 2 membrane-spanning helices (M1 & M2) & a pore region (P) at the extracellular end of the channel 1. P consists of a short pore helix that extends about one-third of the width of the channel & a nonhelical loop that forms the lining of a narrow selectivity filter 2. The selectivity filter acquires its name because of its role in allowing the passage of only K + ions 3. Selectivity filter lining contains highly conserved pentapeptide (Gly-Tyr-Gly-Val-Thr or GYGVT) 4. Mutations within this stretch of amino acids often destroy the channel's ability to discriminate between K + & Na+ ions C. The X-ray crystal structure of the KcsA channel shows that the backbone carbonyl (C=O) groups from the conserved pentapeptide create 5 successive rings of oxygen atoms 1. 4 rings are made up of carbonyl oxygens from the polypeptide backbone & 1 ring consists of oxygen atoms from threonine side chain 2. Each ring contains 4 oxygen atoms (one from each subunit) & has a diameter of ~3Å, which is slightly larger than the 2.7Å diameter of a K+ ion that has lost its normal hydration shell 3. Thus, the electronegative O atoms that line the selectivity filter can substitute for the shell of water molecules that are displaced as each K+ ion enters the pore 4. While the selectivity filter is a precise fit for a dehydrated K + ion, it is much larger than the diameter of a dehydrated Na+ ion 5. Thus, a Na+ ion cannot interact with the 4 carbonyl O atoms needed to stabilize its structure; thus the smaller Na + ions cannot overcome the higher energy barrier required to penetrate the pore D. KcsA structure has been determined at very high (2Å) resolution; it has allowed investigators to visualize individual K+ ions & water molecules 1. The selectivity filter in the models contains 4 potential K+ ion binding site, only 2 of which are occupied at any given time 2. K+ ions are thought to move 2 at a time from one pair of binding sites to the next pair (sites 1 & 3 to sites 2 & 4) 3. The entry of a third K+ ion into the selectivity filter creates an electrostatic repulsion that ejects the ion bound at the opposite end of the line 4. Studies indicate that there is virtually no energy barrier for an ion to move from one binding site to the next, which accounts for the extremely rapid flow of ions across the membrane E. The KcsA channel has a gate, like eukaryotic channels; its opening & closing is regulated by medium pH, rather than by voltage across the membrane or binding of a ligand; it opens in response to very low pH 1. It has been impossible to crystallize the KcsA channel in its open conformation 2. However, the structure of a homologous prokaryotic K+ channel (called MthK) in its open conformation has been crystallized & its structure determined F. Comparison of MthK open structure & the homologous protein KcsA closed structure strongly suggested that their gating happens by conformational changes of the cytoplasmic ends of the inner (M2) helices 1. In closed conformation, the M2 helices are straight & cross over one another to form a "helix bundle" that seals the cytoplasmic face of the pore 2. Channel opens when M2 helices bend by ~30° at specific hinge point where glycine residue is found V. Example of eukaryotic voltage-gated channels: K+ ion channels; genes encoding a variety of distinct voltage-gated K+ (or Kv) channels have been isolated & their molecular anatomy scrutinized A. The Kv channels of plants play an important role in salt & water balance & in cell volume regulation; Kv channels of animals are best known for their role in muscle & nerve function 1. These more complex eukaryotic versions are thought to perform in a manner similar to prokaryotic channels B. Members of this protein family have their N- & C-terminal domains situated on cytoplasmic side of membrane C. Eukaryotic Kv channels contain 6 membrane-associated helices (named S1–S6) grouped into 2 functionally distinct domains 1. A pore domain – has same basic architecture as that of entire bacterial channel & contains selectivity filter that permits the selective passage of K+ ions a. Helices M1 & M2 & the P segment of the KcsA channel are homologous to helices S5 & S6 & the P segment of the voltage-gated eukaryotic channels b. Like the M2 helices of KcsA, the S6 helices line much of the pore & their configuration determines whether the gate to the channel is open or closed 2. A voltage-sensor domain – consists of helices S1-S4; senses the voltage across the plasma membrane VI. The 3D crystal structure of a complete eukaryotic Kv channel purified from rat brain was determined by use of a mixture of detergent & lipid throughout the purification & crystallization process A. Like KcsA channel, a single eukaryotic Kv channel consists of 4 homologous subunits arranged symmetrically around the central ion-conducting pore 1. The selectivity filter, & thus the presumed mechanism of K+ ion selection, is virtually identical in the prokaryotic KcsA & eukaryotic Kv proteins 2. The gate leading into a Kv channel is formed by the inner ends of the S6 helices & is thought to open & close in a manner roughly similar to that of the M2 helices of the bacterial channel B. The S4 helix, which contains several positively charged amino acid residues spaced along the polypeptide chain, acts as the key element of the voltage sensor 1. The voltage-sensing domain is seen to be connected to the pore domain by a short linker helix denoted as S4-S5 2. Under resting conditions, the negative potential across the membrane keeps the gate closed 3. A change in potential to a more positive value (depolarization) exerts an electric force on the S4 helix 4. This force is thought to cause S4 helix to move in such a way that its positively charged residues shift from a position exposing them to the cytoplasm to a new position exposing them the outside of cell C. Voltage sensing is dynamic process; mechanism cannot be resolved by a single static view of protein 1. Several competing models describing the mechanism of action of voltage sensor are currently debated 2. However it occurs, the S4 helix movement in response to membrane depolarization initiates a series of conformational changes within the protein that opens the gate at the cytoplasmic end of channel D. Once pore opened, >100 K+ pass through channel/msec (nearly rate that would occur by free diffusion in solution) 1. Due to large ion flux, opening of a relatively small number of K+ channels has significant impact on the membrane electrical properties 2. After channel is open a few msec, K+ ion movement is automatically stopped by a process known as inactivation E. Eukaryotic Kv channels typically contain a large cytoplasmic structure whose composition varies among different channels 1. Inactivation of the channel is accomplished by movement of a small inactivation peptide that dangles from the cytoplasmic portion of the protein 2. The inactivation peptide is thought to gain access to the cytoplasmic mouth of the pore by snaking its way through one of 4 "side windows" 3. When one of these dangling peptides moves up into the mouth of the pore, the passage of ions is blocked & the channel is inactivated 4. At a later stage of the cycle, the inactivation peptide is released & the gate to the channel is closed 5. Thus the potassium channel can exist in 3 different states: open, inactivated & closed VII. Potassium channels come in many different varieties – C. elegans, a nematode worm with a body of ~1000 cells has >90 different genes that encode K+ channels A. A single cell, no matter the what kind of organism it is in, is likely to possess a variety of different K + channels that open & close in response to different voltages B. The voltage needed to open or close a particular K+ channel can vary depending on whether or not the channel protein is phosphorylated, which, in turn, is regulated by hormones & other factors 1. Ion channel function is under the