1. Analysis of volatiles from Origanum vulgare

Electronic Supplementary Material
Plant defenses against ants provide a pathway to social parasitism in butterflies
Dario Patricelli, Francesca Barbero, Andrea Occhipinti, Cinzia M. Bertea, Simona Bonelli,
Luca P. Casacci, Simon A. Zebelo, Christoph Crocoll, Jonathan Gershenzon, Massimo E.
Maffei, Jeremy A. Thomas* and Emilio Balletto
* E-mail: jeremy.thomas@zoo.ox.ac.uk
Supplementary Material and Methods.
1.
Analysis of volatiles from Origanum vulgare
2.
Analysis of phytohormones by liquid chromatography-tandem mass spectrometry
(LC-MS) of control and Myrmica -infested Origanum vulgare.
3.
Total RNA extraction from control and Myrmica -infested Origanum vulgare leaves, cDNA synthesis and qPCR conditions and analysis.
4.
Total RNA extraction from different ant species, cDNA synthesis and qPCR conditions and analysis.
5.
The rationale for choosing candidate genes involved in the putative perception and detoxification of carvacrol by ants.
Supplementary references
Tables S1-S4
Figure S1
1
1. Analysis of volatiles from Origanum vulgare
O. vulgare leaves were collected in the experimental site Parco Fluviale Gesso e Stura, Cuneo,
North Italy. Samples were immersed in liquid nitrogen and stored at -80°C until used. The extraction was performed with a 2:1 mixture of pentane/diethyl ether (Carlo-Erba, Italy) in a ratio 1:10 p/v, three times. Pulegone (TCI-Europe) was used as internal standard. Qualitative and quantitative analyses were performed by GC-MS (6890N-5973A, Agilent Technologies,
USA) and GC-FID (6890 Agilent Technologies, USA), respectively. VOCs were separated on a Zebron ZB5-MS column (30 m length, 250 µm diameter, 0.25 µm thickness of stationary phase) (Phenomenex, USA). The injector was set up in splitless mode at 250°C. Helium was used as carrier gas at constant flow (1 mL min
-1
). The thermal gradient was: 60°C for 5 min, from 60°C to 250°C at 3°C min -1
, and 250°C for 5 min. For GC-MS analyses, the transfer line to MSD was at 280°C, ionization energy (EI) was 70 eV and the acquisition scan was from 50 to 250 m/z. The separated compounds were identified by comparisons with pure standards, by comparisons of their mass spectra and Kováts retention indexes with those of reference substances, and by comparison with the NIST mass spectral search software v2.0 using the
NIST 98 and Adams’ libraries [S1].
2. Analysis of phytohormones by liquid chromatography-tandem mass spectrometry (LC-
MS) of control and Myrmica-infested Origanum vulgare
Leaves were collected from control and Myrmica scabrinodis infested O. vulgare plants, placed immediately in liquid nitrogen and kept at -80 °C until extraction. Leaf samples (200-
300 mg) were finely ground and 1 ml of methanol (Carlo-Erba, Italy) was added.
Dihydrojasmonic acid (200 ng) (TCI-Europe, Belgium) was added as internal standard.
2
Samples were kept on ice and extracted for 30 min and mixed by vortex every 5 minutes. Plant material was removed by centrifugation for 5 min at 9000 g at room temperature. Sample clean-up was performed by C18 SPE (SampliQ C18 Endcapped, 500mg, Agilent Technologies,
U.S.A.). Phytohormones were eluted with 500 μl of methanol (Carlo-Erba, Italy).
