BY2.3 Part Two - Plants
advertisement

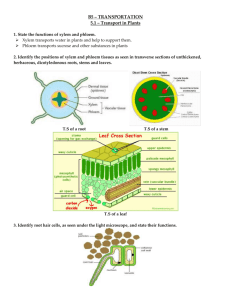
BY2.3 Part 2 – Plants Xylem tissue transports water and mineral salts from the roots to the leaves. Phloem tissue transports the soluble products of photosynthesis (sucrose and amino acids) from the leaves to the other parts of the plant. In each case the walls of tubes are thickened by addition of cellulose and lignin (a wood material) Xylem is made up of four different types of cells: vessels - the main conducting cells, contain pits so that water can travel sideways between one xylem vessel and another (dead) tracheids - also conduct water but are not as well adapted as vessels for this function, joined together with vessel cells to form long narrow tubes, this is where the water is transported (dead) fibres - they have no role in water transport but function in support, they are long, narrow cells with tapered ends (dead) Xylem parenchyma - this acts as packing tissue, stores food and allow lateral transport of materials through vascular tissue (living) Vessels and tracheids are dead cells and they form a system of tubes through which water can travel. The cells are dead because lignin has been deposited on the cellulose cell walls rendering them impermeable to water and solutes. These cells also provide mechanical strength and support to the plant. Tracheid Parenchyma cell Vessel Fibre Lignified wall of vessel Stem Root The distribution of xylem tissue differs in primary stems, leaves and roots: In stems it occurs as part of the peripheral vascular bundles. This organisation gives flexible support but also resistance to bending strain. In leaves the arrangement of vascular tissues in the midrib and network of veins also gives flexible strength and resistance to tearing strains. In roots the central arrangement is ideal for resistance to vertical stresses (pull) and so helps the anchorage of the plant. 1. The large quantities of water lost through the stomata of the leaves by the process of transpiration must be replaced with water from the soil. 2. The region of greatest uptake is the root hair zone where the surface area of the root is enormously increased by the presence of root hairs. 3. The soil water contains a very weak solution of mineral salts and so has a high water potential. 4. The vacuole of the root hair cell contains a strong solution of dissolved substances and has a low water potential. 5. Water passes into the root hair cell down a water potential (WP) gradient from a high WP in the soil to a low WP in the cells by osmosis. 6. The water then continues to move through the plant, across the cortex, to the xylem which has the lowest water potential Root hairs adaptations: - Thin walls Long thin shape Mitochandria (to provide ATP for active transport) Freely permeable Uptake of mineral ions to maintain water potential There are 3 main pathways through which the water can move through from the roots to the xylem: Apoplastic – Goes through non-living parts of the cell (through cell wall) by diffusion/osmosis Symplast – goes through living parts of the cells ( cytoplasm). Cells are linked by cytoplasmic strands that pass through small openings in the cell walls called plasmodesmata -> there is a continuous column of cytoplasm extending from the root hair cells to the xylem Vacuolar /transcellular – water moves from cell to cell via vacuoles. Two main pathways are symplast and apoplast, most water travels by apoplast as it is the fastest. The Endodermis: The endodermis is a layer of cells which surround the pericycle within which lies the vascular tissue (stele) The pericycle is a cylinder of parenchyma cells that lies just inside the endodermis and is the outer most part of the stele of plants Once the water reaches the endodermis (probably by the apoplast pathway), it is blocked by the Casparian Strip The casparian strip is made from water-proof suberin and is found the on radial, lower and upper walls, but not the tangential walls. Therefore, the water has to take the symplast pathway – this means the plant selectively take up which ions enter and leave the xylem. Once in the pericycle, the water retakes the apoplast pathway as the xylem lacks cell contents. Salts are actively secreted into the vascular tissue (xylem) from the endodermal cells, requiring ATP. This makes the WP in the xylem more negative, causing water to be drawn in from the endodermis, so promoting the movement of water into the xylem from the cortex. The water potential gradient produced creates a hydrostatic pressure inside the xylem, known as the root pressure. The force is quite significant in herbaceous plants but is minimal compared to transpirational pull. 3 pieces of evidence for the existence of root pressure: 1. Pressure increases as temperature rises 2. Metabolic inhibitors (e.g. cyanide) that inhibit respiration cause root pressure to cease ( stops the salts being actively secreted) 3. A decrease in O2 availability results in a reduction of root pressure The movement of water from the root to the leaf: Water travels in the xylem up through the stem to the leaves, where most of it evaporates from the internal leaf surface and passes out, as water vapour, into the atmosphere. The transpiration of water from the leaves draws water across the leaf from the xylem tissue along three same pathways as in the root. As water