1304 Exam 3 Review - FacultyWeb Support Center
advertisement

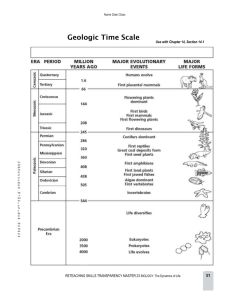
Historical Geology Exam III Review The Early Paleozoic: Cambrian, Ordovician, and Silurian Physical Characteristics: Six major continents existed at the beginning of the Paleozoic Era; four were located near the paleo-equator. During the Early Paleozoic (Cambrian to Silurian), Laurentia (North America) was moving northward and Gondwana was moving to a south polar position as indicated by the tillite deposits. Most continents consisted of two components: 1.) a relatively stable craton over which epieric seas transgressed and regressed, surrounded by 2.) mobile belts (platform materials) in which mountains were built. Collisions between continental masses deformed the platform materials creating Arches and Basins. The arches forming elongated island masses and the basins forming elongated seas (epieric). The geologic history of North America can be divided into cratonic sequences that reflect eustatic (worldwide) sea level changes. (see cratonic sequence chart) The Sauk Sea was the first major transgression onto the craton. At its maximum, it covered the craton except for parts of the Canadian Shield and the Transcontinental Arch, forming a series of large, northeast-southwest trending islands. The Tippecanoe sequence began with the deposition of an extensive sandstone over the exposed and eroded Sauk landscape. During Tippecanoe time extensive carbonate deposition took place. In addition, large barrier reef complexes of Rugosa and Tabulate corals enclosed basins, resulting in evaporite deposition within these basins. The eastern edge of North America was a stable carbonate platform during Sauk time. During Tippecanoe time, an oceanic-continental convergent plate boundary formed, resulting in the Taconic orogeny, the first of several orogenies to create the Appalachian Mountains. The newly formed Taconic Highlands shed sediments into the western epieric sea, producing the Queenstone Delta, a clastic wedge. Early Paleozoic-age rocks contain a variety of mineral resources including building stone, limestone for cement, silica sands, hydrocarbons, evaporates, and iron ores. Paleozoic Orogenies of Laurentia 1.) Taconic – Orovician – east coast - First uplift of the Appalachian Mountains 2.) Acadian – Devonian – upper east coast - Uplift of the northern Appalachians (Maine, New York) 3.) Antler - Devonian – west coast – First beginnings of the uplift of the subsequent Rocky Mountians 4.) Quachita – Mississippian/Pennsylvanian – southern North America (Llano, Texas northeast through Arkansas area) – resulted in the Llano Uplift of central Texas, the Quachita Mountains of Oklahoma and Arkansas. 5.) Alleghenian – Permian – east coast – further uplift of Appalachians. The Late Paleozoic: Devonian, Mississippian, Pennsylvanian, and Permian During the Late Paleozoic, Baltica and Laurentia collided, forming Laurasia. Siberia and Kazakhstania collided and were finally sutured to Laurasia. Gondwana moved over the South Pole and experienced several glacial-interglacial periods, resulting in eustatic sea level changes and transgressions and regressions along low lying craton margins. Laurasia and Gondwana underwent a series of collisions beginning in the Mississippian/Pennsylvanian. During the Permian, the formation of Pangea was completed. Surrounding the supercontinent was a global ocean – Panthalassa. The Late Paleozoic history of the North American craton can be deciphered from the rocks of the Kaskaskia and Absaroka cratonic sequences. A common Late Paleozoic rock series is the Cyclothem. This is a series of non-marine sediments overlain by a clay (subsoil), overlain by a coal layer, overlain by younger marine lithofacies sediments. This is interpreted at the bottom as terrestrial “swampy” conditions of the Carboniferous, followed by an eustatic rise in the sea level inundating the swampy lowlands. This changing sea level was caused by the adancing and retreating of the Gondwanan ice sheets in the polar regions. The basal beds of the Kaskaskia sequence that were deposited on the exposed Tippecanoe surface consisted of either sandstones derived from the eroding Taconic highlands, or carbonate rocks. Most of the Kaskaskia sequence is dominated by carbonates and associated evaporates. The Devonian Period was a time of major reef building in western Canada, southern England, Belgium, Australia, and Russia. A persistent and widespread black shale was deposited over a large area of the craton during the Late Devonian and Early Mississippian, with the Mississippian being dominated for the most part by carbonate deposition. Cratonic mountain building, specifically the Ancestral Rockies, occurred