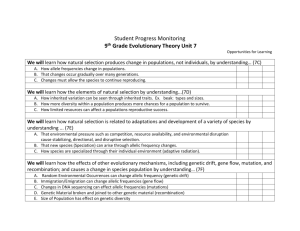
Management implications of intrapopulation and
advertisement

MANAGEMENT IMPLICATIONS OF INTRAPOPULATION AND
INTERPOPULATION GENETIC VARIATION IN NORWAY SPRUCE
Dušan GÖMÖRY & Ladislav PAULE
Technical University in Zvolen, Faculty of Forestry, T. G. Masaryka 24, SK – 960 53 Zvolen, SLOVAK REPUBLIC,
gomory@vsld.tuzvo.sk
Abstract
GÖMÖRY, D., PAULE, L.: Management implications of intrapopulation and interpopulation genetic variation in
Norway spruce.
Spatial genetic structure and kinship relationships were investigated in a montane Norway spruce virgin forest in the
Poana Nature Reserve, Slovakia. A transect of 40 × 150 m was established in the reserve, on which 200 trees were
sampled and analysed for 23 isozyme loci. Twenty–one putatively clonal groups containing 2–6 individuals were
identified on the transect, indicating an important role of the vegetative reproduction. Considerable discrepancies
were found between the observed and expected genotype frequencies at several individuals. Eight independent alleles
were used for the spatial autocorrelation analysis, which showed a weak patchy substructure. The analysis of kinship
relationships proved that practically all individuals belong to half–sib or full–sib families, but there were no clear
relationships between the kinship and the topographical distance of trees. Within a second study, differences among
genetic structures of Norway spruce in 6 primeval forests, 4 naturally regenerated commercial forests and 4 stands
originating from reforestation were assessed. Practically no differences in allelic richness or gene diversity were
found between virgin forests and naturally regenerated stands. However, reforested stands exhibited lower mean
number of alleles as well as lower diversity level as compared to previous two categories. Implications for close to
nature forestry are discussed.
Keywords: spatial genetic structure, genetic differentiation, human impact, Picea abies
Introduction
Virgin forests represent a kind of “standard” for the assessment of human impact on forests, mainly
the consequences of management practices in commercial forests. Nature is frequently considered a
perfectly functioning mechanism where each species and each individual has its defined place and all
processes are at equilibrium. The studies on dynamics of primeval forests have shown that this
assumption is incorrect. A temperate virgin forest passes through three development stages during its
ontogenetical development: the stage of regeneration, optimum stage, and the stage of destruction. The
German terms "das Stadium des Heranwachsens, des Optimums, des Zerfalls" may be more known to
readers familiar with the virgin–forest research (Korpeľ 1995). The stages are dispersed over the area
of the virgin forest in a mosaic–like form and overlap spatially as well as temporally.
Although individual seedlings may survive during the whole development, a massive regeneration
occurs at the beginning of the destruction phase, when trees forming the main canopy start to decline
and more light is transmitted to the soil surface. However, when opening the canopy is not
synchronized with a seed crop, the natural regeneration on mineral soil may be very difficult in
Norway spruce virgin forests due to a strong competition of herbs. Most surviving young spruces with
the height over 1.3 m and thus no more endangered by the competition of herbs can be found on
decaying stems, rocks and other elevated places (Korpeľ 1996). Vegetative propagation by layering of
the lower branches is quite common at high elevations, where the forest is less dense and the crowns
reach the soil.
1
Natural vegetative regeneration is only one of the factors affecting the spatial structure of a natural
forest. The limited pollen and seed flow may contribute to the formation of clumps of half–sibs and
even full–sibs (Brunel & Rodolphe 1985). Microhabitat selection may lead to a non–random
distribution of the genetic variation within a population as well. In case of Central and West European
virgin forest, human activities (introduction of alien genotypes, selective effects through logging etc.)
also cannot be excluded.