Phytohormones were analysed by liquid chromatography (1200 HPLC, Agilent Technologies,
U.S.A.) equipped with a reversed-phase column Luna C18 (3.00 μm, 150 x 3.0 mm,
Phenomenex, U.S.A.). The binary solvent system was: (A) acidified double distilled water
(Millipore, U.S.A.) with 0.1% v/v formic acid (Sigma-Aldrich, USA), and (B) acetonitrile
(Carlo-Erba, Italy) with 0.1% v/v of formic acid. Separation was performed at 0.2 ml min
-1 flow rate, 25°C as column temperature and the following solvent gradient: 40% of B for 1 min,
55% B at 4.5 min, 62% B at 5 min, 65% B at 14 min, 98% B at 16 min and 40% of B at 18 min. The initial mobile phase was re-established for 8 min before the next injection. Mass spectrometry analyses were performed with a 6330 Series Ion Trap LC-MS/MS System
(Agilent Technologies, USA) equipped with an electrospray ionization source (ESI).
Quantitative analyses were made in negative mode with 1.5 kV ion spray voltage, nebulizer gas
(N
2
) at 10 L min
-1 , 325 °C and 1.00 V fragmentation amplitude The monitored mass transitions in MRM mode were 209→59, 211→59, 322→130 and 291→273 m/z for Jasmonic acid (JA),
Dihydrojasmonic acid (IS), (3R,7S)-Jasmonoyl-L-isoleucine (JA-Ile) and cis-(+)-12oxophytodienoic acid (OPDA), respectively. Spectral data were processed and analyzed by
DataAnalysis for 6330 Series Ion Trap LC/MS 4.0 software (Bruker Daltonik, Germany).
External calibration curves were made with standard solutions of Jasmonic acid (Sigma-
Aldrich, USA), JA-Ile and OPDA (Cayman, USA).
3
3. Total RNA extraction from control and Myrmica-infested Origanum vulgare leaves, cDNA synthesis and qPCR conditions and analysis
Total RNA was extracted from 50 mg of ant-induced and control O. vulgare leaves ( n = 6) using the Agilent Plant RNA Isolation Mini Kit (Agilent Technologies, USA). To remove residual genomic DNA, RNA was treated with RNAse-free DNAse I set (Qiagen, Germany).
RNA quality was checked using the Agilent 2100 Bioanalyzer with RNA 6000 Nano LabChip.
Quantitative analysis was performed using the Nano Drop ND-1000 micro-scale spectrophotometer (Thermo Fisher Scientific, USA).
For cDNA synthesis, High-capacity cDNA Reverse Transcription Kit (Applied Biosystems,
USA) was used according to manufacturer’s instructions. Reactions were prepared by adding
1.5 µg total RNA, 2 µl of 10X RT Buffer, 0.8 µl of 25X dNTPs mix (100 mM), 2 µl 10X RT random primer, 1 µl of Multiscribe TM
Reverse Transcriptase and nuclease-free sterile water up to 20 µl. Reaction mixtures were incubated at 25°C for 10 min, 37°C for 2 h, and 85°C for 5 sec. Samples were stored at -20°C for further analyses. Primer pairs for cytochrome P450s
CYP71D178-182 were designed using BeaconDesigner (version 5.0; PremierBiosoft, Palo
Alto). Primers were designed for regions with the largest possible difference between the P450 sequences whilst primer binding sites were chosen for identical regions of all known alleles for the respective genes from O. vulgare and Thymus vulgaris . Each primer pair was tested for potential cross-hybridisation with the other P450s as templates; none was observed at a 60 °C annealing temperature except that the CYP71D179 primers cross-reacted to some extent with the CYP79D178 and D182 sequences due to their close sequence similarities. Primers are listed in table S4. Primer efficiencies for all pairs were calculated using the standard curve method [S2, S3].
4
qPCR analysis was on a Stratagene Mx3000P Real-Time System (La Jolla, CA, USA) using
SYBR green I with ROX as internal loading standard, using 25 μl of mixture consisting of 12.5
µl 2X MaximaTM SYBR Green qPCR Master Mix (Fermentas), 0.5 µl cDNA and 100 nM primers (Integrated DNA Technologies). Controls included non-RT controls (using total RNA without reverse transcription to monitor genomic DNA contamination) and non-template controls (water template). PCR conditions were: 10 min at 95°C, 40 cycles of 15 s at 95°C, 20 sec at 60°C, 30 sec at 72°C. Fluorescence was read after each annealing and extension phase.