during the Pennsylvanian Period, resulting in thick nonmarine detrital rocks and evaporates in the intervening basins. By Early Permian, the Absaroka Sea occupied a narrow zone of the south-central craton. Here, several large reefs and associated evaporates developed. By the end of the Permian Period, the Absaroka Sea had retreated from the craton. This Permian regression of the Absaroka Sea left behind massive Permian Red-Beds consisting of oxidized marine sediments. Since Pangea also had formed, this caused a drying out of the central craton creating very arid, and desert-like conditions on the middles of the cratons. With much of the coastlines destroyed at this time, the reduction of the “coastal effects” of precipitation had ceased. Environmental pressures were HIGH resulting in the Permian Mass Extinction. Approximately 95% of life on earth perished during the Permo-Triassic Extinction! The Cordilleran Mobile Belt – was the site of the Antler orogeny, a minor Devonian orogeny during which deep water sediments were thrust eastward over shallow water sediments. During the Pennsylvanian and Permian Periods, mountain building occurred in the Quachita mobile belt. This tectonic activity was partly responsible for the cratonic uplift that took place in the southwest, resulting in the Ancestral Rockies. During the Paleozoic Era, numerous terranes and microplates such as Avalonia, existed and played an important role in the formation of Pangea. Late Paleozoic –aged rocks contain a variety of mineral resources including petroleum, coal, evaporates, silica sand, lead, zinc, and other metal deposits. Life of the Lower Paleozoic Era Epieric Seas: broad, shallow, warm, nutrient seas that were formed by continental fringe collisions that created "Arches and Basins". These basins were filled with seawater as they formed creating almost an "incubator" effect on marine life forms. As the tilt of the axis of the earth wobbled throughout geologic time causing growth of the ice caps, followed by their melting, these Epieric seas would fill and drain accordingly. Whenever there was times of ice cap buildup, there was a simultaneous "Eustatic" (= "worldwide") sea level drop draining the Epieric seas causing extinctions and intense environmental pressures on the life forms. Whenever the ice caps melted, there was an eustatic rise in sea level resulting in the epieric seas to again fill and the incubator effect would cause life forms to flourish. The subsequent rise and fall of the sea level is recorded in the geologic record as series of changing marine lithofacies upon the continents called Cratonic Sequences. These changes (i.e. the rise and fall of the Sauk Cratonic) were the driving forces of life evolution, molding and establishing ecosystems worldwide. (see Cratonic Sequence Chart) The Cambrian Explosion: the adaptive radiation of the marine life of the Cambrian due to not only the development of the epieric seas, but also on the rise of gene mutations seen in Cambrian marine life that utilized calcium carbonate and calcium phosphate for the first time as body parts: bone, shells, & teeth. Hence, the beginning of the Phanerozoic Eon ("visible life" meaning that there began to be abundant fossils in the geologic record). This increased the competition of the species worldwide resulting in the so-called "Cambrian Explosion of Life". This also is what separates the Edicarian (Late Proterozoic 700MYA - 570 MYA) "soft bodied metazoan" fossil assemblages from the "hard shelled" Cambrian assemblages. Life forms beginning in the Cambrian: 50% of all fossils of the Cambrian are trilobites - Phylum Arthropoda, Class Trilobita. All Periods of the Paleozoic have trilobite Index Fossils. Archeocyathids - little known Coral-like? Sponge like? Organisms that flourished for a while but became extinct in the Early Paleozoic. Brachiopods - Phylum Brachiopoda, Class Articulata & Class Inarticulata became extremely abundant, even throughout the Paleozoic. Most Cambrian life forms fall into the preceding three groups: Trilobites, Brachiopods, and Archeocyathids. Phylum Mollusca - Classes Gastropoda (snails), Bivalvia (clams), Cephalopoda (nautilus, octopus, squid) also were abundant starting in the Cambrian throughout most of the Paleozoic. Early Echinoderms- Phylum Echinodermata Class Crinoidea (et al) were established. Many other bizarre forms of life evolved but became extinct in the Early Paleozoic The Burgess Shale: Discovered in the Canadian Rockies of British Colombia by Charles Doolittle Walcott in 1910 is world renown for excellent fossil evidence of the “Cambrian Explosion”. Neotony: The retention of larval characteristics into the sexually reproductive adult form. This is a major driving