Open Access version via Utrecht University Repository

Serotonin, and probably not Angiotensin II, mediates cyclic strain-induced expression of transforming growth factor-beta 1 in mitral valves.
Annelies Elema
1
, Carla M.R. Lacerda
2
, Maarten van Emst
1
, E.
Christopher Orton
2
.
1 Department of Pathobiology, Utrecht University, Utrecht, The Netherlands
2 Department of Clinical Sciences, Colorado State University, Fort Collins, CO, United States
……………………………………
Abstract
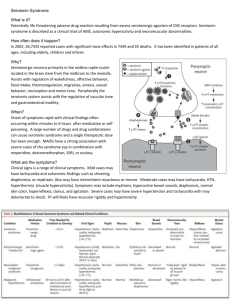
Degenerative (myxomatous) mitral valve disease is a leading cause of disability and death in dogs and humans. Transforming growth factor-beta 1 (TGFβ1) is strongly implicated in the pathogenesis of degenerative mitral valve disease, however initiating stimuli and signaling mechanisms that mediate TGFβ1 expression in mitral valves are not known. This study tested the hypothesis that high tensile strain on mitral valves initiates local synthesis of serotonin and angiotensin II, and that these in turn would act synergistically to increase TGFβ1 expression. Methods and Results: Cultured sheep mitral valves were subjected to 0%
(control), 10% (low physiologic), 20% (high physiologic), and 30% (pathologic) cyclic strain for 24 and 72 h in serum-free culture media. Serotonin and angiotensin II concentrations were measured in the culture media by ELISA. TGFβ1 abundance was measured by Western blot.
Angiotensin II levels were significantly decreased in the media at every step of increasing strain at 24 h and between 10 % and 20% strained mitral valves at 72 h. Serotonin levels were significantly increased in the media of 20% and 30% strained mitral valves at 24 and 72 h.
Pharmacologic inhibition of serotonin with 10 μM SB-206553 (serotonin type 2 receptor blocker) diminished expression of TGFβ1 in 30% strained mitral valves. Conclusion:
Serotonin mediates cyclic strain-induced expression of TGFβ1 in mitral valves. The role of angiotensin is uncertain. These results could point to possible therapeutic strategies to slow progression of degenerative mitral valve disease.
……………………………………
Degenerative (myxomatous) mitral valve disease (DMVD) is a leading cause of disability and death in dogs and humans.
1,2 It is characterized by 1) significant mitral regurgitation (MR) secondary to leaflet prolapse or flail, 2) gross thickening and redundancy of the leaflets with chordate lengthening or rupture, and 3) myxomatous pathology of the valve apparatus.
Myxomatous pathology is characterized by exuberant accumulation of proteoglycan (PG)/ glycosaminoglycan (GAG) in all leaflet layers, fragmentation of elastin fibers, and disarray of collagen bundles, causing degradation of the extracellular matrix (ECM), myofibroblast proliferation, fibrosis, and myxoid matrix deposition.
3,4
At the moment little is known about the cause of DMVD. Transforming growth factor-beta 1
(TGFβ1) has been shown in several studies to play a part in the pathogenesis of DMVD and ventricular remodelling.
5-8
Aupperle et al
5
showed that in normal mitral valves TGFβ3 was expressed by 50% of the interstitial cells and in contrast, TGFβ1 was detected in only 10% of the valvular interstitial cells (VICs) and TGFβ2 in some individual cells. In canine DMVD
TGFβ1, TGFβ3 and α smooth muscle actin (α-SMA) are overexpressed. As DMVD
progressed in severity, the expression of TGFβ1 and TGFβ3 increased markedly within the cells. TGFβ1 was also found within the altered ECM, and TGFβ3 was not found.
5
Under pressure overload, TGFβ1 plasma levels were also significantly increased 8 . TGFβ1 is known to transform fibroblast into metabolically active and functional myofibroblasts
5
. The signaling pathways that mediate strain-induced elevations of TGFβ1 are yet to be discovered.