control of a diverse & complex set of regulatory agents The Movement of Substances Across Cell Membranes: Facilitated Diffusion I. Facilitated diffusion - diffusion during which substance binds selectively to a membrane-spanning protein (facilitative transporter), which facilitates diffusion process A. The term facilitative transporter distinguishes these proteins from active transporters whose activity is coupled to a process that releases energy 1. Technically, transporter applies to membrane protein that can only bind a solute from one side of membrane at a time & in which shape change is mechanism for solute movement across membrane 2. Definition distinguishes transporters from channels, which, if open, can bind solutes from either side of membrane at same time; distinction between them is becoming blurred as more learned B. Solute binding on one side of membrane changes protein shape, exposing solute to other surface from where it can diffuse down its concentration gradient 1. Since they are passive (not coupled to energy release), they mediate solute movement equally well in both directions; net flux direction depends solely on gradient direction (down gradient) C. Facilitated diffusion is similar in many ways to an enzyme-catalyzed reaction 1. Like enzymes, they are specific for the transported molecule; for example, they distinguish between D & L stereoisomers; like enzymes, they exhibit saturation-type kinetics 2. Move hundreds to thousands of solutes/sec while channels can move millions of ions/sec 3. Like enzymes & ion channels, their activity can be regulated to fit present cell needs 4. Facilitated diffusion is very important in mediating the entry & exit of polar solutes (amino acids, sugars) that cannot penetrate the lipid bilayer II. Example: glucose transporter A. Glucose is body's primary source of direct energy; most mammalian cells have a membrane protein that facilitates the diffusion of glucose from the bloodstream into the cell 1. Gradient favoring glucose diffusion into cell from bloodstream is maintained by phosphorylating glucose after it enters cytoplasm, thus lowering intracellular glucose concentration 2. Humans have at least 5 related transporters (isoforms; GLUT 1-5) that facilitate glucose diffusion into cell; they are distinguished by tissue where found & kinetic & regulatory characteristic B. Increase in blood glucose levels triggers insulin secretion by pancreas, which stimulates glucose uptake into various target cells (most notably skeletal muscle, adipocytes or fat cells) 1. Insulin-responsive cells share a common isoform of the facilitative glucose transporter (specifically GLUT 4) 2. When insulin levels are low, these cells contain relatively few glucose transporters on their plasma membrane; instead, the transporters are present in membranes of cytoplasmic vesicles 3. Rising insulin levels act on target cells to stimulate the fusion of cytoplasmic vesicles to the plasma membrane, which provides the transporters necessary to bring glucose into the cell The Movement of Substances Across Cell Membranes: Active Transport I. Many ion gradients are very strong & are generated by the expenditure of energy (active transport) A. [K+] inside a mammalian cell is at least 100 mM; outside - ~5 mM —> K+ ions "want" to leave cell B. [Na+] - ~150 mM outside cell; inside cell - ~10 - 20 mM —> Na+ ions "want" to enter cell C. [Ca2+] - cytosol concentration 10-7 M; outside cell - 10,000 times higher than that inside cell; this is an even greater difference than with Na+ or K+ II. Establishment of such steep gradients depends on integral proteins that selectively bind solute & move it against a gradient (endergonic process); proteins that carry out active transport are often called pumps A. Such steep gradients cannot occur by either simple or facilitated diffusion B. Driven by conformational shift along with energy expenditure; coupled to exergonic reaction like ATP hydrolysis, light absorption, electron transport, flow of other substances down their gradients III. Coupling active transport to ATP hydrolysis: the Na+-K+ pump; only in animals; primary means to maintain cell volume & establish the steep gradients needed for nerve-muscle impulses A. Jens Skou, Danish physiologist (1957) - found ATP-hydrolyzing enzyme in crab nerve cells 1. It was only active in the presence of Na+ & K+ ions; Mg2+ acts as a cofactor by binding ATP 2. Proposed it to be Na+-K+ pump that transported these ions (also called it Na+/K+-ATPase) B. Pump is unidirectional & pumps 3 Na+ ions out of cell for every 2 K+ ions pumped in & is electrogenic, contributing directly to the separation of charge across the membrane 1. It is a P-type ion pump that requires phosphorylation of pump during cycle 2. Phosphate is transferred from ATP after its hydrolysis to an aspartic acid residue of pump, causing an essential conformational change within the protein 3. Changes alter pump's affinity for the 2 cations so it can pick up Na+ & release K+ & vice versa 4. To pick either ion up from a low concentration region, protein must bind them with relatively high affinity; to release them into much higher concentration region, affinity for ion must drop 5. Thus, the affinity for each ion on the two sides of the membrane must be different 6. This is achieved by phosphorylation, which changes the protein's shape & also serves to expose the ion binding sites to the different sides of the membrane C. The Na+-K+ pump cycle repeats - uses ~33% of most animal cells' energy (~67% in nerve cells) 1. Pump binds 3 Na+ ions on inside of cell 2. A bound ATP molecule is hydrolyzed & phosphate transferred to an aspartate residue of pump 3. Pump's shape shifts from the E1 to the E2 conformation; it opens to outside, exposing Na+-binding sites there; since E2 conformation has reduced affinity for Na+ ions, they are released outside cell 4. Once the 3 Na+ ions are released, the pump now picks up 2 K+ ions 5. Pump is then dephosphorylated 6. A molecule of ATP then binds to the protein, causing it to shift back to original E1 conformation; pump is now open to inside of cell, its affinity for the 2 K+ ions is reduced & they are released 7. The cycle is then repeated D. Digitalis (foxglove plant steroid), a good congestive heart failure treatment for 200 years, inhibits Na+ - K+ pump & leads to chain of events that increases Ca2+ availability inside heart muscle cells 1. This, in turn, strengthens heart contraction E. The Na+/K+ pump is found only in animal cells 1. It may have evolved in primitive animals as primary way to maintain cell volume 2. It may also have evolved as a mechanism to generate steep Na+ & K+ gradients that play a key role in the formation of muscle & nerve cell impulses F. Directionality of pump & its orientation in membrane are important 1. In resealed RBC ghosts, K+ had to be outside, Na+ and ATP inside & pump in membrane 2. Ouabain (inhibitor of cation transport similar to digitalis) inhibits pump only if outside ghosts IV. Other ion transport systems – there is no high resolution structure of the Na+/K+-ATPase, but the 3D structure of the related P-type pump, Ca2+-ATPase is instructive A. Ca2+-ATPase - P-type pump in ER membrane; it actively moves Ca2+ ions out of cytosol into ER lumen 1. The Ca2+ pump is accompanied by exceptionally large conformational changes 2. These changes couple ATP hydrolysis to changes in access & affinity of the ion binding sites B. Plant cells have P-type transporter for H+ - this cell membrane proton pump has key role in secondary solute transport, control of cytosolic pH & as cell growth control by acidification of plant cell wall C. Stomach epithelial lining H+/K+-ATPase - P-type pump secreting concentrated acid solution (up to 0.16 N HCl) into stomach chamber 1. At rest, found in cytoplasmic membranes of stomach lining parietal cells & are nonfunctional 2. With food, hormonal message transmitted to parietal