All runs were followed by a melting curve analysis from 55°C to 95°C. The linear range of template concentration to threshold cycle value (Ct value) was determined by performing a dilution series using cDNA from three independent RNA extractions analyzed in three technical replicates. qPCR reactions were run using specific primers [S4]. Three distinct reference genes: elongation factor 1 alpha [S2], actin and 18S rRNA [S5] were used to normalize the results of the real-time PCR. The most stable gene, selected using Normfinder software [S6], was the elongation factor 1 alpha. All amplification plots were analysed with
MX3000P
TM
software to obtain Ct values. Relative RNA levels were calibrated and normalized with the level of the elongation factor 1 alpha mRNA.
4. Total RNA extraction from different ant species, cDNA synthesis and qPCR conditions and analysis
Frozen ants’ total RNA was isolated using Qiagen RNeasy Micro kit and RNase-Free DNase set to remove residual genomic DNA. Sample quality and quantity was checked by using the
RNA 6000 Nano kit and Agilent 2100 Bioanalyzer, and confirmed spectrophotometrically using a NanoDrop ND-1000. High-capacity cDNA Reverse Transcription Kit was used for cDNA synthesis. Reactions were prepared by adding 500 ng total RNA and following the same protocol as in §3 (above). All qPCR experiments were performed on a Stratagene Mx3000P
5
Real-Time System using SYBR green I with ROX as an internal loading standard. The reactions mixtures were prepared as described in §3 (above). PCR conditions were: tubulin
(TUB), Acetylcholinesterase (AChE), actin (ACT)
: 10 min at 95°C, 40 cycles of 15 s at 95°C,
20 sec at 55°C, and 45 sec at 72°C; odorant receptor 1 (
OR1), cytochrome P450 9E2 (CYP450
9E2), Glutathione S transferase (GST), odorant binding protein 6 (OBP6)
: 10 min at 95°C, 40 cycles of 15 s at 95°C, 30 sec at 57°C, and 30 sec at 72°C. Procedures for fluorescence reading, melting curve analysis and determination of the linear range of template concentration to Ct value were as in §3 (above). qPCR reactions were run using specific primers designed on Genbank-available ant sequences using the Primer3 software [S7] (table S1), with efficiencies for all pairs calculated as above [30]. Two reference genes, actin and tubulin, were used to normalize results of the real time PCR, from which actin was selected as in §3 (above) [S6]. Ct values of amplification plots and the calibration and normalization of relative RNA levels (using actin) were as in §3
(above).
5. The rationale for choosing candidate genes involved in the putative perception and detoxification of carvacrol by ants.
The selection of genes analysed in this study was based on their known or suspected function in odour perception and on sequences available from the different ant genomes [S8-S12]. We analysed the expression levels of AChE, GST and a CYP9 for their suspected involvement in the detoxification of carvacrol in different ant species, along with OBP6 and OR1 as potential candidates in the perception of this compound.
The detoxification-related genes we used were previously known from other insects and shown to be involved in the detoxification of different xenobiotics. In particular, the functions
6
of acetylcholinesterases (of insect and mammalian origin) are discussed by Casida and Durkin
[S8]. Inhibitor specificity between the AChE of insects and between susceptible and tolerant species of insects contributes to selective toxicity and resistance. Since insect acetylcholinesterase is often targeted by insecticides and by compounds showing insecticidal properties (such as terpenoids), we deemed it advisable to analyse the expression level of this gene in the different ant species following treatment with carvacrol. But whereas the effect of this compound has been studied on AChE enzyme activities [S9], to the best of our knowledge no study on AchE expression had been carried out following exposure to this compound.
Glutathione S-transferases (GSTs) in insects are a family of enzymes that play a critical role in insecticide detoxification. Since carvacrol and other terpenoids have insecticidal properties, we suspected that they may be able to modulate GST gene expression.