The heart valve consists of VICs. VICs are located within the matrix and endothelial cells.
Characterization of the cellular composition of valve leaflets show a heterogeneity of VICs.
There are at least three phenotypes of VICs identified in the mature native valve. Fibroblast, myofibroblast and smooth muscle cell.
9
The myofibroblast is characterized by prominent stress fibers, associated with smooth muscle α-actin expression and is thought to be involved in proliferation and migration, but they also synthesize collagen and other matrix proteins.
9-12
Disatian et al
13
showed there are more fibroblast phenotype VICs in normal mitral valves and the phenotype of interstitial cells with progressive DMVD showed markedly higher myofibroblast phenotype. Komuro
14
also proposed that a family of fibroblast-like cells exist which varies its phenotype as an adaptive response to the microenvironment.
ANGII is well considered to be an important mediator for cardiac remodeling, including hypertrophy and fibrosis in response to hemodynamic overload.
16-18
The renin-angiotensinaldosterone system (RAAS) and TGFβ1 are key mediators for this transformation.
18,19
They do not act independently from one another but rather act as part of a network that promotes cardiac remodeling. ANGII induces the expression of TGFβ1 via angiotensin type 1 (AT
1
) receptor in ventricular muscle cells.
20,21 In vitro studies have shown that TGFβ1 mRNA and protein are readily upregulated by ANGII in cardiac fibroblasts, myofibroblasts and myocytes 10,19,22-25 . RAAS is activated in response to hemodynamic overload, and that activation of the (local) RAAS contributes to myocardial hypertrophy, fibrosis, and dysfunction.
10,19
In addition, a large number of animal studies and clinical trials in humans have shown that inhibition of ANGII by ACE inhibitors or AT
1
receptor antagonists prevents or reverses ventricular remodeling and improves survival in patients with heart failure.
19
During cardiac remodeling, morphologically distinct cells termed myofibroblasts appear in the myocardium
10
. As said before TGFβ1 is known to transform fibroblast into metabolically active and functional myofibroblasts 5 and self-amplifies its expression in myofibroblasts.
The role of ANGII in pathologic remodeling of heart valves is less well understood. The known role of ANGII in activating TGFβ1 in myocardium suggests a possible role for ANGII in the pathologic remodeling of DMVD.
Next to the many publications that show the influence of TGFβ1 on DMVD
5-8
Obayashi et al
6
showed that addition of angiotensin converting enzyme inhibitor (ACEI) to cultured VICs decreased the concentration of TGF-β3 significantly. This supports a hypothesis that ANGII mediates TGF-β in heart valves.
Studies by Disatian et al
7
, Jian et al
26
, Waltenberger et al
27
, Lacerda et al
28 and Hutcheson et al
29 indicate a possible role for serotonin in the development of DMVD as well. Until recently it was thought that serotonin was primarily synthesized in the gut and the brain, but there are strong indications that serotonin is also made locally by the heart valves 7,26,28,30 . Serotonin is regulated by tryptophan hydroxylase 1 (TPH1). This is the limiting enzyme for peripheral serotonin synthesis and TPH1 is found to be produced by VICs. This implicates an autocrine serotonin signaling mechanism. TPH1 was upregulated in DMVD 30 . Serotonin provides, via
G protein coupled activation of second messengers, for various actions including the transcription of TGFβ1 and therefore possibly the development of DMVD
7,26-28
.
In this study we investigated the role of ANGII and serotonin in mediating the abundance of
TGFβ1 in cultured mitral valves subjected to different levels of cyclic tensile strain.
Material and methods
Cultured mitral valves of sheep cadavers were used. The valves were stretched uniaxial in a specific developed machine (Figure 1). This machine can mimic the effects of a heartbeat on only the valves. The percentage of strain on the valve can be controlled. The valves were kept in a serum-free DMEM-F12 culture medium supplemented 10% Sigma Serum Replacement
2 (albumin, transferring, insulin). The machine was contained in an incubator set at a constant temperature of 37°C and 5% CO
2
- 95% air.