cells that causes pump-containing membranes to move to apical surface 3. They fuse with apical surface membrane & secrete acid (aids digestion but can lead to heartburn) 4. New acid-blocking medications (Zantac, Pepcid, Tagamet) do not inhibit H+/K+-ATPase directly; they block parietal cell receptors & stop hormone activation of cells & thus halt H+/K+-ATPase 5. Another drug, Prilosec, prevents heartburn by inhibiting stomach's H+/K+-ATPase D. V-type pumps use ATP without formation of phosphorylated protein intermediate 1. They actively pump H+ ions across cytoplasmic organelle membranes & vacuoles (hence the name) 2. Found in membranes lining lysosomes, secretory granules & plant cell vacuoles 3. Found in membranes of a variety of cells like kidney tubules - V-type pump helps maintain body's acid-base balance (secretes protons into forming urine) 4. Their structure is similar to that of ATP synthase E. ATP-binding cassette (ABC) transporters – diverse group of proteins that actively transport ions 1. All members of this superfamily share an homologous ATP binding domain; one is involved in cystic fibrosis V. Utilizing light energy for active ion transport - Halobacterium salinarium (previously H. halobium), an archaebacterium that lives in extremely salty environments (Great Salt Lake) A. Its plasma membranes turn purple when conditions anaerobic, due to bacteriorhodopsin which contains retinal (rhodopsin prosthetic group) 1. Rhodopsin is light-absorbing protein of vertebrate retina rods B. Light energy absorption by retinal group induces series of conformational changes in protein, which causes a proton to move from retinal group through a channel in protein to cell exterior C. Pumped H+ is replaced by one transferred to protein from cytoplasm; this generates a steep H+ gradient across membrane D. Gradient subsequently drives ATP synthesis by enzyme (similar to mitochondrial electron transport) VI. Cotransport: Coupling active transport to existing ion gradients A. Gradient establishment can be used to store free energy in cells; the potential energy stored in these ionic gradients is then used in various ways to perform work, e. g. solute transport B. Sodium ions involved in cotransport of glucose across intestinal epithelium & then into bloodstream after high MW polysaccharides have been hydrolyzed into simple sugars within intestinal lumen 1. Movement of glucose across apical membrane of intestinal epithelial cells against a concentration gradient occurs by cotransport with Na+ ions 2. Primary transport of Na+ out of cell done by Na+-K+ ATPase in basal & lateral membranes; keeps Na+ concentration very low within cells (this is the primary active transport system) 3. Na+-K+ ATPase pumps Na+ ions out of cell against a concentration gradient 4. Na+ ion tendency to diffuse back across apical membrane is tapped by epithelial cells to drive glucose molecule cotransport into cell against their concentration gradient (2° active transport) 5. Na+ moves glucose up its gradient as Na+ flows down its gradient across apical membrane C. Transport protein (Na+/glucose cotransporter) moves 2 Na+ ions & 1 glucose molecule with each cycle 1. The crystal structure of a number of bacterial cotransporters have been solved in recent years; this has led to competing hypotheses as to how these proteins operate 2. Surprisingly, some cotransporters appear to contain a gated ion channel, which has blurred the distinction between channels & pumps 3. Regardless of the mechanism of uptake, once inside the cell, glucose diffuses through cell & moves across basal membrane by facilitated diffusion D. The Na+ ion gradient is very strong; the cotransporter is capable of moving glucose into a cell against a gradient greater than 20,000-fold 1. Free energy change for moving 1 mole of Na+ ions into cell is equal to –3.1 kcal/mole & thus –6.2 kcal for 2 moles of Na+ ions is available to move 1 mole of glucose up gradient into cell 2. Using equation for movement of nonelectrolyte like glucose across the membrane: G = RT ln (Ci/Co) = 2.303 RT log10 (Ci/Co) 3. Use equation to calculate how steep a concentration gradient of glucose (X) the cotransporter can generate; at 25°C, -6.2 kcal/mole = 1.4 kcal/mole x log10 X —> log10 X = -4.43 —> X =1/23,000 E. Plants use 2° active transport systems to move many nutrients (sucrose, amino acids, nitrate) into cell, coupled to the downhill influx of H+ ions instead of Na+ ions VII. Types of transport systems – uniports vs. cotransport systems (symports, antiports) A. Uniport system - transport one solute at a time B. Cotransport systems - >1 solute moved across membrane at same time by a single transport molecule 1. 2 transported solutes move in same direction (symport) 2. If the transported solutes move in opposite directions (antiport) a. Many cells maintain proper cytoplasmic pH by coupling downhill Na+ ion movement into cell to H+ ion movement outward; proteins that act as antiports are usually called exchangers Membrane Potentials and Nerve Impulses I. All organisms possess the ability to respond to external stimulation, a property referred to as irritability; it is a function of the cell membrane; seen in nerve cells & amoebae, which can respond to stimuli A. Poke amoeba with fine glass needle —> it withdraws pseudopodia, rounds up, moves in new direction B. Irritability in amoeba depends on same basic membrane properties that lead to formation & propagation of nerve impulses II. Nerve cells (neurons) are specialized for the collection, conduction & transmission of information, which is coded in the form of fast-moving electrical impulses; the basic parts of a neuron are discussed below: A. Neuron nucleus is located within expanded region called cell body, the metabolic center of cell & the site where most of its material contents are manufactured B. Extending from cell bodies of most neurons are a number of miniscule extensions (dendrites), which receive incoming information from external sources, typically other neurons C. Also emerging from cell body is a single, more prominent extension (an axon), which conducts outgoing impulses away from cell body toward the target cell(s) 1. Impulses are generally initiated in axon hillock, the region where the cell body merges into axon 2. Some axons may be only a few micrometers in length; others extend for many meters in body of a large vertebrate (giraffe, whale) 3. Most axons split near their ends into smaller processes, each ending in terminal knob, a specialized site where impulses are transmitted from neuron to target cell 4. Many neurons in brain end in thousands of terminal knobs, allowing these brain cells to communicate with thousands of potential targets D. Most neurons in the vertebrate body are wrapped in lipid-rich myelin sheath III. Voltage (electric) potential difference can exist between inside & outside of cell; a positive ion excess on one side of membrane & a negative ion excess on the other A. One can measure a voltage potential difference with fine glass electrodes (microelectrodes; one inserted into cell cytoplasm, one placed in the extracellular fluid outside the cell) 1. The microelectrodes are then connected to a voltmeter, an instrument that measures a difference in charge between two points B. The potential difference of the squid giant axon was determined - difference is ~70 millivolts with the cell interior being more negative with respect to the outside (-70 mV); not unique to neurons 1. Such potentials seen in all cell types (range from ~-15 to ~-100 mV); in nonexcitable cells (other than neurons & muscle cells), it is called membrane potential 2. In nerve or muscle cells, it is called resting potential, since it can change dramatically C. Voltage magnitude & direction across membrane is determined by ion concentration differences on either side of membrane & their relative permeabilities 1. Na+/K+-ATPase sets up steep gradient of these