molecules leave xylem cells in the leaf, they pull up other water molecules. This pulling effect is known as the transpiration pull and is possible because of the large cohesive forces between water molecules and the adhesive forces which exist between the water molecules and the hydrophilic lining of the vessels. These two forces combine to maintain the column of water in the xylem. The theory of the mechanism by which water moves up the xylem is known as the Cohesion- Tension theory. Capillarity is another force that may contribute to the rise of water in the xylem. Water rises up narrow tubes by capillary action but this force has more relevance in small plants than large trees. Cohesion – Water molecules sticking together Adhesion – Water molecules sticking to xylem wall 3 Pieces of evidence for cohesion-tension theory: 1. Diameter of tree trunks varies on transpiration rate. Greater levels of transpiration during daylight -> narrower diameter. Reduced transpiration levels at night -> wider diameter. 2. Broken xylem vessels cause a tree to have difficulty drawing up water. Water column is broken so water molecules are not stuck together at this point. Tree would eventually die as water cannot reach higher parts of plant 3. Broken xylem vessels do not leak water (as would expected if there was a positive pressure) but instead air is drawn in (as would be the case with a negative pressure) Transpiration rate – the rate at which water leaves the plant via the stomata, by evaporation Factors affecting the transpiration rate: Temperature - a rise in temperature provides more kinetic energy for the movement of water molecules. This additional energy increases the rate of evaporation of water from the plant and speeds up the rate of diffusion of water vapour into the surrounding atmosphere. The water potential of the atmosphere becomes lower as its temperature is raised and it can hold more moisture. Humidity – if the air outside of the leaf is very humid (i.e. high level of water vapour), then the water potential gradient from inside the leaf to the surrounding atmosphere would be low and so the transpiration rate would be low. However, if it were a particularly dry day, there would be a high water potential gradient and so the water would move down this concentration gradient and leave the plant, increasing the rate of transpiration Air movement - transpiration in still air results in a layer of saturated air at the surfaces of leaves. This offers considerable resistance to the diffusion of water vapour through stomata and thus reduces the rate of transpiration, as the water potential gradient would be low. If it is a windy day, the water molecules that have left the plant are swiftly moved away, causing a high water potential gradient and so a higher transpiration rate. Light intensity - also affects transpiration by controlling the degree of stomatal opening (most stomata open during the day when it’s sunny) In reality, these factors do not act independently but interact with each other, e.g. more water is lost on a dry, windy day than on a humid, still day. This is because the sub-stomatal chamber has a high water potential as the walls of the spongy mesophyll cells are saturated with water. The water evaporates from the walls and moves down a gradient of water potential from the plant to the atmosphere which has a low percentage relative humidity; the wind having reduced the thickness of the layer of saturated air at the leaf surface. The rate of transpiration is measured using a potometer. It actually measures the rate of water absorption but if the cells of the plant are fully turgid the rate of water absorption and the rate of transpiration are the same. The reservoir is used to push the air bubble back to “0” The air bubble moves along the capillary tube as water is absorbed by the shoot – measuring the rate of transpiration Plants can be classified on the basis of their structure in relation to the prevailing water supply, into hydrophytes, mesophytes and xerophytes. Hydrophytes (adapted to living in aquatic environments) - Submerged roots with floating leaves Increased number of stomata on upper side of leaves Thin cuticle as cuticles primarily prevent water loss Less rigid structure – water pressure supports them Large air spaces in stem and leaf tissue for flotation Poorly developed xylem – no need for transport tissue as surrounded by water Mesophytes (terrestrial plants neither adapted to dry or wet environment) Mesophytes grow well with an adequate water supply, but in prolonged dry periods they survive by: - Shedding their leaves to reduce transpiration (e.g. during winter) Dormant seeds The aerial parts of many non-woody plants die off as a result of frost or cold winds but their underground organs survive e.g. bulbs, corms. Xerophytes ( adaptations which allow them to tolerate very dry conditions) Rolled leaves - large thin-walled epidermal cells at the bases of the grooves shrink when they lose water from excessive transpiration, causing the leaf to roll onto itself. This has the effect of reducing the leaf area from which transpiration can occur. This also creates a micro climate which is very humid -> water potential gradient lowered -> diffusion rate lowered Sunken stomata - stomata are found in grooves on the inner side of the leaf. They are located in pits or