Allelic structure in a virgin forest is the result of long–term evolutionary processes. Except
adaptation through natural selection, plant history plays a major role in the formation of the gene–pool
structure. Populations originating from different glacial refugia and/or migratingüto their recent
location by different paths during the Holocene, may differ considerably in the representation of
alleles and genotypes (Taberlet et al. 200x). Founder events during migration or population
bottlenecks may decrease allelic richness and sometimes they create very strange local structures
(Comps et al. 2001). On the other hand, gene flow (mainly through pollen, but in less extent also
through seeds) may wipe out the differences arosen through adaptation or genetic drift. In any case,
gene pool of a primeval population is a dynamic system.
In a commercial forests, most management pracitces are based on negative or (less frequently)
positive phenotype selection. Since the traits used by the forester as the basis for decision are at least
partially genetically controlled, this phenotype selection must be reflected in changed genotype and
sometimes allelic structure of a stand. The criteria of this artificial selection need not necessarily be the
same as those of natural selection.
Natural regeneration in a commercial forest does not allow all trees to reproduce. Variation in
fertility and flowering phenology generally reduces effective population size. Only those trees which
remained after the perparatory cuttings and are flowering and producing seeds at an appropriate time
participate in the formation of young generation. In case of dominant tree species, this is mostly not a
problem. However, in case of rare species, selective harvest or removal of these species as “weeds”
may cause reduction of population size below the level necessary for self–regeneration (Namkoong et
al. 2000).
In case of reforestation, the continuity of a population is interrupted. Local population is replaced
by material with gene and genotype pools formed in completely different environments and under
different circumstances. All the operations starting from the collection of seeds up to planting of
seedlings in the forest represent strong interventions into the genetic pool of a stand.
In this paper we tried to address following questions:
-
To what extent is a virgin forest an equilibrium population with random distribution of genotypes?
-
How natural and artificial regeneration in commercial forests affect genetic structures and levels of
genetic variation?
Materials and Methods
The study consisted of two parts. Within the first one, a transect of 40 × 150 m was established
near the summit of the mountain Zadná Poľana (1,458 m a.s.l.) in a natural Norway spruce forest. A
major part of the transect seems to be at the beginning of the stage of destruction (Zerfallsstadium).
Twigs with dormant winter buds were sampled from all adult individuals and those young spruces on
the transect, whose size allowed to take enough material for the analysis without destroying the tree
(200 samples in total). The position (coordinates x, y) and breast–height–diameter were recorded for
each individual.
Fourteen isozyme systems controlled by 23 loci were used: diaphorase (DIA, EC 1.6.4.3),
fluorescent esterase (FEST, EC 3.1.1.1), glutamate dehydrogenase (GDH, EC 1.4.1.2), glutamate–
2
oxaloacetate transaminase (GOT, EC 2.6.1.1), isocitrate dehydrogenase (IDH, EC 1.1.1.42), leucine
aminopeptidase (LAP, EC 3.4.11.1), malate dehydrogenase (MDH, EC 1.1.1.37), NADH–
dehydrogenase (NDH, EC 1.6.99.3), peroxidase (PER, EC 1.11.1.7), phosphoglucomutase (PGM, EC
2.7.5.1), phosphoglucose isomerase (PGI, EC 5.3.1.9), shikimate dehydrogenase (SKDH, EC
1.1.1.25), superoxide dismutase (SOD, EC 1.15.1.1). For most isozymes, the interpretation of
zymograms followed Muona et al. (1987), Lagecrantz et al. (1988) and Poulsen et al. (1983). In
addition, gels were stained also for 6–phosphogluconate dehydrogenase (EC 1.1.1.44), but the scoring
of the loci controlling this isozyme was not reliable, therefore, 6–PGD phenotypes were only used for
the identification of clonal groups.
Based on diploid genotypes, the genotypic and allelic frequencies were calculated. The deviation of
the observed genotype frequencies at each locus from the Hardy–Weinberg expected ones was tested
using 2–test. The expected multilocus–genotype frequencies under the assumption of free
recombination of all involved loci were calculated for each individual and compared with the observed
frequencies.