ions: Na+ ions high outside, K+ ions high inside 2. Na+ should leak in & K+ should leak out through their respective ion channels because of the directions of their gradients 3. But vast majority of open membrane ion channels in resting nerve cell are selective K+ leak channels that allow more K+ to leak out than Na+ leaking in; there is slight Na+ & Cl- permeability D. The nature of K+ leak channels is still controversial 1. Recently, a new K+ channel family (the KCNK family) that lacks S4 voltage sensor & seems not to respond to changes in voltage has been found 2. K+ leak channels appear to be members of this family (mammalian brain tissue studies) 3. These proteins are called twin-pore channels since each subunit contains 2 ion pores rather than the single ion pore found in subunits of other K+ channels E. Since K+ ions are the only charged species with significant permeability in the resting nerve cell, their outflow through membrane leaves excess of negative charges on cytoplasmic side of membrane 1. While the K+ gradient favors continued K+ efflux, the electrical gradient (excess "-" charge inside the cell) favors K+ retention inside cell 2. If these 2 opposing forces are balanced, the system is at equilibrium & no more net K + movement F. Using the Nernst equation, one can calculate membrane potential (V m) that would be measured at equilibrium if the nerve cell membrane is only permeable to K+ ions 1. Nernst equation derived by setting G = 0, which is the case when the ion movement is at equilibrium; in this case, Vm would be equal to potassium equilibrium potential (EK) 2. EK = 2.303 RT/zF x log10 ([K+o]/[K+I]); for a squid giant axon, the internal [K+I] is ~350 mM while the external [K+o]) is ~10 mM 3. Thus, at 25°C (298°K) & z = +1 for the univalent K+ ion, EK = 59Log100.028 = -91 mV 4. A similar calculation of the Na+ equilibrium potential (ENa) would produce a value of ~+55 mV 5. Measurements of voltage across the resting nerve membrane are similar in sign & magnitude (-70 mV compared to –91 mV) to the potassium equilibrium potential 6. The above calculations of potentials suggest that movement of K+ ions is the most important factor in determining resting potential 7. Slight Na+ & Cl- leakage accounts for difference between calculated & measured resting potentials IV. Action Potential (AP) - our present knowledge of membrane potentials & nerve impulses rests on work on giant squid axons in late 1940s & early 1950s by Alan Hodgkin, Andrew Huxley & Bernard Katz A. Squid axon (~1 mm in dia) carries a high speed impulse, so squid can escape rapidly from predators B. Stimulate squid (or other axon) - poke it with fine needle or jolt it with very small electric current 1. Some of its Na+ channel gates open allowing a limited number of ions in; potential gets less negative (drop in polarity between 2 sides of membrane); membrane less polarized (depolarization) 2. If stimulus is small & depolarization is slight (-70 mV to –60 mV) —> potential returns rapidly to resting potential when stimulus stops C. Stronger stimulus depolarizes membrane more & if it gets to certain point (threshold; ~-50 mV), an AP starts; causes voltage-gated Na+ channels to open & Na+ ions flow freely in 1. Na+ ions move down both their concentration & electric gradients; membrane potential briefly reverses itself becoming positive (~+40 mV); close to the Na+ equilibrium potential 2. After ~1 msec, Na+ channels spontaneously inactivate, blocking further Na+ influx; results from random diffusion of inactive peptide into channel pore opening (similar to that for K + channels) 3. The membrane potential change caused by Na+ influx causes voltage-gated K+ channels to open so K+ ions diffuse freely out of cell & down their steep concentration gradient 4. The decreased Na+ permeability & increased K+ permeability makes potential swing back to a negative value again (~80 mV), approaching that of K+ equilibrium potential 5. The large negative membrane potential causes voltage gated K+ channels to close —> returns cell to resting potential D. Collectively, the changes in membrane potential are called an action potential; the entire sequence of AP changes takes ~5 msec in squid axon & less (<1msec) in myelinated mammalian nerve cell axon E. Na+ channels cannot open for several msec after inactivation, so there is a brief refractory period after an AP during which membrane cannot be restimulated 1. Once negative resting potential is reestablished, the inactivated Na+ ion channels close until they are stimulated to open again by threshold depolarization F. AP greatly changes membrane voltage, but only a minute percentage of ions on the two sides of the membrane are involved in given AP 1. Dramatic changes not caused by changes in Na+ & K+ ion concentrations on two sides of membrane, which are insignificant 2. Instead, they are caused by the movements of charge in one direction or the other that result from the fleeting changes in permeability to these ions 3. Those ions that do cross membrane during AP are pumped back by Na+/K+ ATPase 4. Even if Na+/K+ ATPase is inactivated, a neuron can continue to fire thousands of impulses before the ionic gradients established by the pump are dissipated V. Further characteristics, vital statistics & information about APs A. Certain local anesthetics (procaine & novocaine) act by closing ion channels in sensory cells & neurons & prevent APs; as long as these channels stay closed —> no APs in these cells 1. Affected cells are unable to inform the brain of events occurring at skin or teeth —> no pain B. All-or-None Law - if cell depolarizes to threshold, there is an automatic, full-blown response with no further stimulation, there is no in-between response 1. A subthreshold depolarization is incapable of triggering AP 2. Stronger stimuli cannot produce "bigger" impulses than weaker stimuli C. AP proceeds without more energy expenditure (energy is just needed to establish gradients) – an AP simply results from the flow of ions down their respective electrochemical gradients VI. Propagation of APs as an impulse A. Once AP is initiated, the impulse is propagated down the length of the cell axon (nerve impulse) 1. APs are normally initiated at axon hillock of neuron & move from there to opposite ends of cell (the nerve terminals) 2. Stimulate axon in center as in experiment above, AP moves from stimulus site in both directions B. AP at one site is large enough to cause depolarization above threshold at adjacent site causing sodium channels to open, etc. 1. Large depolarization of AP draws positive ions toward that site on membrane outer surface 2. It also forces the positive ions away from the depolarization site on the inner membrane surface 3. This current flow causes the membrane in the region just ahead of AP to depolarize 4. Since the depolarization accompanying AP is very large, the adjacent membrane region is depolarized to a level greater than threshold value 5. The Na+ channel gates swing open in this adjacent region generating another AP C. Thus, once triggered, a succession of APs travels down the entire length of the neuron to target cell with no loss of intensity (strength); arrives at its target cell with the same strength as at its initiation D. Differences in strengths of stimuli can be detected — how if all impulses exhibit the same strength? – depends on several factors: 1. A stronger stimulus (scalding H2O) activates more nerve cells than weaker stimulus (warm H2O) 2. Stronger stimulus also activates high-threshold neurons that would not fire with weaker stimulus 3. Stimulus strength is also encoded in pattern & frequency by which APs are launched down particular neuron (generally a stronger stimulus generates a larger number of impulses) VII. Speed impulse can travel depends on: A. Diameter of axon - if larger, there is less resistance to local current flow & AP is easier to propagate (an AP at one