Chymotrypsin
advertisement

Biochemistry 462a – Enzyme Mechanisms
Reading - Chapter 16
Practice problems - Chapter 16 - 1,2,4,5; Enzymes extra problems
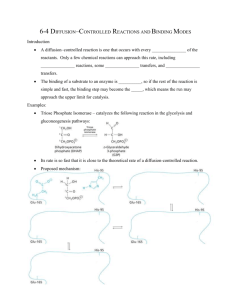
Catalytic Mechanisms
General acid-base catalysis is a
commonly employed mechanism
in enzyme reactions, e.g.,
hydrolysis of ester/ peptide
bonds, phosphate group reactions,
addition to carbonyl groups, etc.
General acid catalysis involves
donation of a proton by the
catalyst.
General base catalysis involves
abstraction of a proton by the
catalyst.
The side chains of Asp, Glu, His,
Cys, Tyr and Lys can be involved
in general acid-base catalysis.
Covalent catalysis involves rate enhancement by the transient formation of a catalystsubstrate covalent bond.
o Covalent catalysis is a commonly employed mechanism in enzyme reactions. The
side chains of His, Cys, Asp, Lys and Ser can participate in covalent catalysis.
o The coenzymes pyridoxal phosphate and thiamine pyrophosphate function mainly
by covalent catalysis.
Metal ion catalysis is widely used.
o Binding substrates in the proper orientation.
o Mediating oxidation-reduction reactions.
o Electrostatically stabilizing or shielding negative charges.
o Metalloenzymes contain tightly bound metal ions: Fe+2, Fe+3, Cu+2, Zn+2, Mn+2.
o Metal-activated enzymes contain loosely bound metal ions: Na+, K+, Mg+2, Ca+2.
Electrostatic catalysis refers to the fact that when a substrate binds to an enzyme, water is
usually excluded from the active site.
o This causes the local dielectric constant to be lower,
o Which enhances charge-charge interactions in the active site.
Proximity and orientation effects are important in enzymatic reactions.
o The three dimensional structure of the enzyme can bring several reactive side chains
into close proximity in the active site.
o Binding of the substrate in the active site can orient the substrate for most efficient
interaction with these side chains.
1
Preferential Transition State Binding is probably the most important rate enhancing
mechanism available to enzymes.
o This means that the enzyme binds the transition state of the reaction more tightly than
either the substrate or product, therefore Go‡ is lowered.
o Weak interactions between the enzyme and substrate are optimized in the transition
state.
Preferential Transition State Binding - how does it work?
Consider this mechanical analogy,
the enzyme stickase.
If the active site of the "enzyme" is
complementary to the substrate, then
Go‡ is raised not lowered and the
“enzyme” does not enhance the
reaction.
If the active site is complementary
to the transition state, then Go‡ is
lowered and the enzyme enhances
the reaction.
G for forming the transition state is partially "paid for" by the energetics of weak
interactions between the enzyme and the transition state.
How much binding energy is needed to account for the large
rate accelerations brought about by enzymes? For a 106-fold
increase in rateGo = 34.3 kJ/mol. This amounts to the
formation of two additional hydrogen bonds between the
enzyme and the transition state, as compared to the interaction
between the enzyme and the substrate.
2
Transition state stabilization by enzymes
The best inhibitor is the planar
transition state analog - pyrrole-2carboxylate, whereas the tetrahedral
substrate analog is a poor inhibitor.
Antibodies against a
transition
state
analog
are
enzymatically active;
antibodies against the
substrate have no
enzymatic activity.
These antibodies have
a site that binds and
stabilizes the transition
state, therefore they
catalyze the reaction.
The Binding Energies of Enzymes and Substrates
The net binding energy represents the differences between the binding energies of the
substrate with water and the substrate with the enzyme. Thus, the dissociation constant of an
enzyme-substrate complex reflects the relative stabilities of the substrate bound to the
enzyme and when it is free in solution. The constant depends on
o The strength of the hydrogen bonding between the enzyme and the substrate,
compared with the combined strengths of their separate hydrogen bonding to water
o The stability of salt linkages between the enzyme and the substrate, compared with
the tendency of individual ions to be solvated by water.
o The presence of hydrophobic interactions.
Although each difference might be small, when summed over all the interactions, the energy
can be considerable. Note that hydrogen bonding and salt linkages between the enzyme and
substrate are entropically favored due to release of water.