The valves were subjected to 0% (control), 10% (low physiologic), 20% (high physiologic), or 30% (pathologic
31-33
) cyclic strain, either for 24 or 72 hours. A sinusoidal dynamic cycling protocol at 0,5 Hz was used. Applied tensile load was measured every 30 min during each experiment. Figure 2 shows that during time less load is needed to maintain the same strain on the valve.
Figure 1: Heart valve stretch machine; Apparatus for applying cyclic strain to cultured mitral valves. Valve leaflets were mounted onto custom-built devices with clamps to fix the leaflets and placed into a 4-well culture chamber (A). A miniaturized axial loading machine capable of delivering precise tensile cyclic strain and measuring tensile load on cultured mitral valves was housed within a standard incubator (B). Compression of the chamber lid by the axial loading machine increases the distance between the clamps of the valve holders resulting in tensile strain on the mounted valve leaflets.
Figure 2: Representative curves of measured tensil load necessary to maintain 10, 20 and 30% strain on cultured mitral valves over 30h.
ELISA
Specific ELISAs were used for the assessment of the amount of both serotonin and
ANGII in the culture medium of strained mitral valves. The manufacturer instructions were used (ADI-900-204 and ADI-900–175; Enzo Life Sciences, Plymouth Meeting, PA).
For the ELISA of ANGII, the manufacturer’s standards and our samples were added to wells coated with a goat-anti-rabbit IgG antibody. A yellow solution of rabbit polyclonal antibody to ANGII was added. The plate was incubated at room temperature for a hour. A blue solution of ANGII conjugated to biotin was added and incubated again for a hour at room temperature. During this incubation the specific antibody binds to the ANGII in the sample or conjugate. The plate was washed with washing buffer, leaving only bound
ANGII or conjugate. A solution of streptavidin conjugated to horseradish peroxidise
(HRP) was added to each well, to bind the biotinylated angiotensin. The plate was again incubated for a hour. After incubation the plate was again washed with washing buffer to remove unbound HRP conjugate. Tetramethylbenzidine (TMB) substrate solution was added. An HRP-catalyzed reaction generates a blue color in the solution. This substrate reaction had a half hour to work after it was stopped by adding a stop solution (a solution of hydrochloric acid in water). The resulting yellow color was read by a spectrophotometer at 450 nm. The amount of signal is inversely proportional to the level of ANGII in the sample. The ELISA for serotonin followed the same principle.
Concentrations of ANGII and serotonin were calculated using a standard curve of known concentrations supplied by the manufacturers.
Results of ANGII and serotonin concentrations were analyzed by one-way ANOVA and student T-test. A two-sided P-value <0,05 was considered significant.
Inhibition of Serotonin
The valve underwent a cyclic strain of 30% for 72 hours. During this time serotonin was inhibited by the tryptophan hydroxylase inhibitor 4-chloro-DL-phenylalanine. A concentration of 250 µM was used.
Western blot
Western blot was used to look at the different levels of the protein TGFβ1. This was measured from the proteins obtained from the valve tissue lysate. Strained valves were compared with and without serotonin receptor blockers. The TGFβ1proteins in the media were separated using gel electrophoresis. The samples and a marker were loaded in equal amounts (40µg/lane) into wells in the gel (4-12% Tris-glycine polyacrylamide; Invitrogen,
Carlsbad, CA). The proteins migrated through the gel at different speeds, depending on their size, when voltage is applied along the gel. This will result in different bands. The next step was to transfer the separated proteins in the gel onto a membrane made of nitrocellulose
(Invitrogen) by electro blotting. The membranes were blocked with 1% BSA for 30 minutes to bind to any remaining places on the nitrocellulose where the target proteins have not attached and then incubated overnight with non-fat dry milk along with mouse monoclonal antibody, which binds to its specific protein. The protein band densities are quantified on
Image J and compared by ANOVA: single factor. A P-value <0,05 was considered significant.