Cytochrome P450s are a class of heme proteins that may also be involved in the oxidation of exogenous substances, such as environmental contaminants. In insects, cytochrome P450s are important detoxification enzymes of organophosphate, carbamate insecticides and other types, including terpenoids with insecticidal properties. Several cytochrome P450s have been reported in insect antennae and may play a role in odorant processing and/or xenobiotic metabolism [S10]. It has recently been reported that CYP4, CYP6 and CYP9 in Bombyx mori participate in the metabolism of plant toxins and insecticides [S11].
Proteins coded by several different gene families contribute to the reception of chemical messages. Growing evidence suggests that odorant binding proteins (OBP) are involved in solubilizing and transporting odorants and pheromones through aqueous haemolymph in insects. The chemical messages carried by the OBPs are decoded when odorant receptors (OR) selectively bind different chemicals [S12].
7
Supplementary References
S1. Adams RP. 2007 Identification of essential oil components by gas chromatography/mass spectrometry . Illinois, Allured Publishing.
S2. Crocoll C, Asbach J, Novak J, Gershenzon J, Degenhardt J . 2010 Terpene synthases of oregano ( Origanum vulgare L.) and their roles in the pathway and regulation of terpene biosynthesis. Plant. Mol. Biol.
73 , 587-603. (doi: 10.1007/s11103-010-9636-1)
S3. Pfaffl MW. 2001 A new mathematical model for relative quantification in real-time RT-
PCR. Nucleic Acids Res.
29 , 2002-2007. (doi: 10.1093/nar/29.9.e45)
S4. Crocoll C (2010) Biosynthesis of the phenolic monoterpenes, thymol and carvacrol, by terpene synthases and cytochrome P450s in oregano and thyme . Ph.D dissertation,
Friedrich-Schiller-Universität. Max-Planck-Institut für chemische Ökologie.
S5. Zebelo SA, Bertea CM, Bossi S, Occhipinti A, Gnavi G, Maffei ME. 2011 Chrysolina herbacea modulates terpenoid biosynthesis of Mentha aquatica . PLoS One 6 , e17195.
(doi: 10.1371/journal.pone.0017195)
S6. Andersen CL, Jensen JL, Ørntoft TF. 2004 Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res.
64
5245-5250. (doi: 10.1158/0008-5472.CAN-04-0496)
S7. Rozen S, Skaletsky H. 2000 Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics Methods and Protocols (eds S Krawetz, SA Misener), pp
365–386. Totowa: Humana Press.
S8. Casida JE, Durkin KA. 2013 Neuroactive insecticides: Targets, selectivity, resistance, and secondary effects. Annu. Rev. Entomol.
58 , 99-117. (doi: 10.1146/annurev-ento-120811-
153645)
8
S9. Jukic M, Politeo O, Maksimovic M, Milos M, Milos M. 2007 In Vitro Acetylcholinesterase
Inhibitory Properties of Thymol, Carvacrol and their Derivatives Thymoquinone and
Thymohydroquinone. Phytotherapy Res.
21 , 259-261. (doi: 10.1002/ptr.2063)
S10. Pottier MA, Bozzolan F, Chertemps T, Jacquin-Joly E, Lalouette L, Siaussat D,
Maïbèche-Coisne, M. 2012 Cytochrome P450s and cytochrome P450 reductase in the olfactory organ of the cotton leafworm Spodoptera littoralis. Insect Mol. Biol.
21, 568-
580. (doi: 10.1111/j.1365-2583.2012.01160.x)
S11. Gu ZY, Li FC, Wang BB, Xu KZ, Ni M, Zhang H, Shen WD, Li, B. 2015 Differentially expressed genes in the fat body of Bombyx mori in response to phoxim insecticide.
Pesticide Biochemistry and Physiology 117 , 47-53.
( doi:10.1016/j.pestbp.2014.10.007)
S12. McKenzie SK, Oxley PR, Kronauer DJC.
2014 Comparative genomics and transcriptomics in ants provide new insights into the evolution and function of odorant binding and chemosensory proteins. BMC Genomics 15 , 718.