site can activate adjacent membrane regions more rapidly) 1. Some invertebrates (squid, tube worms) evolved giant axons to escape danger; this approach has limits, since conduction speed increases (slowly) as the square root of the increase in diameter 2. A 16X increase in diameter gives only a 4X rise in speed of conduction; an axon 480 µm in diameter can conduct AP only 4 times faster than one that is 30 µm in diameter B. During vertebrate evolution, an increase in conduction velocity was achieved when the axon became wrapped in a myelin sheath 1. Myelin sheath is formed from nervous system accessory cells (oligodendrocytes in central NS, Schwann cells in peripheral NS) 2. Since it is composed of many layers of lipid-containing membranes, the myelin sheath is ideally suited to prevent the passage of ions across the cell membrane 3. Nearly all of the ion channels of a myelinated neuron reside in the unwrapped gaps (nodes of Ranvier) between adjacent Schwann cells that make up the sheath 4. Thus, nodes of Ranvier are the only sites where APs can be generated; AP at one node triggers AP at next node; impulse jumps from node to node without having to activate intervening membrane 5. Impulse propagation by this mechanism called saltatory conduction; conduction speed along myelinated axon is nearly 120 meters/sec (>20X faster than unmyelinated axon of same diameter) C. Importance of myelination is dramatically illustrated by multiple sclerosis, a disease associated with deterioration of myelin sheath that surrounds axons in various parts of nervous system 1. Manifestations of disease usually begin in young adulthood 2. Victims experience weakness in their hands, difficulty in walking & problems with their vision 3. MS is characterized by progressive muscular dysfunction VIII. Neurotransmission: Jumping the synaptic cleft A. Neurons are linked with their target cells at specialized junctions (synapses) - no direct contact; they are separated from each other by narrow gap of ~20 – 50 nm (gap called the synaptic cleft) 1. Presynaptic cell (PrSC; neuron, sensory cell) conducts impulses toward a synapse, which it must cross to get to postsynaptic cell (PoSC; neuron, muscle, gland cell) 2. PoSC always lies on receiving side of synapse 3. Between axon & muscle cell, synapse is called neuromuscular junction or motor end plate B. Studies done decades ago indicated that a chemical substance is involved in the transmission of an impulse from one cell to another 1. Terminal knobs (tips) of axon branches have been seen in electron microscope to contain large numbers of HO O + H 3 C — C — O — CH 2 — CH 2 — N(CH 3) 3 Acetylcholine (ACh) HO H + C — CH 2 — NH3 OH N orepinephrine C. D. E. F. G. synaptic vesicles that serve as storage sites for chemical transmitters that act on PoSCs 2. They contain neurotransmitters (NTs), like two of the best-studied NTs, acetylcholine & norepinephrine, which transmit impulses to skeletal & cardiac muscles Sequence of initial events in neurotransmission (synaptic transmission) 1. Impulse reaches terminal knob & the accompanying depolarization induces the opening of a number of voltagegated Ca2+ channels in this part of the PrSC plasma membrane 2. Ca2+ ions diffuse from extracellular space into PrSC (Ca2+ ions are usually present at a very low concentration within neurons as in all cells [~100 nM]); [Ca2+] rises >1000-fold near channels) 3. When gates open, Ca2+ ions diffuse from extracellular fluid into terminal knob of neuron, causing [Ca 2+] to rise >1000-fold within localized microdomains near the channels 4. The elevated [Ca2+] triggers rapid synaptic vesicle fusion (one or a few) with PrSC membrane —> neurotransmitters released into synaptic cleft 5. After release from synaptic vesicles, NTs diffuse across narrow gap of cleft & bind selectively to receptor molecules concentrated directly across synaptic cleft on PoSC membrane Such NT molecules can have one of two opposite effects depending on the type of receptor on the target cell membrane to which it binds: 1. Triggers cation-selective channel opening in PoSC membrane; leads to mainly Na+ ion influx & less "-" (more "+") potential; depolarization excites cell; more likely to respond with its own AP 2. Anion-selective channels in PoSC membrane open —> mainly Cl- ion influx & more "-" potential (hyperpolarized), inhibits cell; AP generation less likely (need >Na+ influx to reach threshold) There is also an important class of NT receptors that are not ion channels & thus do not directly affect membrane voltage 1. This other group of receptors are members of a class of proteins called GPCRs 2. When a NT binds to one of these receptors, it can initiate a variety of responses, which often includes the opening of ion channels by an indirect mechanism Most nerve cells in brain receive both excitatory & inhibitory signals from many different PrSCs —> summation of these opposing influences determines if PoSC generates AP or not 1. All terminal knobs of a given neuron release the same NT(s) 2. However, a given NT may excite one particular postsynaptic membrane & inhibit another 3. Example: acetylcholine inhibits heart contractility, but stimulates skeletal muscle contractility In brain, glutamate is primary excitatory NT & gamma-aminobutyric acid (GABA) is primary inhibitory NT 1. A number of general anesthetics, as well as valium & its derivatives, act by binding to GABA receptor & enhancing the activity of the brain's primary "off" switch IX. Actions of drugs on synapses - NTs must have short half-life (a few msec) after release from PrSC to prevent extended effect, PoSC would not recover from this —> thus impulse lasts only a few msec A. NT is thus eliminated from synapse in 2 ways: 1. Enzymes destroy NT in synaptic cleft or 2. Proteins transport NTs back to the presynaptic terminals from which they were released (reuptake) B. Interference with NT destruction or reuptake can have dramatic physiological & behavioral effects C. Example: acetylcholinesterase (AChase) - located in synaptic cleft where it hydrolyzes acetylcholine (ACh); inhibit AChase by nerve gas DFP exposure —> muscles contract violently due to high [ACh] 1. Many drugs act by inhibiting the transporters that sweep NTs out of synaptic cleft 2. A number of widely prescribed antidepressants, including Prozac, inhibit the reuptake of serotonin, a NT implicated in mood disorders D. Example: dopamine (DOP) – released by certain nerve cells in limbic system portion of brain, which contains the brain's pleasure or reward centers 1. Sustained DOP presence in limbic system synaptic clefts produces short-lived euphoric feeling, as well as a strong desire to repeat the activity 2. Cocaine interferes with dopamine reuptake in brain limbic system causing "high", then "crash" 3. With repeated use, the pleasurable effects of cocaine are increasingly reduced, but its addictive properties are enhanced E. Example: amphetamines also act on DOP-releasing neurons; they are thought to stimulate excessive DOP release from presynaptic terminals & interfere with the reuptake of the NTs from synaptic cleft 1. Mice genetically engineered to lack the DOP transporter (DAT; protein responsible for DOP reuptake) show same behavior as normal mice given doses of cocaine or amphetamines 2. Cocaine or amphetamine doses have no additional behavioral effects on mice lacking DAT gene F. The active compound in marijuana (9-tetrahydrocannabinol) acts by a totally different mechanism 1. It binds to cannabinoid (CB1) receptors located on presynaptic terminals of certain brain neurons 2. The binding reduces the likelihood that that these neurons will release NTs 3. CB1 receptors normally interact with compounds called endocannabinoids, which are produced by postsynaptic neurons after depolarization 4. These substances diffuse backwards across the synaptic cleft to the presynaptic membrane where they bind to CB1 receptors, suppressing