To examine the kinship relationships within the investigated set of trees, we used the procedure of
Brunel & Rodolphe (1985). Using a Bayesian approach, they quantify the probability of parental
relationship conditionally to the genotypes of a pair of trees (x, y) as:
P{Ai/(x, y)} = P{(x, y)/Ai}.P(Ai) / (∑i P{(x, y)/Ai}.P(Ai))
where: Ai is the kinship degree (A1 – nonrelated, A2 – half sibs, A3 – full sibs), P(Ai) is the proportion
of kinship types, and P{(x, y)/ Ai} is the probability of observing the pair (x, y) under the hypothesis of
the Ai type of kinship. Kinship was then determined for each pair of trees as the type with the highest
probability. The half–sib and full–sib families were subsequently identified using a complete–linkage
cluster analysis.
The patterns of the spatial distributions of individual alleles were identified using the spatial
autocorrelation analysis. The alleles with a frequency less than 0.05 or more than 0.95 were excluded
from the analysis, in biallelic loci (or when the sum of the two most frequent alleles was more than
0.95), only the most frequent allele was used. Moran's I–statistics (Sokal & Oden 1978) was estimated
for 15 distance classes with approximately equal sample sizes.
Within the second part, genetic structure of 14 Norway spruce populations in Slovakia was studied.
Pairs of geographically proximate primeval forests and commercial forests originating from either
natural regeneration or artificial reforestation were chosen for sampling. Among the analyzed
populations, six stands represented virgin forests, four stands were naturally regenerated commercial
forests and four stands originated from reforestation. In each population, twigs with dormant winter
buds were taken from 30 to 60 non–adjacent trees randomly distributed over the stand. Six isozyme
systems (ACO, GDH, GOT, LAP, PGI, SKDH – for full names and EC codes, see above) controlled
by 8 loci were used. Based on diploid genotypes, allelic frequencies, observed and expected
heterozygosities, and genetic distances (Nei 1978) were calculated. The matrix of genetic distances
was interpreted using principal coordinate analysis.
Results
Spatial genetic structure of a virgin forest
Most loci in the forest of the Polana reserve were monomorphic or exhibiting minor
polymorphisms. Therefore, for the identification of clonal groups also the 6PGD–phenotypes were
used. Despite the fact that we did not score the loci controlling the variation in this isozyme system,
both 6–pgd–B and 6–pgd–C loci seem to exhibit major polymorphisms providing a wide variety of
phenotypes and thus increasing considerably the reliability of clone discrimination.
3
At two loci, an unexpected distribution of alleles in individual genotypes was observed. In the
locus Per–A, rare alleles 97 and 103 occur exclusively in 2 individuals (probably belonging to the
same clone, although separated by a distance of 6 m with another
Table 1. Observed and expected frequencies of selected Norway spruce multilocus genotypes
No.
X
Y
5
14
17
45
49
69
92
12.9
26.1
16.5
9.6
17.6
6.3
26.7
8.2
19.2
6.1
45.7
51.6
66.5
85.7
Genotype frequency
observed
expected
0.005
2.8933E–09
0.005
5.6249E–09
0.005
8.2221E–08
0.005
1.5305E–08
0.005
7.0235E–08
0.005
2.1067E–09
0.005
5.5857E–10
No.
X
Y
107
131
133
143
160
161
170
12.3
18.2
12.8
–4.2
31.0
30.5
12.4
87.4
103.8
103.5
112.3
126.1
127.4
123.4
Genotype frequency
observed
Expected
0.005
1.6092E–10
0.010
7.7993E–15
0.010
7.7993E–15
0.005
5.1889E–08
0.010
1.2831E–12
0.010
1.2831E–12
0.005
5.5114E–08
tree located between them). Exactly the same situation is with the Pgm–A locus: the alleles 104 and 88
occur in two heterozygote trees growing in a clump, but nowhere else.