3
Utilization of enzyme-substrate binding energy in catalysis
The maximum binding energy between an enzyme and a substrate will occur when there is
maximal complementarity between the structure of the binding site and the structure of the
substrate.
o The structure of the substrate changes during the reaction, becoming first the
transition state and then products.
o The structure of the enzyme can only be complementary to one form of the substrate.
o If the structure of the active site is complementary to the transition state, then binding
increases as the reaction proceeds, which lowers the activation energy.
o Having the active site bind the transition most strongly also means that products are
bound weakly, which favors dissociation of the products from the enzyme once the
reaction is complete.
o When the active site is complementary to the transition state the full substrate binding
energy is only realized as the reaction reaches the transition state, which lowers the
activation energy of the reaction by the magnitude of the binding energy.
Evolution of the maximum rate of an enzyme-catalyzed reaction - strong binding of the
transition state and weak binding of the substrate
o Maximizing KM at constant kcat/KM. Consider a hypothetical case where kcat/ KM =
106 M-1s-1 and [S] = 10-3 M. This table shows that the rate of the reaction = v/[E] =
{[S] kcat]/[KM + [S]} = mols of product formed per mol of enzyme per second is
maximal when KM >> [S].
KM (M)
kcat (s-1)
Rate (s-1)
10-6
1
1
10-5
10
9.9
10-4
102
100
10-3
103
500
10-2
104
909
10-1
105
990
1
106
999
This result contradicts the belief that strong binding of substrate, low KM, is an
important component of enzyme catalysis.
This result is consistent with the principle that enzymes bind the transition state
strongly.
4
The evolution of an enzyme to give maximal rate may be divided into two hypothetical
steps
o The value of kcat/KM is maximized by having the enzyme be complementary to the
transition state.
o The concentration of free enzyme [E] is maximized by having KM be high, which
means that as much of the enzyme as possible is in the free state. This follows from
the fact the Michaelis-Menten equation can be rewritten as v =[E][S] kcat/ KM.
Key regulated enzymes often have a low KM
o A low KM may be advantageous for the first enzyme in a metabolic pathway.
o This would regulate the entry of material into the pathway over a wide range of
substrate concentrations because the enzyme would always operate at maximal
velocity.
Molecular Mechanisms for the Utilization of Binding Energy
Strain is a classic concept in which it was supposed that binding of the substrate to the
enzyme somehow caused the substrate to become distorted toward the transition state.
Transition state stabilization is a more modern concept, which states that it is not the
substrate that is distorted but rather that the transition state makes better contacts with the
enzyme than the substrate does, so the full binding energy is not achieved until the transition
state is reached.
Induced fit assumes that the active site of an enzyme is not complementary to that of the
transition state in the absence of the substrate. Such enzymes will have a lower value of
kcat/KM, because some of the binding energy must be used to support the conformational
change in the enzyme. Induced fit increases KM without increasing kcat.
Strain or stress? It is unlikely that there is enough energy available in substrate binding to
actually distort the substrate toward the transition state.
o It is possible that the substrate and enzyme interact unfavorably and this unfavorable
interaction is relived in the transition state.
o We might think that the substrate is under stress meaning that it is subjected to forces
but not distorted by them.
o It is more likely that the enzyme is strained, as for example in induced fit.
Enzyme Mechanisms
Induced fit
Hexokinase catalyzes the ATP-dependent phosphorylation
of glucose.
The binding of glucose to the enzyme induces a major
conformational change - the induced fit.
5
Phosphoglycerate
synthesis of ATP.
kinase
catalyzes
the
The two substrates are brought together in the
active site by a conformational change.
An induced fit at an interface. Lipases are enzymes that
hydrolyze esters of long chain fatty acids. These enzymes
show virtually no activity against water-soluble substrates
and only hydrolyze substrates present in the interface
between water and lipid - interfacial activation. The
activation of the lipases at the interface is a result of a
conformational change, induced fit, caused by the enzyme
binding to the interface. This conformational change
exposes the active site of the enzyme.
Enzyme Mechanisms
Lysozyme
Lysozyme catalyzes the hydrolysis of the (1 4) linkage between N-acetylmuramic acid
(NAM) and N-acetylglucosamine (NAG) in bacterial cell wall polysaccharides and (1
4)-linked poly NAG (chitin). The lysozyme mechanism illustrates:
o Transition state stabilization
o Acid-base catalysis
o Several residues in the protein participate in substrate binding.
o The binding of NAM4 in the chair conformation is unfavorable.
o But the binding of residue 4 in the half-chair conformation is favorable (preferential
transition state binding).
o Hydrolysis involves acid-base catalysis (Glu35 serves as a proton donor to the
oxygen of the leaving alcohol. The resulting carbonium ion (+) is stabilized by
the ionized side chain of Asp52 until it can react with water)
Many glycosidases utilize a covalent intermediate in their mechanism. Here is a good
example of transition state stabilization using a bacterial glycosidase.
Another excellent example that clearly shows how transition state stabilization works is
found in the structure of this transferase.