Results
ELISA Angiotensin II
The results of the ELISA of ANGII are found in table 1. The optical density (OD) found by the ELISA is converted to ANGII concentration in nanogram per milliliter using a standard curve. The two-sided P value showed that except for 20% strain (P = 0,015), there is no significant between 24h and 72h within a strain group. The two-sided P value for medium is
0,94, 0% is 0,21, 10% 0,20 and 30% strain has a value of 0,42.
The results show that there was already ANGII present in the medium. In the serum the concentration of ANGII has a tendency to decrease when the strain goes up (Fig 2). The
ANOVA showed that there was a significant difference within the 24h group (P = <0,001) as well as within the 72h group (P = <0,001).
The 24 hours group showed no significant difference (P = 0,38) between the medium and non-strained, but every rise in tensil strain leads to a significant decline in ANGII concentration. There is a significant (P = <0,01) decrease of the serum level of ANGII between non-strained and 10% strained. Also between 10% and 20% strain and between
20% and 30% strain the level of ANGII in the serum is significant lower (P = 0,026 resp.
P = <0,01).
For 72 hours the tendency is the same. It also decreases when the strain goes up. There is again no significance between the medium and non-strained group (P = 0,081). There is a significance between the medium and 10% with P = < 0,01. And between 10 and 20% there is a significance of P = 0,015. There is no significance between 0 and 10% (P=
0,057) and also not between 20 and 30% strain (P = 0,10).
Table 1: The angiotensin II concentration in ng/ml
24 h
Medium 0% 10% 20% 30%
72 h
Medium 0% 10% 20% 30%
N =
Mean
STDEV
0,30 0,30 0,16 0,04
0,40 0,33 0,12 0,06
0,37 0,41 0,08 0,07
0,39 0,27 0,12 0,05
4 4 4 4
0,36 0,33 0,12 0,06
0,05 0,06 0,03 0,01
0,01
0,02
0,01
0,03
4
0,02
0,01
0,37 0,20 0,10 0,01 0,01
0,33 0,21 0,05 0,02 0,01
0,33 0,39 0,10 0,04 0,02
0,44 0,18 0,11 0,03 0,02
4 4 4 4 4
0,37 0,24 0,09 0,03 0,02
0,05 0,10 0,03 0,01 0,00
Figure 2: Concentration of Angiotensin II in ng/ml, measured in the media of mitral valves subjected to 0, 10,
20 or 30% cyclic strain for 24 or 72 hours. * Significant difference from 0% strain. ** Significant difference from 10% strain. *** Significant difference from 20% strain. P < 0,05 is considered significant. Data are means
± SD. N = 4 for every group.
ELISA serotonin
The results of the ELISA of serotonin are found in table 2. The optical density (OD) found by the ELISA is converted to serotonin concentration in nanogram per milliliter using a standard curve. The two-sided P value showed that, except for 10% strain (P =
0,026), there is no significance between 24h and 72h within a strain group. The two-sided
P value for 0% is 0,81, 20% 0,53 and 30% strain has a value of 0,33.
In the serum the concentration of serotonin has a tendency to increase when the strain goes up (Fig 3). The ANOVA showed that there was a significant difference within the
24h group (P = <0,001) as well as within the 72h group (P = <0,001).
The 24 hours group showed significance between 10 and 20% (P = <0,001). There is no significance (P = 0,99) between 0% and 10 % strained valves and also no significant difference between 20% and 30% strain (P = 0,13).
For 72 hours group has a significant decrease in concentration between 0% and 10% strain (P
= 0,015). Between 10% and 20% there is a significant increase (P = <0,001). This increase leads also to a significance between 0 and 20% (P = <0,001). There is no significance between 20 and 30% (P = 0,43).