Within the transect, fourteen clumps containing two or three stems forked at the ground level were
found. All stems were sampled and treated as separate individuals. In three cases, the clumps
contained different genotypes. On the other hand, 21 groups of identical genotypes containing 2 to 6
individuals occurred on the investigated plot. Most of them covered an area corresponding to a crown
or a root system of an adult tree (diameter of the group up to 8 m), but two groups were larger and
seven groups of identical genotypes contained individuals distributed over different parts of the
transect. In 12 cases, compact groups contained individuals of different ages, in one case even
different generations.
At the single–locus level, there are no significant deviations from the Hardy–Weinberg equilibrium
(where tests are possible, considering the sample size). However, the comparison of the observed and
expected multilocus genotype frequencies proved that there is a considerable deviation from the
equilibrium (Table 1). The rare alleles at several loci (Per–A, Pgm–A, Mdh–C) tend to be concentrated
in few individuals rather than to be dispersed at random over the whole examined genotype set.
For the evaluation of kinship following Brunel & Rodolphe (1985), only the adult trees with BHD
18 cm were used, and the putative clonal groups were pooled (together, 132 genotypes were used).
The distribution of individual kinship types among tree pairs is as follows: PNR = 0.48, PHS = 0.32, PFS
= 0.20. Almost all trees belong to some clonal group, half–sib or full–sib family. There seems to be,
however, practically no relationship between the kinship and the distance between individuals (Fig.
1a–b).
4
Fig. 1a. Distribution of the percentage and number
of half–sib pairs over 5 m distance classes
Fig. 1a. Distribution of the percentage and number
of full–sib pairs over 5 m distance classes
Fig. 2a. Numbers of statistically significant Moran’s
indices and average values – clonal groups pooled
Fig. 2b. Numbers of statistically significant Moran’s
indices and average values – all individuals
Spatial autocorrelation analysis was performed for 8 frequent and independent alleles. Since this
analysis was aimed at discovering the patterns arising through the generative regeneration, the
genotypes belonging to the same putative clonal group were pooled and the center was taken as the
position of the group. There is no concordance among the correlograms of individual alleles. Out of
120 values, ten Moran's indices were significant (Fig. 2a). Surprisingly, only one positive Moran's I
was found in the first distance class. Nevertheless, some generalization based on the average I indices
and the occurrence of significant values is possible. The autocorrelation is generally positive in the
first distance class (0–10.8 m). Then, it varies around the expected value, and for distance classes of
31.4–36.6 m, 36.6–42.1 m, and 42.1–48.1 m it becomes negative. Significant positive values occur
again in the distance class of 71.5–81.6 m. If clonal groups are not pooled, the tendencies are similar,
even more pronounced (Fig. 2b). The average Moran's I as well as the bars of the numbers of
significant I–values seem to follow a sinusoid–like curve with the maxima around 0 and 75 meters and
the minimum around 40 m.
5
Effect of stand management on genetic structures
The estimates of average heterozygosities and allelic richness is based on only 8 loci, which are
generally polymorphic, so they are biased upwards. However, they allow comparisons among
individual populations and categories of stands.
Table 2 presents mean values of parameters of allelic richness, gene diversity and heterozygosity
over the investigated 8 loci in individual stands and categories of stands. Although the number of
investigated populations is too small to allow any statistical tests of the differences observed, a general
trend can be identified. The averages of genetic variation parameters in naturally regenerated managed
stands are fully comparable with those of virgin forests, which are considerd to be standards, they are
even higher in most cases. On the contrary, populations of reforested stands contain less alleles and
exhibit lower levels of heterozygosity and gene diversity.
No clear geographic pattern of the genetic differentiation could be identified. On the other hand,
there are considerable differences among categories of stands in the levels of differentiation (Fig. 3).