synaptic transmission 5. CB1 receptors located in many brain areas, including hippocampus, hypothalamus & cerebellum; explains effects of marijuana on memory, motor coordination & appetite, respectively 6. If marijuana increases appetite by binding to CB1 receptors, it follows that blocking these receptors might decrease appetite 7. This line of reasoning has led to development of a new CB1-blocking weight-loss drug (Acomplia) that is likely to be available in the near future G. Significant for Parkinson’s disease studies (results from low DOP levels in certain parts of brain) 1. Drop in DOP levels results in motor dysfunction 2. If we develop drugs that block the DOP transporter, limited DOP released by neurons of Parkinson's patients could be more effective in producing a proper response in PoSCs X. Synaptic plasticity - synapses are more than just connecting sites between adjacent neurons; they are key determinants in the routing of impulses through nervous system and their study is important A. The human brain is thought to contain at least 100 trillion synapses; these synapses act like gates stationed along the various pathways 1. They allow some pieces of information to pass from neuron to neuron or 2. They hold back other pieces of information or reroute them in another direction B. Dynamic changes in synapse structure & activity (synaptic plasticity), a remarkable property, play a role in learning & memory, even though synapses are often perceived as fixed, unchanging structures 1. Synaptic plasticity is particularly important during infancy & childhood when the neuronal circuitry of the brain achieves its mature configuration C. Synaptic plasticity is most readily seen in studies of neurons from the hippocampus (a part of the brain vitally important in learning & short-term memory) 1. The hippocampus is one of the major brain regions destroyed by Alzheimer's disease 2. Repeated hippocampal neuron stimulation over short time period —> synapses connecting them to neighbors are strengthened by long-term potentiation (LTP); may last days, weeks, even longer D. LTP research has focused on NMDA receptor, one of several receptor types that bind the excitatory NT glutamate 1. Glutamate binding to a postsynaptic NMDA receptor, opens an internal cation channel that allows Ca 2+ ion influx into the postsynaptic neuron 2. The Ca2+ influx triggers a cascade of biochemical changes that lead to synaptic strengthening 3. Synapses that have undergone LTP transmit weaker stimuli & evoke stronger responses in PoSCs 4. These changes are thought to play a major role as newly learned information or memories are encoded in brain neural circuits 5. When lab animals are treated with drugs that inhibit LTP, like those that interfere with the activity of the NMDA receptor, their ability to learn new information is greatly reduced E. Synaptic malfunction or dysfunction may cause disease (myasthenia gravis, Parkinson's disease, schizophrenia, & even depression) THE HUMAN PERSPECTIVE: DEFECTS IN ION CHANNELS AS A CAUSE OF INHERITED DISEASE I. Several severe, inherited disorders are caused by mutations in genes that encode ion channel proteins A. Many such disorders affect movement of ions across the membrane of excitable cells (muscle, nerve & sensory cells); reduces the ability of these cells to develop or transmit impulses B. Cystic fibrosis (CF; best-studied & most common inherited ion channel disorder) results from a defect in epithelial cell ion channels 1. On average, ~1 in every 25 people of Northern European descent carries 1 copy of gene that can cause cystic fibrosis; heterozygotes show no symptoms & thus most do not know they are carriers 2. Thus, ~1 in every 2500 infants in Caucasian population (1/25 x 1/25 x 1/4) is homozygous recessive at this locus & born with CF 3. CF affects various organs (intestine, pancreas, sweat glands, reproductive tract) but the respiratory tract usually exhibits the most severe effects 4. CF victims produce a thickened, sticky mucus that is very hard to propel out of airways 5. Affected individuals typically suffer from chronic lung infections & inflammation, which progressively destroy pulmonary function II. The CF gene was isolated in 1989; its sequence & the amino acid sequence of the corresponding protein were determined; the protein was found to be a member of the ABC transporter superfamily A. Like other members in superfamily, polypeptide contains 2 domains situated within lipid bilayer & 2 nucleotidebinding domains (NBDs) that project into cytoplasm B. Unlike other superfamily members, CF protein has a regulatory (R) domain with several serine residues that can be phosphorylated by cyclic AMP (2nd messenger)-activated protein kinase (PKA) C. Called it cystic fibrosis transmembrane conductance regulator (CFTR); were not sure what it did 1. Question was thought to be answered when protein was purified, incorporated into artificial lipid bilayers & shown to act as cyclic AMP-regulated chloride channel, not a transporter D. Subsequent studies have added numerous complications to story as it has been shown that CFTR also: 1. Conducts bicarbonate (HCO3- ions 2. Suppresses the activity of an epithelial Na+ ion channel (ENaC) and 3. Stimulates the activity of a family of epithelial chloride/bicarbonate exchangers III. As CFTR role has gotten more complex, it has become increasingly difficult to establish how a defect in this protein leads to chronic lung infection development; most agree with following general statements: A. Since osmotic H2O movement out of epithelial cells follows salt movement, abnormalities in Cl -, HCO3-,&/or Na+ flux caused by CFTR deficiency leads to drop in fluid volume bathing airway epithelial cells 1. A reduction in volume of surface liquid & a resulting increase in the viscosity of secreted mucus impairs function of cilia responsible for pushing bacteria out of respiratory tract B. Among the bacteria that infect airways of CF patients, Pseudomonas aeruginosa is most prevalent & destructive species 1. This bacterium is rarely seen in airways of individuals suffering from other types of lung disease 2. It is not certain why CF patients are so susceptible to it 3. Studies show that P. aeruginosa binds to extracellular end of the CFTR protein & it is thought that this attachment may lead to ingestion & destruction of bacterium by epithelial cells 4. Individuals who lack CFTR protein in plasma membrane, as do many CF patients, may be unable to clear the bacterium from their respiratory tract IV. In past decade, researchers have identified >800 different mutations that give rise to CF A. However, nearly 70% of alleles responsible for CF in U. S. contain the same genetic alteration (F508); they are all missing 3 base pairs of DNA that encode a phenylalanine at position 508 1. Position 508 is within one of the domains of the CFTR polypeptide 2. CFTR polypeptides lacking this particular amino acid fail to be processed normally within the endoplasmic reticulum membrane & thus never reach the epithelial cell surface 3. CF patients homozygous for the F508 allele completely lack the CFTR channel in their plasma membranes; they have a severe form of the disease B. Other CF patients with less severe forms have mutant alleles that encode a CFTR that is able to reach the surface of cells but mediates a reduced anion conductance 1. The mildest forms are characterized by infertility, with little or no damage to major organs C. According to one estimate, the F508 mutation had to have originated >50,000 years ago to have reached such a high frequency in the population 1. The fact that the CF gene has reached this frequency suggests that heterozygotes may receive some selective advantage over those lacking a copy of the defective gene 2. It has been proposed that CF heterozygotes may be protected from the effects of cholera, a disease characterized by excessive fluid secretion by the wall