Species abundance patterns
advertisement

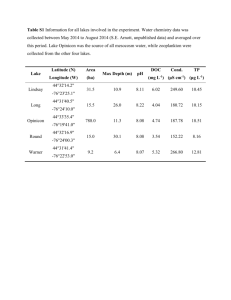
Community Ecology Species abundance patterns Outline: 1. Most species are rare, few are common. A. Overview of dominance-diversity plots (species-abundance distributions) B. Some explanations for most rare/few common pattern: i. Geometric series (sequential niche breakage model) ii. Log series iii. Broken-stick model (simultaneous niche breakage model) iv. Log-normal model C. Are these explanations based in biology or in statistics? 2. Abundance generally highest in the center of a species’ range, decreasing towards the periphery 3. Common species tend to be widespread, rare species tend to be restricted in distribution. A. Some explanations: i. Sampling artefact ii. Resource requirements iii. Density-dependence iv. Vital rates v. Core-satellite hypothesis (Hanski 1982) Terms/people: abundance dominance-diversity plot (species-abundance distribution, Whittaker plot, rank-abundance plot) geometric series (Motomura) broken-stick model (MacArthur) log series (Fisher et al.) log-normal (Preston) sequential-breakage model simultaneous-breakage model Hanski (core-satellite hypothesis) Central Limit Theorem veil line (Preston) abundance surrogates for abundance Three general patterns regarding abundance: 1. Species have unequal distributions: most species are relatively rare, few are common. (Rarity based on population size, range, endemicity, habitat specialization, etc.; see Rabinowitz 1981, Gaston 1994.) dominance-diversity plots (a.k.a. species-abundance distributions, Whittaker plots, or rank-abundance plots) There are over 2 dozen forms of species-abundance distributions (McGill 2011), each resulting from a different process (considerable debate about whether a given species-abundance distribution is ecological or statistical in origin); 4 especially common: 1) geometric series (a.k.a. sequential breakage model) - Motomura (1932); very steep; sequential niche breakage: the probability of any niche fragment being subdivided is independent of its size. Ecological dominance effect: early arriving species can pre-empt resources. An early arriver can become dominant in abundance and thereby limit later arrivers (resulting in a steep dominance-distribution plot). Occurs when species arrive at an unsaturated habitat at regular intervals of time, occupying fractions of remaining niche hyperspace. Tends to hold for communities in settings where there is only one or a few dominating environmental factors (e.g. light limitation for understory plants, or soil nutrient limitation for desert microbes). Tends to be seen more in communities with low richness and early seral stages. 2) log series (a.k.a. logarithmic series) - Fisher et al. (1943); less steep; occurs when species arrive at an unsaturated habitat at random intervals of time. Like the geometric series, tends to hold for communities in settings where there is only one or a few dominating environmental factors. Tends to be seen more in communities with low richness. 3) broken stick (a.k.a. simultaneous breakage model, random niche-boundary model) MacArthur (1957); more shallow (almost flat), with most species almost equally abundant; simultaneous niche breakage: resource is partitioned more equitably and no severe dominance is possible (therefore, a species’ abundance is proportional to its resource use), resulting in a lesssteep dominance-diversity plot than is seen with the two previous mechanisms. Typically found in narrowly defined communities of closely related species. Question for the class: why do you think that closely related groups are more likely to fit the broken-stick model than unrelated groups of species? 4) log-normal - Preston (1948, 1962); see also Sugihara (1980); a few spp. are very common, most species VERY rare (i.e., highly right-skewed), often stair-stepped, especially when divided into octaves; veil line illustrates importance of sample size; is the most common distribution (but see #5), assumed as null for undisturbed communities (disturbance drives more towards geometric series); but is this distribution a function of large samples and/or the Central Limit Theorem? (See http://vimeo.com/75089338 for an explanation of what the CLT is.) The log-normal is usually considered the undisturbed default for most communities; it is arguably the most commonly observed of the distributions (Ulrich et al. 2010). As a community is disturbed, it tends towards the geometric series. See Kevan et al. (1997) for an example on bees exposed to pesticides in Canadian blueberry fields. We will go over each of these in class in some detail, after which you should be familiar with what the shape of each of the models above looks like (be able to determine, from a dominance-diversity plot, which of the models is the appropriate model) and how they differ in terms of mechanism. For more information, I highly recommend Magurran (2004). We will not fit or test models (a process that is quite involved); for worked examples, I highly recommend Krebs (1999). Grad students will have a worksheet to do on these models. 