Virgin forests and naturally regenerated commercial stands are characterized by rather low level of
diferetiation, genetic distances among them are relatively small, not exceeding 0.02. On the other
hand, reforested stands are generally very divergent, but not in a consistent way. The highest value of
genetic distance was observed just between two reforested stands Zádiel and Kremnica Mts. (D =
0.087).
Table 2. Mean number of alleles per locus (na), expected (He) and observed (Ho) heterozygosities in
categories of Norway spruce stands in Slovakia
Category
Population
Bielovodská valley
Chopok reserve
Zadná Poana
Babia hora
Virgin forests
Vtánik reserve
Mláik
Mean
Tatranské Matliare
Chopok commercial forest
Naturally regenerated
Poana commercial forest
commercial forests
Levoa Mts.
Mean
Oravské Beskydy Mts.
Vtánik commercial forest
Stands originating from
Kremnica Mts.
the reforestation
Zádiel
Mean
na
2.87
2.37
2.87
2.87
2.87
3.00
2.81
2.87
2.87
3.00
2.62
2.84
2.62
2.87
2.75
2.37
2.65
He
0.337
0.318
0.353
0.370
0.397
0.337
0.352
0.350
0.422
0.371
0.377
0.380
0.328
0.338
0.311
0.247
0.306
Ho
0.278
0.330
0.350
0.331
0.333
0.310
0.322
0.318
0.376
0.345
0.360
0.350
0.286
0.337
0.253
0.225
0.275
6
axis 2 (18.5 %)
2
1
0
-1
-2
-5
-4
-3
-2
-1
0
1
2
3
4
5
6
axis 1 (55.3 %)
virgin forests
natural regeneration
reforestation
Fig. 3. Principal coordinate analysis of the matrix of Nei’s genetic distances among the investigated
Norway spruce populations.
Discussion
The single–locus tests did not reveal any significant deviation from the Hardy–Weinberg
equilibrium, but there were several trees observed, whose expected multilocus genotype frequencies
are extremely low. We are aware of the fact that the assumption of free recombination does not
completely hold for the set of investigated loci, so that the true expected genotype frequencies could
be slightly higher in several particular cases. However, we do not suppose that the difference between
the true expected frequencies and those calculated under the assumption of free recombination could
be essential and could affect the interpretation of results. Firstly, the investigated population is a
natural forest, surviving at the same site for tens, maybe over hundred generations, which is a
sufficient time for establishing the linkage equilibrium. Secondly, most loci used in this study are
unlinked or loosely linked (Lundkvist 1979, Muona et al. 1987, Poulsen et al. 1983). The most
plausible explanation for such deviation is mating among the relatives. This hypothesis is supported by
comparatively high proportions of half–sib and full–sib pairs of trees, higher than those observed by
Brunel & Rodolphe (1985). The difference between the outhcomes of both studies was probably
caused by life history. Brunel & Rodolphe examined an almost even–aged, semi–natural forest,
regenerated from seed–trees, growing at an altitude of 875 m, i.e. in optimum conditions. It follows
from the description of the forest stand that the regeneration at the time of stand establishment was
distributed regularly over the stand area, and that the spacing among trees was probably regulated by
thinnings. In such conditions, the family structure, probably existing at early stages of the stand
ontogeny due to a limited seed dispersal, is gradually destroyed by stochastic process of reduction of
the number of trees. On the other hand, our object was a transect in a virgin forest, uneven–aged,
growing in adverse climatic conditions of a high–elevated mountain summit. The regeneration is
spatially and temporally irregular, mostly limited to decaying stems, rocks and other elevated places.
Seed crops are small and scarce due to unfavourable climate. Since the spatial distribution of trees is
incomparably more irregular than in a managed forest and there is no human intervention aimed at a
more regular spacing, the chance for a continuous preservation of the family structure is much higher.
In addition, we cannot exclude that in some cases the parent–offspring relationships between some
trees occurred in our material. We tried to limit the other consanguinity relationships than the sibling
ones by excluding the younger trees from the analysis, but we did not determine age for all
individuals. Groups which arose probably though vegetative regeneration, are present in the Poana
7
forest as well. The mating among individuals belonging to the same clone produces offsprings similar
to those produced by mating among full–sibs. This is probably the main cause of an unexpectedly high
PFS–value.