(doi:10.1186/1471-2164-15-
718)
9
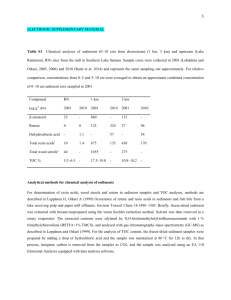
Table S1.
Comparison of volatiles produced by Origanum vulgare plants after experiencing root disturbance by Myrmica sabuleti and My. scabrinodis in experimental terraria. Mean values between treatments ( n = 20, 1df) were compared by ANOVA; data are expressed as μg g -1 fresh weight ± s.e.m..
Compound
p-Cymene
γ-Terpinene
trans-Sabinene hydrate
Methylthymyl ether
Thymol
Carvacrol
α-Cubebene
Kovats index
Plants with
M. sabuleti
Plants with
M. scabrinodis
Statistical test
1024 0.877±0.272 0.947±0.311 F=0.862
1055 0.930±0.261 0.919±0.263 F=0.167
1098 1.086±0.366 0.946±0.310 F=0.972
1173 0.919±0.288 1.210±0.458 F=1.725
1290 1.369±0.452 1.210±0.434 F=0.312
1299 1.312±0.429 1.309±0.401 F=0.001
1351 0.854±0.262 0.880±0.283 F=0.301
α-Copaene
β-Ylangene
γ-Elemene
α-Bergamotene
γ-Muurolene
1413 0.818±0.279 0.844±0.282 F=0.222
1480 2.214±0.575 1.706±0.447 F=1.688
Bicyclosesquiphellandrene 1484 1.248±0.349 1.317±0.365 F=0.181
Germacrene-D 1485 6.503±1.721 4.941±1.484 F=0.653
α-Amorphene
1377 1.058±0.310 1.022±0.312 F=0.131
1421 4.777±1.219 4.221±1.240 F=0.158
1437 1.246±0.366 1.408±0.425 F=0.536
(E)-β-Farnesene
Bicyclogermacrene
α-Bisabolene
δ-Cadinene
α-Humulene epoxide
1485 1.114±0.395 0.989±0.293 F=0.597
1457 3.977±1.139 5.299±1.276 F=0.931
1500 1.068±0.322 1.045±0.287 F=0.063
1507 0.864±0.322 0.791±0.254 F=0.814
1523 0.886±0.281 0.995±0.310 F=1.919
1608 1.730±0.564 1.841±0.524 F=0.067
0.429
0.450
0.347
0.805
0.379
0.183
0.799
0.590
0.722
0.695
0.473
0.643
0.210
0.676
p-value
0.366
0.687
0.337
0.206
0.583
0.990
10
Table S2.
The abundance of major compounds found in leaf extracts of Origanum vulgare growing naturally in the field with or without a Myrmica ant colony in the surrounding soil.
Mean values between treatment and control plants ( n = 10) were compared by ANOVA; significant increases were reported. tr = traces detected, nc = not computable. Data are expressed as μg g
-1
fresh weight ± s.e.m.. * Significanc pvalues controlled for False Discovery
Rate (FDR) by using the Benjamini-Hochberg procedure
Compounds
α-Thujene
α-Pinene
1-Octen-3-ol
Myrcene
α-Terpinene
β-Trans-ocimene
γ-Terpinene
Allo-ocimene
Thymol
Carvacrol
Bicycloelemene
β-Caryophyllene
β-Copaene
Germacrene D
Bicyclogermacrene
α-Farnesene
Germacrene D-4-ol
Kovats index
912
924
976
989
1016
1035
1055
1120
1312
1319
1348
1417
1430
1484
1500
1505
1574
Plants with
Myrmica
1.91 ± 0.07
0.79 ± 0.04
11.55 ± 1.97
7.31 ± 0.76
7.66 ± 0.62
30.53 ± 5.75
84.60 ± 4.00
28.22 ± 0.59
17.16 ± 0.40
Control plants without
Myrmica
0.94 ± 0.48
Tr
8.09 ± 0.33
6.74 ± 0.32
Tr
41.24 ± 0.52
13.61 ± 0.80
50.43 ± 12.81
1.83 ± 0.61
72.89 ± 1.09
20.61 ± 3.40
37.09 ± 1.87
25.56 ± 2.95
46.93 ± 13.34 132.45 ± 24.29
16.53 ± 1.19 21.58 ± 1.96
35.41 ± 1.04 38.17 ± 1.94
31.21 ± 1.12
10.75 ± 3.34
10.64 ± 3.32
34.70 ± 1.57
57.92 ± 20.28
23.16 ± 4.30
Statistical test
F=4.034
Nc
F=3.008
F=0.484
Nc
F=3.439
F=302.332
F=2.997
F=437.157
F=272.937
F=1.212
F=9.522
F=4.856
F=1.578
F=3.283
F=5.266
F=5.317
p-value
<0.0001
0.303
0.015
0.059
0.244
0.108
0.051
0.050
0.079
Nc
0.121
0.506
Nc
0.101
<0.0001*
0.122
<0.0001*
11
Table S3 .