Serine proteases
6
Serine proteases are endoproteases that cleave internal peptide
bonds in proteins. The serine protease mechanism illustrates:
o Transition state stabilization.
o Covalent catalysis, involving a catalytic triad of Asp, His
and Ser in the active site.
The currently accepted serine protease mechanism follows below
7
All serine proteases follow the same mechanism. Their specificity is determined by the
nature of the specificity pocket.
In the trypsin specificity pocket, the basic residue of the substrate interacts with an Asp at the
bottom of the pocket.
Chloroketones are inhibitors of serine proteases, which bind in the active site and react with
His 57.
Pancreas contains a small protein (6 kDa) that is a potent inhibitor of trypsin.
The inhibitor is a naturally occurring transition state analog of a protein substrate.
Divergent Evolution of Enzymes - Substrate specificity of pancreatic serine proteases.
The serine proteases all have the same substrate, a polypeptide chain. Different enzymes
cleave the chain adjacent to different amino acids. While the catalytic mechanism remains
the same, altering the amino acids that form the specificity pocket changes the specificity.
Convergent Evolution of Enzymes - Chymotrypsin and Subtilisin.
Subtilisins are a group of serine proteases produced by bacteria. Both chymotrypsins and
subtilisins hydrolyze polypeptide chains adjacent to large hydrophobic amino acid side
chains - they have the same specificity. Yet, the subtilisins and chymotrypsins have no amino
8
acid sequence similarity and their tertiary structures are quite different. Chymotrypsin is
composed of primarily -sheets and the active site lies between two distinct domains.
Subtilisin is an -structure without domains.
Chymotrypsin
Subtilisin
Despite the differences in tertiary structure, the active sites of the two enzymes have very
similar features: The catalytic triad, an oxyanion hole, a specificity pocket (not shown) and
a nonspecific binding region, are common features of the active sites of these two enzymes.
Chymotrypsin
Subtilisin
What is the rest of the protein for?
It is usually stated that "the rest of the protein" is needed to establish the tertiary structure of
an enzyme, which leads to formation of the active site. That this is an oversimplification is
shown by experiments that have attempted convert chymotrypsin to trypsin.
9
o
o
o
o
o
o
Chymotrypsin and trypsin have a very similar tertiary structure and the active site is
also quite similar.
At first glance, it seems that the major difference is in the specificity pocket. In
chymotrypsin the bottom of the specificity pocket is Ser 189, while it is Asp 189 in
trypsin.
So, substituting Asp for Ser at position 189 would seem to be all that would be
necessary to convert chymotrypsin the trypsin, right?
Well, when the experiment was done, what was obtained was an enzyme with very
little activity toward either chymotrypsin or trypsin substrates.
Although extensive substitutions have been made in chymotrypsin, one has yet to
succeed in converting chymotrypsin to trypsin, although enzymes with altered
specificity have been produced.
Clearly, some of "the rest of the protein" is important both in determining specificity
and catalytic efficiency. As yet, we can't offer a fuller explanation.
Zymogen activation - creating the transition state-binding site
What is the basis for the lack of enzymatic activity of a zymogen, such as trypsinogen?
As this Chime routine shows, the zymogen has a fully formed catalytic triad and specificity
pocket, but lacks a functional oxyanion hole, which stabilizes the transition state.
This is yet another example of the critical importance of transition state stabilization as the
primary factor explaining the catalytic activity of enzymes.
Triose phosphate isomerase
The triose phosphate isomerase mechanism illustrates:
o Transition state stabilization
o Acid-base catalysis
o Electrostatic catalysis
The triose phosphate isomerase mechanism is an example of concerted acid-base catalysis.
Many electron transfer reactions involve transferring electrons from one protein to another.
For small molecules that are substrates for enzymes, electrostatic interactions are often
critical.
Many enzymatic reactions are complex and involve more than one substrate
Enzyme Catalysis-Summary
10
Enzymes accelerate reactions using
o General acid catalysis.
o General base catalysis.
o Covalent catalysis.
o Metal ion catalysis.
o Electrostatic catalysis.
o Proximity and orientation effects.
o Preferential Transition State Binding.
Enzymes accelerate chemical reactions by lowering the activation energy, Go‡
Strong binding of the transition state and weak binding of the substrate leads to the maximum
rate because all the binding energy is used to lowerGo‡.
Any process that uses some of the binding energy for another purpose, e.g., a protein
conformational change, will lower the maximum rate.
Under physiological conditions, the maximum rate is achieved by maximizing kcat/KM AND
having KM greater than the physiological [S]. This means that most of the enzyme will be free to
interact with a substrate molecule and that every encounter will lead to a reaction.
11