There is a coincidence of the proportions of half–sibs and full–sibs with correlograms. However,
this coincidence might result from the similarity of principles of both methods and identical source
data. The sole existence of compact groups of genetically identical trees (which are very probably
clonal) is a reason for considering the spatial distribution of genes non–random. However, even when
clonal groups are pooled, i.e. when only the patterns arising through generative regeneration are
considered, a spatial structure can be identified, although the patterning is very weak. The sinusoidal
course of the average correlogram indicates a patchy structure, with a patch size of approx. 20–25 m.
A similar patchy structure has been found in other forest trees as well. The results presented by
Knowles (1991) indicate even a similar patch size at least in one of the two investigated stands, when
the generally positive Moran's I in the distance class 0–7.5 m changes to significant negative values
for the class 16.5–30 m. On the other hand, Epperson & Allard (1989) found practically no spatial
pattern of allele distribution in lodgepole pine.
There are several mechanisms of the formation of patches, among them the restricted pollen and
seed dispersal and spatially varying microenvironmental selection seem to be the most important
(Epperson 1992). In the conditions of the Poana virgin forest, where the intraspecific competition is
weak due to a low density of regeneration and due to the fact that most seedlings survive on decaying
stems, selection does not seem to play an important role in the formation of the spatial structure.
Mitton (1983) provides many examples of the existence of family structure in conifers. Although
the Norway spruce pollen may be transported by the wind over tens of kilometers without the loss of
viability, most matings occur between the closest neighbours (Langlet ex Schmidt–Vogt 1978).
Similarly, the reach of seeds (defined as the sedimentation distance of 96% of grains) amounts to 66
m, (Müller ex Schmidt–Vogt 1978), but most seeds fall beneath the tree. Based on these
considerations, a natural Norway spruce stand may be expected to be composed of overlapping,
loosely aggregated families. We can thus conclude that the investigated spruce virgin forest does not
seem to exhibit the traits of a panmictic equilibrium population. Nevertheless, this does not mean that
it cannot be used as a standard for the evaluation of the effects of human activities.
Within the second study, primeval forests were just used as standards for monitoring the effects of
regeneration procedures in commercial forests on the levels and patterns of genetic variation. We have
not found any pronounced difference in genetic variation between virgin forests and managed stands
originating from natural regeneration. As indicated by genetic distances, there are very small
differences in allelic structures between pairs of populations growing in the same region. The whole
group of naturally regenerated forests (virgin and managed ones) is quite compact, as expected, since
the whole investigated territory is small as compared to the distribution range of spruce. All
parameters of genetic variation, i.e. allelic richness, gene diversity and heterozygosity, are even higher
in managed forests compared to primeval ones, although the difference is small and if the number of
sampled populations allowed testing, it would probably be nonsignificant. This confirms previous
results obtained in Douglas–fir (Neale 1985) and Scots pine (Yazdani et al. 1985). High outcrossing
rate and a large effective population size of the parental population can explain high levels of genetic
variation in case of most silvicultural procedures based on natural regeneration.
On the other hand, the genetic structure of artificially established stands is more altered. The levels
of genetic variation are lower as compared to naturally regenerated stands, the populations are
depleted of a part of alleles, and in two of four investigated populations a substantial change of allelic
frequencies occured, as reflected by high genetic distances from the other populations as well as from
each other. The restricted number of maternal trees and resulting genetic drift may be the cause. The
minimum number of maternal trees has not been prescribed neither in Slovakia nor elsewhere in
8
Europe for a long time. Theoretically, sufficient genetic variation should be ensured through pollen
dispersal even when cones are collected from few trees, but apparently pollen dispersal is not as
perfect as it appears on the first sight.