depressions so that humid air is trapped outside the stomata. This reduces the water potential gradient between the leaf and the atmosphere and so reduces the rate of diffusion of water. Hairs - stiff, interlocking hairs trap water vapour and reduce the water potential gradient. Thick cuticle - the cuticle is a waxy covering over the leaf surface which reduces water loss. The thicker this cuticle the lower the rate of cuticular transpiration. Reduced number of stomata – less area for transpiration Small leave – reduced surface area of leaf for transpiration Succulent steems and leaves – water storage Some close their stomata during the day Shallow extensive root systems to quickly absorb rain water and overnight condensation E.g. Marram Grass Ammophila arenaria (marram grass) is an example of a xerophyte which colonises sand dunes around the coast. The sand dune habitat makes it difficult for a mesophyte to survive there because there is no soil, rapid drainage of rain water occurs, there are high wind speeds, salt spray and a lack of shade from the sun. Translocation The phloem contains a very concentrated solution of dissolved solutes, e.g. sucrose, amino acids. This solution is called cell sap and the transport of solutes in the phloem is called translocation which occurs by mass flow, the bulk movement of substances through the phloem vessels. Phloem is a living tissue and consists of four types of cells: sieve tubes companion cells phloem fibres phloem parenchyma The main ones are companion cells and sieve tubes. The sieve tubes are elongated and their ends are perforated with pores. These areas are called sieve plates. Cytoplasmic filaments/strands containing phloem protein extend from one sieve cells to the next through the sieve plates. The sieve tubes do not possess a nucleus. Each sieve tube is connected to a companion cell by numerous plasmodesmata. Companion cells have many mitochondria used for active transport of sucrose into the Phloem. Mass Flow Theory: Areas in a plant where sucrose is loaded into the phloem are called sources (e.g. leaf) Areas were sucrose is removed from the phloem are known as sinks (a lot of sucrose is removed from the phloem to form starch in the root) Unlike water in the xylem, the contents of the phloem can move up or down a plant stem, often simaultaneously (but not in the same sieve tube) 1) Photosynthesising cells (source cells) produce glucose which is converted into sucrose, which is loaded into the phloem sieve tube by active transport (using ATP) 2) By increasing the level or solutes in the phloem, the water potential is lowered and water moves in from the adjacent xylem by osmosis, down a water potential gradient 3) This raises the hydrostatic pressure in the phloem so that It has a higher pressure 4) At the roots/growing points (sink cells) there is a lower concentration of sucrose. Sucrose is too big to diffuse across the cell membrane so there must be an active process to remove sucrose from sieve tubes. Where it is converted to starch for storage or converted to glucose to be respired. The loss of sucrose from the phloem raises the water potential higher than in the xylem 5) Water enters the xylem by osmosis 6) Water moves from phloem to xylem down a water potential gradient, causing a reduction in the Hydrostatic pressure 7) Water moves up the xylem by transpirational pull 8) Sucrose and dissolved solutes move by mass flow from a high to low hydrostatic pressure, down a pressure gradient Translocation experiments Aphid stylet experiments o Aphids, such as greenfly, have specialised mouthparts called stylets, which they use to penetrate phloem tubes and sup of the sugary sap therein. o If the aphids are anaesthetised with carbon dioxide and cut off, the stylet remains in the phloem so pure phloem sap can be collected through the stylet for analysis. o This surprising technique is more accurate than a human with a syringe and the aphid’s enzymes ensure that the stylet doesn’t get blocked Ringing experiments o Since the phloem vessels are outside the xylem vessels, they can be selectively removed by cutting a ring in a stem just deep enough to cut the phloem but not the xylem. o After a week there is a swelling above the ring, reduced growth below the ring and the leaves are unaffected. This was early evidence that sugars were transported downwards in the phloem. Using Radioactive tracers o Radioactive isotopes can be used trace precisely where different compounds are being transported from and to, as well as measuring the rate of transport. The radioactivity can be traced using photographic film (an autoradiograph). This technique can be used to trace sugars, ions or even water. o In a typical experiment a plant is grown in the lab and one leaf is exposed for a short time to carbon dioxide containing the radioactive isotope 14C. This 14CO2 will be taken up by photosynthesis and the 14C incorporated into glucose and then sucrose o The plant is then frozen in liquid nitrogen to kill and fix it quickly, and placed onto photographic film in the dark. The resulting autoradiograph shows the location of compounds containing 14C. Problems with the mass flow hypothesis Mass flow is unable to describe for movement in opposite directions at the same time and different rates of sucrose flow Other hypotheses include: o Diffusion o Cytoplasmic streaming