force of evolutionary change as in: Echinoderm Bipinnaria Larvae: This is one of the best examples of neotony known. The bilaterally symmetrical echinoderm larvae is the stock group of the Phylum Chordata. By remaining nektonic, some larvae developed a notochord for support and locomotion. This created the Phylum Chordata. By the addition of mutations within this group, namely the absorption of calcium salts from the sea, allowed for the formation of internal bony supports protecting the spinal cord. This created the first member of the Subphylum Vertebrata. First Vertebrate Organism: The Agnathans – These were the first vertebrate fishes of the Cambrian. They in turn became the stock group that gave rise to all subsequent vertebrate groups. Ontogeny Recapitulates Phylogeny – Change within an organism’s lifetime (ontogeny) retells (recapitulates) its evolutionary history (phylogeny). The Rise of the Chordates and Vertebrates: The Cambrian seas were the site of another adaptive radiation of the vertebrate fish organisms. From the “jawless” Agnathan (Ostracoderm) stock, there arose the following: Placoderms – “armored fishes” - Silurian to Mississippian Acanthodii – Silurian to Permian Chondrichthyes – “sharks, skates, and rays” - Devonian to Recent Osteichthyes – “bony fishes” - Devonian to Recent The Ordovician This is marked by a major extinction due to the regression of the Sauk. Bryozoans and Graptolites began to inhabit the seas. But, with the rise of the Tippecanoe Cratonic Sequence, life again began to rebound. Evidence shows that the climate worldwide began to stabilize in the Ordovician. Corals (Rugosa and Tabulata) began to inhabit the warm shallow seas. The Silurian First coral reefs appeared with their lagoons mimicking epieric seas. This production of calm, warm, nutrient-rich lagoons, along with the production of numerous niches and habitats was their major importance. Know the cross-section of a coral reef: land, lagoon, reef core, reef tallus, fore-reef, and back-reef The first land plants (modified Chlorophyte Algae w/ waxy cuticle and vascular tissue) altering the climate and creating a more oxygen rich atmosphere. Ozone levels began to rise as lowland forests spread across the continents. Eumeric flora in Laurasia and Glossopteris sp. seed fern forests in Gondwana. (see evolution of Plants page). The rise of terrestrial plants provided a food supply that was soon exploited. Eurypterus sp. "sea scorpions" were the first animal life to crawl onto land and it is thought that they were exploiting the terrestrial plant diversity as a food supply. This lead to Eurypterus sp. giving rise to terrestrial arthropods (insects, et al.) creating yet another food supply to become exploited. The Taconic Orogeny (convergence of N. America and Africa) was first uplift of the Appalachians, diverting the jet stream and altering the weather patterns resulting in a very long "swampy" climate of most of the equatorial continents. Evolutionary Relationships of Terrestrial Plants to Algal Ancestors The fossil evidence for the evolution of modern plants supports the idea that all terrestrial plants evolved from a common ancestor, namely marine algae related to members of the Phylum Chlorophyta (the Green Algae). If one compares the physiology of Chlorophytan cells to modern terrestrial plants, one is able to see many similarities: a cellulose cell wall, storing excess food as starch, similar pigments, etc. The two main differences between Green Algae and Modern Plants are the absence of a “Cuticle” and “Vascular Tissue” in the Green Algae. It is thought that the “Cuticle” (which is a waxy, water-proofing coating found on the outside of plant epidermis) evolved through natural selection from a group of inter-tidal Chlorophytan-type seaweeds that were thrown into intermittent tidal pools and subjected to desiccation. Having the ability to form a protective coat, this group of algae was able to exploit a new habitat in the semi-dry environment of the beach- fronts. This stock group was also subjected to natural selection by the competition to survive in this new “frontier”. From this group arose individuals that had series of internal tissues (“Vascular Tissues”) that were able to transport water and nutrients throughout the “plant”. The specialization of these tissues gave rise to the vascular tissues of modern terrestrial plants. Through natural selection, because of changing environmental pressures, the stage was set for the exploitation of the terrestrial niches and habitats by these early primitive vascular plants such as Cooksonia sp. and Asteroxylon sp. It is thought that these groups evolved into other primitive vascular plants such as the Pterophytes (“True Ferns”), Sphenophytes “Horsetails”), Psilophytes (“Whisk Ferns”), and Lycophytes (“Club Mosses”). These were Spore-producing plants that did not produce seeds for reproduction. A spore must first germinate into a “Gametophyte”, which produces the sperm and eggs necessary for producing the “Sporophyte” (sexual phase plant) after fertilization. One group, the Phylum Pteridospermophyta (the “Seed Ferns”), did develop the “seed” as a means of reproduction, but they became extinct at the beginning of the Mesozoic because of the changing climate. At the end of the Paleozoic, Selaginella sp . (a member of the Phylum Lycophyta), developed the property of producing both male and female spores on a structure called a “Strobilus”. This resembled a “pinecone” and allowed for the retention of the female spores and the releasing the male spores creating a process similar to Pollination. Also, after the strobilus containing the female spore received a male spore, fertilization occurred in the strobilus (rather than on the ground, separate from the mother plant such as in the other primitive vascular plants). Furthermore, the developing sporophyte was nourished and protected by the mother plant’s strobilus, producing for at least the second time during the evolution of life on Earth, the “SEED”. From this stock group arose the “Seed Plants” called the “Gymnosperms” (“Naked Seed” because no fruit is produced). Gymnosperms flourished during the Mesozoic to the point of taking over many niches and habitats of the Primitive Vascular plants that had given rise to them. One sees a reduction in the number of Primitive Vascular plants during the Mesozoic because of the rapid spread of Gymnosperms. Included in this Gymnosperm group are the Phyla Coniferophyta (“Conifers”), Ginkgophyta (“Ginkgos”), Cycadophyta (“Cycads”), and Gnetophyta (“Gnetophytes”). All of these members produce their seeds in “Cone-like“structures, releasing their seeds to the wind. Because of changing environmental pressures along with intense competition of all extant plant groups of the time, natural selection caused the development of a certain group of Gymnosperms, namely the Gnetophytes, to undergo an important evolutionary change. This change is seen in the fossil evidence in plants related to the Gnetophyte group containing a plant called Gnetum sp. This tropical, vine-like Gymnosperm had a reproductive habit of clustering its strobili at the ends of their branches and protecting them with modified leaves and other plant organs. This was the birth of flowering and fruiting plants: the “Angiosperms” (Phylum Anthophyta). One of the earliest Angiosperm fossils found dates back to the Late Jurassic/Early Cretaceous (approximately 150 million years ago). These fossil flowers resemble the modern Magnolia flower. Since the seeds of Angiosperms are kept to developmental term in the ovary of the flower (the “Fruit”), they were more successful at reproductive fecundity (creating offspring) than the Gymnosperms. So, once again we see a more successful product of natural selection, the flower and fruit, take over the niches and habitats of a previously successful Gymnosperm group. Near the end of the Mesozoic, we see from the fossil evidence a rapid decline in the number and variety of Gymnosperms and the rapid adaptive radiation and proliferation of the Angiosperms throughout the Cenozoic through today. Botanically speaking, the Paleozoic is known as the “Age of Primitive Vascular Plants”, the Mesozoic as the “Age of Gymnosperms (particularly the Cycads)”, and the Cenozoic as the “Age of Angiosperms”. It has even been speculated, and in many cases substantiated by fossil evidence, that the rise of the Angiosperms contributed to the demise of the Dinosaurs. By diminishing the Gymnosperm food supply for the herbivorous dinosaurs, upsetting predator/prey ratios of the populations, and by possibly producing pollen that was foreign to some dinosaur’s immune systems, the Angiosperms are thought to have played a major role in the extinction of the dinosaurs. What possibilities lie ahead for the next successor in the evolution of the Plant Kingdom? Life of the Lower Paleozoic Era Epieric Seas: broad, shallow, warm, nutrient seas that were formed by continental fringe collisions that created "Arches and Basins". These basins were filled with seawater as they formed creating almost an "incubator" effect on marine life forms. As the tilt of the axis of the earth wobbled throughout geologic time causing growth of the ice caps, followed by their melting, these Epieric seas would fill and drain accordingly. Whenever there was times of ice cap buildup, there was a simultaneous "Eustatic" (= "worldwide") sea level drop draining the Epieric seas causing extinctions