of the intestine 3. A problem with this idea is that there is no record of cholera epidemics in Europe until the 1820s 4. Another proposal suggests that heterozygotes are protected from typhoid fever since the bacterium responsible adheres poorly to an intestine wall having a reduced number of CFTR molecules V. Ever since the isolation of the CF gene, the development of a cure by gene therapy (replacement of the defective gene with the normal version) has been a major goal of researchers A. CF is a good candidate for gene therapy since the worst symptoms of the disease result from defective activities of epithelial cells that line the airways B. These cells are accessible to substances that can be delivered by inhalation of an aerosol; clinical trials have begun using several different types of delivery systems – examples are adenovirus & liposomes C. Example - the normal CFTR gene is incorporated into the DNA of a defective adenovirus (a virus that normally causes upper respiratory tract infections) 1. Recombinant virus particles are then allowed to infect the cells of the airway, delivering the normal gene to the genetically deficient cells 2. The primary disadvantage in using adenovirus is that viral DNA (& the normal CFTR gene) does not get integrated into infected host cell chromosomes; thus it must be readministered frequently 3. Also, the virus induces an immune response that eliminates the virus & leads to lung inflammation 4. Researchers are hesitant to employ viruses that do integrate their genomes for fear of initiating the formation of cancers D. Example – DNA encoding the normal CFTR gene has been linked to positively charged liposomes that can fuse with plasma membranes of airway cells, delivering their DNA contents into cytoplasm 1. Lipid-based delivery has an advantage over viruses in being less likely to stimulate a destructive immune response in the patient after repeated treatments 2. It has the disadvantage of being less effective in achieving genetic modification of target cells E. Early clinical trial results are encouraging especially those relying on liposomes, which have not caused any significant side effects in patients after a single treatment 1. In most patients, the normal gene enters a sufficient percentage of airway cells to at least partially correct the underlying Cl- defect & to reduce the adherence of bacteria to lung tissue 2. The development of more effective DNA delivery systems, which can genetically alter a greater percentage of airway cells, would probably be required for effective treatment of CF 3. It also remains to be shown whether repeated administration of the normal gene can control the disease over long periods of time EXPERIMENTAL PATHWAYS: THE ACETYLCHOLINE RECEPTOR I. Claude Bernard (foremost physiologist of 19th century; once a pharmacist & aspiring playwright) – interested in elucidating the mechanism by which nerves stimulate skeletal muscle contraction A. Used curare (highly toxic drug isolated from tropical plants & used by native S. American hunters to make poisonous darts) 1. Found that curare paralyzed skeletal muscle without interfering with either the ability of nerves to carry impulses to the muscle or the ability of the muscle to contract on direct stimulation 2. He concluded that curare somehow acted on the region of contact between the nerve & muscle B. John Langley (physiologist, Cambridge Univ.) – confirmed & extended Bernard's conclusion 1. Studied the ability of nicotine (also derived from plants) to stimulate contraction of isolated frog skeletal muscles & the effect of curare in inhibiting nicotine action 2. 1906 – Langley concluded that nerve impulse does not pass from nerve to muscle by electric discharge but instead by secretion of a special substance on the end of the nerve 3. Proposed that the chemical transmitter was binding to a receptive substance on muscle cell surface at the same site that bound nicotine & curare – farsighted conclusions C. Otto Loewi (Austrian-born physiologist, 1921) – confirmed Langley's suggestion that nerve-muscle stimulus was transmitted by a chemical with ingenious experiment that came to him in a dream 1. Vertebrate heart rate is regulated by input from 2 opposing (antagonistic) nerves; Loewi isolated a frog heart with both nerves intact 2. Stimulated inhibitory (vagus) nerve —> chemical released from heart prep into salt solution, which drained into medium bathing second isolated heart —> second heart's rate slowed dramatically 3. Second heart acted like its vagus (inhibitory) nerve was activated; called substance that inhibited frog's heart "Vagusstoff"; soon showed its chemical/physiological properties matched acetylcholine 4. Concluded acetylcholine (ACh) was substance released by tips of nerve cells in vagus nerve D. David Nachmansohn (neurophysiologist at the Sorbonne; 1937) – visited Paris World's Fair & observed several living electric fish of species Torpedo marmarota 1. These rays have electric organs that deliver strong shocks (40 – 60 volts) that can kill potential prey 2. Studied acetylcholinesterase (destroys ACh after its release from motor nerve tips) & realized electric organs were derived from modified skeletal muscle tissue & asked for fish to study 3. He found that the electric organ was an extraordinarily rich source of acetylcholinesterase & also a very rich source of nicotinic acetylcholine receptor (nAChR) 4. nAChR is present on postsynaptic membranes of skeletal muscle cells & binds ACh released from motor nerve tips; nAChR is called nicotinic since it is activated by nicotine as well as by ACh 5. This contrasts with muscarinic ACh receptors of parasympathetic nerve synapses (activated by muscarine, but not nicotine; inhibited by atropine, but not curare) 6. Smokers' bodies get accustomed to high nicotine levels & experience withdrawal symptoms when they stop smoking, since postsynaptic neurons possessing nAChRs are not stimulated at usual level 7. Recent studies have shed considerable light on the relationship between nAChR structure & nicotine addiction II. nAChR is integral membrane protein & wasn't isolated until 1970s, since isolation techniques were not perfected until then A. The ideal nAChR assay was a compound that bound selectively & tightly to it (found in 1963 by Chen-Yuan Lee et al. at National Taiwan Univ.), -bungarotoxin (found in venom of a Taiwanese snake) 1. -bungarotoxin causes paralysis by binding tightly to nAChR on skeletal muscle cell postsynaptic membrane, blocking response of muscle to ACh 2. Using labeled -bungarotoxin to assay for nAChR, electric organs as a source & a detergent to solubilize membrane proteins, many researchers isolated acetylcholine receptors in the 1970s B. In one study, membranes containing nAChR were isolated by homogenizing the electric organs in a blender & centrifuging the suspension to pellet membrane fragments 1. Triton X-100, a nonionic detergent, gave the best results 2. It has a long hydrophobic portion that can replace the phospholipids that normally surround the hydrophobic portions of membrane proteins & a hydrophilic end that is soluble in water 3. Unlike most detergents, which bear a negatively charged carboxyl group, Triton X-100 lacks a charge (nonionic) & is thus less disruptive of protein structure 4. Proteins were extracted from membrane fragments with Triton X-100 & mixture was passed through column containing tiny beads coated with synthetic compound, whose end looks like ACh 5. 