2. Abundance generally highest in the center of a species’ range, decreasing towards the periphery (Grinnell 1922) Field data on birds by Emlen et al. 1986 confirms this; ditto for Bock and Ricklefs 1983 (Christmas Bird Count data), who found significant positive correlation between range size and mean local abundance. 3. Common species are widespread, rare species are restricted in distribution - Hanski 1982 called this a "general rule" of nature Pattern is not universal, however: Gaston 1996 in a review of 90 studies, found 80% positive, 5% negative, and 15% non-significant For exceptions, see e.g. Fuller 1982 (British birds), Gaston and Lawton 1989 (insects) Possible explanations for this pattern: 1. Sampling artefact 2. Function of resource requirements 3. Density-dependent habitat selection 4. Vital rates 5. Core-satellite hypothesis Next time: Community assembly and null models References: Bock, C.E., and R.E. Ricklefs. 1983. Range size and local abundance of some North American songbirds: a positive correlation. Am. Nat. 122:295-299. Clarke, K.R. 1990. Comparisons of dominance curves. J. Exp. Mar. Biol. Ecol. 138:143-157. Cotgreave, P., and P.H. Harvey. 1994. Evenness of abundance in bird communities. Journal of Animal Ecology 63: 365-374. Emlen, J.T., M.J. DeJong, M.J. Jaeger, T.C. Moermond, K.A. Rusterholz, and R.P. White. 1986. Density trends and range boundary constraints of forest birds along a latitudinal gradient. Auk 103:791-803. Fisher, R.A., A.S. Carbet, and C.B. Williams. 1943. The relation between the number of species and the number of individuals in a random sample from an animal population. J. Anim. Ecol. 12:42-58. Fuller, R.J. 1982. Bird Habitats in Britain. T & A D Poyser, Calton, UK. Gaston, K.J. 1994. Rarity. Chapman and Hall, London, UK. Gaston, K.J. 1996. The multiple forms of the interspecific abundance-distribution relationship. Oikos 76:211-220. Gaston, K.J., and J.H. Lawton. 1989. Insect herbivores on bracken do not support the coresatellite hypothesis. Am. Nat. 134:761-777. Grinnell, J. 1922. The trend of avian populations in California. Science 56:671-676. Hanski, I. 1982. Dynamics of regional distribution: the core and satellite species hypothesis. Oikos 38:210-221. Hanski, I., and M. Gyllenberg. 1997. Uniting two general patterns in the distribution of species. Science 275:397-400. He, F.L., and K.J. Gaston. 2000. Estimating species abundance from occurrence. Am. Nat. 156:553-559. Kevan, P.G., C.F. Greco, and S. Belaoussoff. 1997. Log-normality of biodiversity and abundance in diagnosis and measuring of ecosystem health: pesticide stress on pollinators on blueberry heaths. J. Appl. Ecol. 34:1122-1136. Krebs, C.J. 1999. Ecological Methodology (2nd ed.). Addison-Welsey Educational Publ., Inc., Menlo Park, CA. [highly recommended] Kunin, W.E. 1998. Extrapolating species abundance across spatial scales. Science 281:15131515. Kunin, W.E., S. Hartley, and J.J. Lennon. 2000. Scaling down: on the challenge of estimating abundance from occurrence patterns. Am. Nat. 156:560-566. Lawton, J.H. 1993. Range, population abundance and conservation. Trends Ecol. Evol. 8:409413. Loreau, M. 1992. Species-abundance patterns and the structure of ground-beetle communities. Ann. Zool. Fenn. 28:49-56. MacArthur, R.H. 1957. On the relative abundance of bird species. Proc. Natl. Acad. Sci. USA 43:293-295. Magurran, A.E. 2004. Measuring Biological Diversity. Blackwell Publishing, Malden, MA. [highly recommended] Magurran, A.E., and P.A. Henderson. 2003. Explaining the excess of rare species in natural species abundance distributions. Nature 422:714-716. Maina, G.G., and H.F. Howe. 2000. Inherent rarity in community restoration. Conservation Biology 14:1335-1340. May, R.M. 1975. Patterns of species abundance and diversity. Pp. 881-120 in: Ecology and Evolution of Communities (M.L. Cody and J.M. Diamond, eds.). Harvard University Press, Cambridge, MA. McGill, B.J. 2011. Species abundance distributions. Pp. 105-122 in: Biological Diversity: Frontiers in Measurement and Assessment (A.E. Magurran and B.J. McGill, eds.). Oxford University Press, New York, NY. Preston, F.W. 1948. The commonness and the rarity of species. Ecology 29:254-283. Preston, F.W. 1962. The canonical distribution of commonness and rarity of species. Ecology 43:185-215. Rabinowitz, D. 1981. Seven forms of rarity. Pp. 205-217 in: Biological Aspects of Rare Plant Conservation (H. Synge, ed.). Wiley, Chichester, UK. Sugihara, G. 1980. Minimal community structure: an explanation of species abundance patterns. Am. Nat. 116:770-787. Tokeshi, M. 1990. Niche apportionment or random assortment: species abundance patterns revisited. J. Anim. Ecol. 59:1129-1146. Tokeshi, M. 1996. Power fraction: a new explanation for species abundance patterns in speciesrich assemblages. Oikos 75:543-550. Ulrich, W., M. Ollik, and K.I. Ugland. 2010. A meta-analysis of species-abundance distributions. Oikos 119:1149-1155. Wilson, J.B. 1991. Methods for fitting dominance/diversity curves. J. Veg. Sci. 2:35-46.