and intense environmental pressures on the life forms. Whenever the ice caps melted, there was an eustatic rise in sea level resulting in the epieric seas to again fill and the incubator effect would cause life forms to flourish. The subsequent rise and fall of the sea level is recorded in the geologic record as series of changing marine lithofacies upon the continents called Cratonic Sequences. These changes (i.e. the rise and fall of the Sauk Cratonic) were the driving forces of life evolution, molding and establishing ecosystems worldwide. (see Cratonic Sequence Chart) The Cambrian Explosion: the adaptive radiation of the marine life of the Cambrian due to not only the development of the epieric seas, but also on the rise of gene mutations seen in Cambrian marine life that utilized calcium carbonate and calcium phosphate for the first time as body parts: bone, shells, & teeth. Hence, the beginning of the Phanerozoic Eon ("visible life" meaning that there began to be abundant fossils in the geologic record). This increased the competition of the species worldwide resulting in the so-called "Cambrian Explosion of Life". This also is what separates the Edicarian (Late Proterozoic 700MYA - 570 MYA) "soft bodied metazoan" fossil assemblages from the "hard shelled" Cambrian assemblages. Life forms beginning in the Cambrian: 50% of all fossils of the Cambrian are trilobites - Phylum Arthropoda, Class Trilobita. All Periods of the Paleozoic have trilobite Index Fossils. Archeocyathids - little known Coral-like? Sponge like? Organisms that flourished for a while but became extinct in the Early Paleozoic. Brachiopods - Phylum Brachiopoda, Class Articulata & Class Inarticulata became extremely abundant, even throughout the Paleozoic. Most Cambrian life forms fall into the preceding three groups: Trilobites, Brachiopods, and Archeocyathids. Phylum Mollusca - Classes Gastropoda (snails), Bivalvia (clams), Cephalopoda (nautilus, octopus, squid) also were abundant starting in the Cambrian throughout most of the Paleozoic. Early Echinoderms- Phylum Echinodermata Class Crinoidea (et al) were established. Many other bizarre forms of life evolved but became extinct in the Early Paleozoic The Burgess Shale: Discovered in the Canadian Rockies of British Colombia by Charles Doolittle Walcott in 1910 is world renown for excellent fossil evidence of the “Cambrian Explosion”. Neotony: The retention of larval characteristics into the sexually reproductive adult form. This is a major driving force of evolutionary change as in: Echinoderm Bipinnaria Larvae: This is one of the best examples of neotony known. The bilaterally symmetrical echinoderm larvae is the stock group of the Phylum Chordata. By remaining nektonic, some larvae developed a notochord for support and locomotion. This created the Phylum Chordata. By the addition of mutations within this group, namely the absorption of calcium salts from the sea, allowed for the formation of internal bony supports protecting the spinal cord. This created the first member of the Subphylum Vertebrata. First Vertebrate Organism: The Agnathans – These were the first vertebrate fishes of the Cambrian. They in turn became the stock group that gave rise to all subsequent vertebrate groups. Ontogeny Recapitulates Phylogeny – Change within an organism’s lifetime (ontogeny) retells (recapitulates) its evolutionary history (phylogeny). The Rise of the Chordates and Vertebrates: The Cambrian seas were the site of another adaptive radiation of the vertebrate fish organisms. From the “jawless” Agnathan (Ostracoderm) stock, there arose the following: Placoderms – “armored fishes” - Silurian to Mississippian Acanthodii – Silurian to Permian Chondrichthyes – “sharks, skates, and rays” - Devonian to Recent Osteichthyes – “bony fishes” - Devonian to Recent The Ordovician This is marked by a major extinction due to the regression of the Sauk. Bryozoans and Graptolites began to inhabit the seas. But, with the rise of the Tippecanoe Cratonic Sequence, life again began to rebound. Evidence shows that the climate worldwide began to stabilize in the Ordovician. Corals (Rugosa and Tabulata) began to inhabit the warm shallow seas. The Silurian First coral reefs appeared with their lagoons mimicking epieric seas. This production of calm, warm, nutrient-rich lagoons, along with the production of numerous niches and habitats was their major importance. Know the cross-section of a coral reef: land, lagoon, reef core, reef tallus, fore-reef, and back-reef The first land plants (modified Chlorophyte Algae w/ waxy cuticle and vascular tissue) altering the climate and creating a more oxygen rich atmosphere. Ozone levels began to rise as lowland forests spread across the continents. Eumeric