What can be concluded from the second study is that reforestation may (but not necessarily needs
to) result in forest stands with substantially altered genetic structures and decreased genetic variation.
Again, they have not necessarily to be less adapted or less adaptable than stands originating from
natural regeneration, but the risk seems intuitively to be higher. In any case, natural regeneration
apparently allows to avoid this risk at all.
References
Brunel, D., Rodolphe, F., 1985: Genetic neighbourhood structure in a population of Picea abies L.
Theoretical and Applied Genetics 71:101–110.
Comps, B., Gömöry, D., Letouzey, J., Thiébaut, B., Petit, R.J., 2001: Diverging trends between
heterozygosity and allelic richness during postglacial colonization in the European beech. Genetics
157(1): 389–397.
Epperson, B.K., 1992: Spatial structure of genetic variation within populations of forest trees. New
Forests 6:257–278.
Epperson, B.K., Allard, R.W., 1989: Spatial autocorrelation analysis of the distribution of genotypes
within populations of lodgepole pine. Genetics 121:369–377.
Knowles, P., 1991: Spatial genetic structure within two natural stands of black spruce (Picea mariana
(Mill.) B.S.P.). Silvae Genetica 40(1):13–19.
Korpeľ, Š., 1995: Die Urwälder der Westkarpaten. Gustav Fischer Verlag, Stuttgart – Jena – New
York, 310 pp.
Korpeľ, Š., 1996: Dynamika a formovanie prírodných lesov v biosférickej rezervácii Poana.
Research report, Technical University in Zvolen, Zvolen, 57 pp.
Lagercrantz, U., Ryman, N., Ståhl, G., 1988: Protein loci in diploid tissue of Norway spruce (Picea
abies K.): description and interpretation of electrophoretic variability patterns. Hereditas 108:149–
158.
Lundkvist, K., 1979: Allozyme frequency distribution in four Swedish populations of Norway spruce
(Picea abies K.). I. Estimation of genetic variation within and among populations, genetic system
and mating system parameters. Hereditas 108:149–158.
Mitton, J.B., 1983: Conifers. In: S.D. Tanksley, T.J. Orton (eds.), Isozymes in Plant Genetics and
Breeding, Part B. Elsevier Science Publishers, Amsterdam, 443–472.
Muona, O., Yazdani, R., Lindqvist, G., 1987: Analysis of linkage in Picea abies. Hereditas 106:31–
36.
Namkoong, G., Koshy, M.P., Aitken, S., 2000: Selection. In: Forest Conservation Genetics. Principles
and Practice. A. Young, D. Boshier and T. Boyle (eds.), CSIRO Publishing, Collingwood,
Australia and CABI Publishing, Oxon, United Kingdom, p. 101–115.
Neale, D.B., 1985: Genetic implications of shelterwood regeneration of Douglas–fir in Southwest
Oregon. Forest Science 31: 995–1005.
Nei, M., 1978: Estimation of average heterozygosity and genetic distance from a small number of
individuals. Genetics 89: 583–590.
9
Poulsen, H.D., Simonsen, V., Wellendorf, H., 1983: The inheritance of six isozymes in Norway spruce
(Picea abies (L.) Karst.). Forest Tree Improvement 16:12–33.
Schmidt–Vogt, H., 1978: Genetics of Picea abies (L.) Karst. Annales Forestales 7(5):147–186.
Sokal, R.R., Oden, N.L., 1978: Spatial autocorrelation in biology. 1. Methodology. Biological Journal
of the Linnean Society 10:199–228.
Taberlet, P., Fumagalli, L., Wust–Saucy, A.–G., Cosson, J.–F. 1998: Comparative phylogeography
and postglacial colonization routes in Europe. Molecular Ecology 7(4): 453–464.
Yazdani, R., Muona, O., Rudin, D., Szmidt, A.E., 1985: Genetic structure of a Pinus sylvestris L.
seed–tree stand and naturally regenerated understory. Forest Science 31: 430–436.
10