Expression of Origanum vulgare terpene biosynthesis related genes from plants treated with Myrmica scabrinodis ants compared with untreated control plants. Logtransformed data are expressed as mean values of at least three biological replicates (± s.e.m.).
Three technical replicates were run for each biological replicate. ANOVA was used to determine significant differences between treatments and controls (n=6). Gene descriptions not given in figure 2: GPS , geranyl diphosphate synthase; OvTPS6 , a sesquiterpene synthase producing ( E )-β-caryophyllene and α-humulene.
Gene
OvTPS6
GPS
Plants with
Myrmica
Control plants without
Myrmica
0.185 ± 0.030 0.200 ± 0.027
0.739 ± 0.258
0.893 ± 0.102
Statistical test
F=0.127
F=0.307 p-value
0.740
0.609
12
Table S4.
Primers used in this work
Gene
OvEF1alpha
18S rRNA
Act
OvTPS2
OvTPS6
GPS
CYP71D178
CYP71D178/179/182
CYP71D180
Gene
ACT
TUB
OR1
OBP
AChE
GST
CYP4509E2
Origanum vulgare PRIMERS
Forward primer (5’→3’)
CTCCAGTTCTTGATTGCCACAC
ATGATAACTCGACGGATCGC
GCTCCAAGGGCTGTGTTCC
GTGGCTGAGTTTGGTGGAAGG
AGGGCGTATCGGAGGAAGAAG
ATGATAAGCGGGCTGCATAG
CAAGGAATGACTGCTGCTGAC
Reverse primer (5’→3’)
GCTCCTTTCCAGACCTCCTATC
CTTGGATGTGGTAGCCGTTT
TCTTTCTGTCCCATGCCAAC
TTGGCGTTCTCTAGGTATTCTGC
GGTCGTAAGGATTGGAACAGAGG
CCGAAATTCCTCAGCTTCTG
TTGGATTGTGGATTGTTGGAACC
CGCTCTTCTTCACCCTATGC
GATTGAACGTGTCGGGATCT
CGTGGCTTCTCAACCTTCTC
GCAAAGAAGAATGCGAGGTC
ANT PRIMERS
Forward primer (5’→3’)
CTACGGCTGAGCGAGAAATC
CGGCTATTCAGGAGCTGTTC
AAGCCAGCTGGAAAAAGGAT
GAGTCGCCACAGCTGACATA
ACACCGCCGTCAATAAAGAG
GTGGTTACGCAGTCGGTTCT
GGAGCCTACCTTCCTTTTGG
Reverse primer (5’→3’)
AAGGAAGGCTGGAAAAGAGC
CTGCTGATACTCGGACACGA
CTGCTTGGAAGCCAGTTAGG
TTTCTCAACCTCGGCTCTGT
GCTGCGGCAGATACTTTTTC
ATGAGGTCGTCGTGATAGCC
ATCGGCATCGATGTCTTTTC
13
Figure S1.
Establishing the terraria used for experiments
14