flora in Laurasia and Glossopteris sp. seed fern forests in Gondwana. (see evolution of Plants page). The rise of terrestrial plants provided a food supply that was soon exploited. Eurypterus sp. "sea scorpions" were the first animal life to crawl onto land and it is thought that they were exploiting the terrestrial plant diversity as a food supply. This lead to Eurypterus sp. giving rise to terrestrial arthropods (insects, et al.) creating yet another food supply to become exploited. The Taconic Orogeny (convergence of N. America and Africa) was first uplift of the Appalachians, diverting the jet stream and altering the weather patterns resulting in a very long "swampy" climate of most of the equatorial continents. Life in the Upper Paleozoic: The Devonian "Age of Fishes" Agnathan/Ostracoderm Stock Group – These Cambrian jawless fishes became the stock group for the rise of fishes during the Devonian. Placoderms – “armored fishes” - Silurian to Mississippian Acanthodii – Silurian to Permian Chondrichthyes – “sharks, skates, and rays” - Devonian to Recent Osteichthyes – “bony fishes” - Devonian to Recent Rise of Placoderms - the formation of the "Basic Vertebrate Skeleton" – This group genetically established the “tetrapod” (four appendage) body form, possessing an anterior “pectoral girdle” with two appendages, and a posterior pelvic girdle, also possessing two appendages. These Placoderms are named so because of the heavy platy armored coverings found on the surfaces of these fishes. These armors were probably in response to increased environmental pressures and competition between groups. Development of the "Jaw" and Teeth" – Embryonically, the jaws (maxilla and mandible) arise from the calcification and forward rotation of the first two pharangeal branchial arches associated with the gill slits of the Agnathan stock group. In later groups, (chondrichthyes) developed acrodont teeth (set on top of the jaw and held in place with ligaments) from the Dermal Placoid Scales imbedded in its skin. Class Chondrichthyes – the sharks, skates, and rays. These possess cartilaginous skeletons with the only ossified portion being the teeth. These immediately became very successful and diverse, with many forms (i.e. sharks) becoming top carnivore in the sea. The sharks achieved an almost perfect body adaptation to their environment Class Osteichthyes – the “bony” fishes. These possess calcified, internal bony skeletons and diversified greatly in the Devonian and Late Paleozoic. Lobe Fin vs. Ray Fin & Peduncle comparisons – Some groups adapted for fast swimming by developing “ray” fins (thin and maneuverable for speed), and remained in open waters. The peripheral isolates of these coastal groups that could not compete moved into deltaic or swamp environments. These isolate groups developed characteristics or adaptations to anerobic reducing environments: Lung for air breathing Lobe fins covering the pentadactyl appendage Girdle positioning allowing for limbs to be used for support on the bottom These are now called Rhipidistrian Fishes The Pentadactyl Appendage - The 5-fingered vertebrate hand that was chosen for its support superiority: a middle phalange for support, two smaller pair on each side for lateral support. Any more phalanges and they interfere with movement; any less and support is lost. This is an evolutionary archetype common in terrestrial vertebrate organisms. The Labrynthodonts - the First "Amphibian" of the Devonian – were direct descendants the Rhipidistrian stock groups. An “amphibian” means “both life” indicating that these organisms must return to water for reproduction due to their possessing a shell-less egg: the Anamniotic Egg. Modern forms include the frogs, toads, salamanders, newts, etc. Amphibian Characteristics: 3-chambered heart to cope with higher energy requirements having become terrestrial (i.e. coping with gravity) eyelids for prevention of desiccation of the eyes by exposure to the air. Further girdle rotation for limb’s position in a more under body mode. Development of the primal terrestrial ear for hearing in the air. (inner ear bones evolving from pieces of the mandible) Anamniotic shell-less eggs: must be lain in water Example: Semouria sp. The Cotylosaurs - the First "reptile" of the Devonian – beginnings of the formation of Pangea began to dry out some portions of the earth causing reptilian characteristics to develop from the Labrynthodons stock groups. Reptilian Characteristics: 3-chambered heart to cope with higher energy requirements having become terrestrial (i.e. coping with gravity) Even further girdle rotation for limb’s position in an even more “under body mode” for ground clearance when walking on land. Body developed epidermal “scales” for protection from desiccation A better developed ear The development of the amniotic, shelled egg cut the ties with water creating the first truly terrestrial reptilian group – the Cotylosaurs of the Devonian The Mississippian & Pennsylvanian "Age of Coal Forests” During the Mississippian and Pennsylvanian, many of the continental plates were equatorial whereby conditions were swampy. This created the vast coal deposits of today found in the upper eastern states of America. All over the northern continents of Pangea (North America and Eurasia) there were also coal deposits formed during this time from Eumeric Flora (primitive vascular plants). Cyclothems are sequences of coal bearing strata with marine sediments on the bottom, then clay layers (paleo-soils) then the coal seam, then terrestrial layers on top. The presence of these cyclothems in strata core samples tells the modern geologist where coal can be found. In Gondwana, the Glossopteran Flora were “Seed Ferns” of the Phylum Pteridospermophyta, Genus Glossopterus sp. These seed ferns became extinct, but provide evidence that the “seed” has evolved at least twice on the earth. Many coal beds of Gondwana are made of fossil seed ferns. As the continents continued to collide towards the end of the Paleozoic to eventually form Pangea, the coal swamps dried up and the conditions became very arid at the end of the Paleozoic. From the diversification of Cotylosaurs we get Pelycosaurs and Therapsids: The Permian – the forming of Pangea Pangea continues to form, creating arid condition inland dur to a loss of the “coastal effect of weather” since coastlines of individual continents were being fused with other coastlines of other continents. Permian-Triassic “Red beds” were formed from the evaporation of shallow seas and subsequent oxidation of the marine deposits (i.e. Panhandle of Texas – Paladura Canyon) Marine environments were being crushed between the converging continents. Land bridges opened between continents that had been separated by seas. This caused very strong environmental pressures so that there was another Mass Extinction of the Permian. Some estimates place the loss of life from 50% to 80% worldwide. Reptilian characteristics (especially the amniotic, shelled egg) were chosen for by those transitional amphibian groups for survival. The Pelycosaurs – of the Permian - The carnivorous reptilian stock group (i.e. Dimetrodon – sail-back lizards) of the Permian became the stock group of the Mesozoic modern reptiles, marine reptiles, flying reptiles, and Dinosauria. These Pelycosaurs were “thecodonts” having teeth set in sockets. The Therapsids – of the Permian - The “Mammalian Stock Group” of the Permian that gave rise to the first mammal of the Triassic Period. These thecodont reptile-like ancestors of the mammal had a moveable jaw that allowed for “chewing” of foodstuffs. This, along with the abundant Haversian Canals in the bones, in an indication these were “endothermic”, able to create their own body heat. Metabolic Terms associated with body heat: Endothermic - i.e. an organism capable of creating their own body heat (i.e. deriving more energy from its food and having a higher rate of metabolism). Ectothermic – i.e. an organism that must rely upon an external heat source for its metabolism such as basking in the sun. Homeothermic – i.e. an organism that is capable of maintaining a constant body temperature such as birds and mammals. Poikilothermic – i.e. an organism that is not capable of maintaining a constant body temperature such as amphibians and reptiles Evolutionary Relationships of Terrestrial Plants to their Algal Ancestors The fossil evidence for the evolution of modern plants supports the idea that all terrestrial plants evolved from a common ancestor, namely marine algae related to members of the Phylum Chlorophyta (the Green Algae). If one compares the physiology of Chlorophytan cells to modern terrestrial plants, one is able to see many similarities: a cellulose cell wall, storing excess food as starch, similar pigments, etc. The two main differences between Green Algae and Modern Plants are the absence of a “Cuticle” and “Vascular Tissue” in the Green Algae. It is thought that the “Cuticle” (which is a waxy, water-proofing coating found on the outside of plant epidermis) evolved through natural selection from a group of inter-tidal Chlorophytan-type seaweeds that were thrown into intermittent tidal pools and subjected to desiccation. Having the ability to form a protective coat, this group of algae was able to exploit a new habitat in the semi-dry environment of the beachfronts. This stock group was also subjected to natural selection by the competition to survive in this new “frontier”. From