2 proteins with binding sites for ACh, nAChR & acetylcholinesterase (AChE) stuck to the beads 6. The remaining 90% of protein in the extract failed to bind to the beads & simply passed through 7. Next, a solution of 10-3 M flaxedil was passed through the column; it selectively removed nAChR from beads leaving AChE behind 8. nAChR measured by -bungarotoxin was purified >150-fold in one step (affinity chromatography) C. The structure of the nAChR receptor – Arthur Karlin (Columbia Univ.) determined that nAChR is a pentamer (a protein consisting of 5 subunits) 1. Each receptor contained 2 copies of a subunit called & one copy each of , & subunits 2. They could be distinguished by extracting membrane proteins in Triton, purifying nAChR by affinity chromatography & then doing SDS-PAGE separating the proteins according to size III. Purified receptor was found to act as both a site for ACh binding & a channel for the passage of cations A. Jean-Pierre Changeux (Pasteur Inst., Paris) – postulated years earlier that binding of ACh to receptor caused conformational change that opened ion channel within protein 1. Na+ ion influx through the channel could then lead to depolarization of membrane & activation of muscle cell 2. During last half of 1970s, Changeux et al. incorporated purified nAChR molecules into artificial lipid vesicles 3. Using vesicles containing various concentrations of labeled Na+ & K+ ions, they showed that binding of ACh to receptors in lipid bilayer initiated a flux of cations through membrane B. The pure ACh protein contains all the structural elements needed for chemical transmission of an electrical signal — an ACh binding site, an ion channel & a mechanism for coupling their activity IV. During past decade, researchers have focused on determining nAChR structure & mechanism by which it becomes concentrated within one small part of muscle membrane, the part adjacent to motor neuron tip A. Scientists used purified genes, amino acid sequence determination & site-directed mutagenesis to identify specific parts of polypeptides that span membrane, bind neurotransmitter or form ion channel B. Electron microscopy was employed in one approach – nAChR first seen in EM of electric organ membranes 1. Receptors appeared ring-shaped with diameter of 8 nm & central channel of 2 nm diameter & protruded out from lipid bilayer into external space C. Nigel Unwin et al. (Med. Research Coun., England) – used mathematical analysis of frozen electric organ membrane electron micrographs; able to analyze nAChR structure as it is in its native lipid environment 1. He described the arrangement of the 5 subunits around central channel 2. Ion channel consists of narrow pore lined by a wall made of 5 inner (M2) -helices, one from each of the surrounding subunits 3. The gate to the pore lies near the middle the membrane, where each of the M2 -helices bends inward to form a kink in the unactivated receptor 4. The side chain of a leucine residue projects inward from each kink; the leucine residues from the 5 helices form a tight hydrophobic ring that prevents the ions from crossing the membrane 5. The gate opens after binding of 2 ACh molecules, one per subunit, each one binding to a site located within a pocket of an subunit D. Unwin then studied the changes in the nAChR during channel opening 1. nAChR-rich membrane preparations were applied to a support grid that was allowed to fall into a bath liquid nitrogen cooled ethane, which freezes the membranes 2. ~5 msec before they reached the freezing bath surface, the grids were sprayed with a solution of ACh, which bound to the receptors & triggered conformational change that was required to open channel 3. Compared electron micrographs of nAChRs trapped in open vs. closed state; found that ACh binding triggers small shape change in receptor subunit extracellular domains near the 2 ACh binding sites 4. This conformational change is propagated down the protein, causing a small (15°) rotation of the M2 helices that line the ion conduction pore 5. The rotation of these inner helices breaks apart the hydrophobic gate, which allows the Na+ ions to enter the cell QUESTÕES 1. Está a estudar uma molécula envolvida no transporte. Essa molécula parece ligar-se à molécula a ser transportada. Durante o transporte não há gasto de energia. A adição de outra molécula, que se assemelha bastante à molécula normalmente transportada, inibe o transporte. Um aumento na concentração da molécula que normalmente é transportada, aumenta a taxa de transporte, mesmo na presença de concentrações constantes da molécula inibidora. Que tipo de transporte é referido e de que tipo é a molécula inibidora? 2. Se estivesse a estudar o impulso nervoso in vitro e se adicionasse ao sistema oubaína, um inibidor da ATPase de Na+ K+, seria de esperar que a transmissão do impulso nervoso parasse imediatamente? 3. A taxa de condução de impulso nervoso é diferente em dois neurónios. O primeiro neurónio apresenta uma taxa de condução substancialmente maior (cerca de 20 vezes) do que o Segundo. Ambos apresentam axónios com o mesmo diâmetro. Qual a possível explicação para as diferenças observadas na taxa de condução de impulso nervoso? 4. Quais são as possíveis explicações para a tendência dos doentes com fibrose quistica desenvolverem infecções pulmonares crónicas? 5. Quais os dois eventos separados necessários para abrir o canal CFTR? 6. Os pacientes com a forma mais severa da fibrose quística não conseguem realizar o transporte de qualquer ião cloreto, enquanto nos casos menos severos ocorre o transporte de alguns iões cloreto. Qual a explicação para este facto? 7. A maioria dos alelos responsáveis pela fibrose quística nos Estados Unidos da América contém a mesma alteração genética, designada ∆F508. Qual é o significado desta alteração e de que modo leva ela ao desenvolvimento da fibrose quística? 8. A mutação ∆F508, que origina a fibrose quística, está presente em níveis muito altos na população caucasiana. Pensa-se que, para atingir os níveis actuais, terá tido origem há mais de 50000 anos. Os indivíduos homozigóticos para o alelo recessivo mutante têm uma saúde extremamente depauperada. Então, o que poderá explicar o facto de este alelo ter sido mantido ao longo de tantas gerações até atingir níveis tão altos? 9. Diga onde são sentidos os principais sintomas da fibrose quística e descreva dois potenciais tratamentos para estes sintomas, referindo qual deles deverá ser o mais vantajoso! 10. Isolou-se um coração de rã mantendo os dois principais nervos controladores intactos e a ele ligados. O nervo excitatório foi estimulado e a solução salina que banhava o coração foi transferida para outro coração cujos dois nervos tinham sido retirados. a) O que terá acontecido a este coração? b) O que aconteceria se adicionasse ao coração inicial uma solução contendo acetilcolinesterase, antes do nervo vagus ser estimulado? 11. Porque é que fumar de forma prolongada torna mais provável o surgimento de doenças cardíacas e porque é que os fumadores que param de fumar sentem sintomas de abstinência? 12. Foram preparadas vesículas membranares artificiais contendo receptores de acetilcolina. Foram banhadas numa solução com iões K+ e Na+ marcados. O que acontece se for adicionada acetilcolina à mistura? 13. A figura 4,35 mostra o que acontece a um eritrócito quando é colocado em soluções de diferentes concentrações. Considere que um eritrócito com uma osmolaridade interna de 0,15 M NaCl é colocado numa solução com uma concentração de 0,35 M NaCl. a) o que acontece ao eritrócito? b) que palavra descreve o interior da célula relativamente à solução que a rodeia? c) que palavra descreve a solução que rodeia o glóbulo vermelho em relação à sua concentração interna? d) se a solução que rodeia o eritrócito tivesse uma concentração de 0,15 M NaCl, que palavra a descreveria? 14. Como retratado na figura 4.45, por que razão o transporte mediado por proteínas atinge um valor máximo enquanto a taxa da difusão simples aumenta linearmente? 15. De acordo com a figura 4,49 como é que a glucose se move do lúmen intestinal para a corrente sanguínea? 16. O que acontece se estimular um axónio, como o da figura 4.53, a meio da sua extensão? 17. De acordo com a figura 4.54, que tipo de condução de impulso nervoso é proporcionado pela bainha de mielina? 18. Qual o papel da digitalina na contração muscular?