this group arose individuals that had series of internal tissues (“Vascular Tissues”) that were able to transport water and nutrients throughout the “plant”. The specialization of these tissues gave rise to the vascular tissues of modern terrestrial plants. Through natural selection, because of changing environmental pressures, the stage was set for the exploitation of the terrestrial niches and habitats by these early primitive vascular plants such as Cooksonia sp. and Asteroxylon sp. It is thought that these groups evolved into other primitive vascular plants such as the Pterophytes (“True Ferns”), Sphenophytes “Horsetails”), Psilophytes (“Whisk Ferns”), and Lycophytes (“Club Mosses”). These were Sporeproducing plants that did not produce seeds for reproduction. A spore must first germinate into a “Gametophyte”, which produces the sperm and eggs necessary for producing the “Sporophyte” (sexual phase plant) after fertilization. One group, the Phylum Pteridospermophyta (the “Seed Ferns”), did develop the “seed” as a means of reproduction, but they became extinct at the beginning of the Mesozoic because of the changing climate. At the end of the Paleozoic, Selaginella sp . (a member of the Phylum Lycophyta), developed the property of producing both male and female spores on a structure called a “Strobilus”. This resembled a “pinecone” and allowed for the retention of the female spores and the releasing the male spores creating a process similar to Pollination. Also, after the strobilus containing the female spore received a male spore, fertilization occurred in the strobilus (rather than on the ground, separate from the mother plant such as in the other primitive vascular plants). Furthermore, the developing sporophyte was nourished and protected by the mother plant’s strobilus, producing for at least the second time during the evolution of life on Earth, the “SEED”. From this stock group arose the “Seed Plants” called the “Gymnosperms” (“Naked Seed” because no fruit is produced). Gymnosperms flourished during the Mesozoic to the point of taking over many niches and habitats of the Primitive Vascular plants that had given rise to them. One sees a reduction in the number of Primitive Vascular plants during the Mesozoic because of the rapid spread of Gymnosperms. Included in this Gymnosperm group are the Phyla Coniferophyta (“Conifers”), Ginkgophyta (“Ginkgos”), Cycadophyta (“Cycads”), and Gnetophyta (“Gnetophytes”). All of these members produce their seeds in “Cone-like“structures, releasing their seeds to the wind. Because of changing environmental pressures along with intense competition of all extant plant groups of the time, natural selection caused the development of a certain group of Gymnosperms, namely the Gnetophytes, to undergo an important evolutionary change. This change is seen in the fossil evidence in plants related to the Gnetophyte group containing a plant called Gnetum sp. This tropical, vine-like Gymnosperm had a reproductive habit of clustering its strobili at the ends of their branches and protecting them with modified leaves and other plant organs. This was the birth of flowering and fruiting plants: the “Angiosperms” (Phylum Anthophyta). One of the earliest Angiosperm fossils found dates back to the Late Jurassic/Early Cretaceous (approximately 150 million years ago). These fossil flowers resemble the modern Magnolia flower. Since the seeds of Angiosperms are kept to developmental term in the ovary of the flower (the “Fruit”), they were more successful at reproductive fecundity (creating offspring) than the Gymnosperms. So, once again we see a more successful product of natural selection, the flower and fruit, take over the niches and habitats of a previously successful Gymnosperm group. Near the end of the Mesozoic, we see from the fossil evidence a rapid decline in the number and variety of Gymnosperms and the rapid adaptive radiation and proliferation of the Angiosperms throughout the Cenozoic through today. Botanically speaking, the Paleozoic is known as the “Age of Primitive Vascular Plants”, the Mesozoic as the “Age of Gymnosperms (particularly the Cycads)”, and the Cenozoic as the “Age of Angiosperms”. It has even been speculated, and in many cases substantiated by fossil evidence, that the rise of the Angiosperms contributed to the demise of the Dinosaurs. By diminishing the Gymnosperm food supply for the herbivorous dinosaurs, upsetting predator/prey ratios of the populations, and by possibly producing pollen that was foreign to some dinosaur’s immune systems, the Angiosperms are thought to have played a major role in the extinction of the dinosaurs. What possibilities lie ahead for the next successor in the evolution of the Plant Kingdom?