Table 2: Serotonin concentration in ng/ml
24 h
0%
72 h
10% 20% 30% 0% 10% 20% 30% n 12 12 12 12 12 12 12 12
Mean
STDEV
2,06 2,07 3,67 4,55 2,04 1,29 3,61 4,06
0,42 0,80 1,08 1,53 0,32 0,81 1,20 1,17
Serotonin
7
*
6
24 h
72 h
5 * **
**
4
3
2
1
*
0
0% 10% 20% 30%
Strain
Figure 3: Concentration of serotonin in ng/ml, measured in the media of mitral valves subjected to 0, 10, 20 or
30% cyclic strain for 24 or 72 hours. * Significant difference from 0% strain. ** Significant difference from 0 and 10% strain. P < 0,05 is considered significant. Data are means ± SD. Per group n=12.
ELISA serotonin inhibition
The serotonin level in the serum of 72h with 30% cyclic strain was 2,716 ng/ml with a standard deviation of 0,825. This was significant (P = <0,001) higher than the 72 hours with
30% cyclic strained valves with the TPH inhibitor. Here the average was 1,072 ng/ml with a standard deviation of 0,195. For each group there were eight valves used.
4
Serotonin inhibition
3
2
1
*
0
Strain Strain + TPH inhibitor
Figure 4: The concentration of serotonin, 72h strained heart valves after it has been inhibited with 4-chloro-DLphenylalanine (TPH inhibitor). * Significant difference (P < 0.05).
Western blot TGF-β1
The western blot shows that there is less expression of TGFβ1 when serotonin signaling is inhibited with 10 µM SB-206553 (serotonin type 2 receptor blocker) compared with no inhibition. After quantification by Image J and calculation by ANOVA there was a statistically significant difference between the two groups (P = 0.006) (Fig 5).
Figure 5: Western blot of TGFβ1 expression in the 72h 30% cyclic strained heart valves; Expression of TGFβ1 was decreased after inhibition of serotonin signaling with 10 uM SB-206553 (serotonin type 2 receptor blocker).
Statistically significant difference between groups was indicated with a P-value of <0,05.
Table 3: Results of the Western blot calculated by Image J.
Area Percent
No inhibition 1 2.043.577 23.193
2 1.944.497 22.069
3 1.900.456 21.569
Inhibited with
5HT-receptor blocker
4 608.577 6.907
5 1.164.941 13.221
6 1.149.113 13.042
Discussion
The aim of this study was to determine the influence of different levels of cyclic tensile strain applied to mitral valves on the concentration of ANGII and serotonin in culture medium. If serotonin was depended on local TPH and we also tested the effects of strain and serotonin receptor inhibition on the abundance of TGFβ1 in mitral valves.
Sadoshima et al
34
showed that release and action of ANGII is triggered by strain in cardiac myocytes. Sadoshima et al
34
also showed that the concentration of ANGII in the medium of cultured cardiac myocytes would transiently go up during strain, but not in the medium of cultured fibroblasts. VICs are classified more as myofibroblasts. They have myocyte like
(contraction) and fibroblast like (ECM-syntheses) functions.
9-12
VICs contract in response to epinephrine and ANGII stimulation in vitro.
11,15
ANGII and TGFβ1 are well known for their role in cardiac hypertrophy and remodeling. ANGII is known to activate TGFβ1 10,16-19 .
Because TGFβ1 is strongly implicated in DMVD
5-8
it was expected that ANGII levels would rise once strain was put upon the valve leaflets. However, this study showed that the concentration of ANGII in the medium of mitral valves decreases in response to increasing tensile strain. In the 24h group each increase in strain causes a significant lower ANGII concentration. Also, in the 72h group there is a significant drop between 10% and 20% strain.
Possibly the serum-free media of cultured valves does not contain sufficient ANGI and is therefore not able to produce sufficient ANGII. On the other hand, several studies show an indication of a local RAAS, but most of them where done in cardiac cells
14,15,34
and renal cells
35,37
. The study of Katwaet al
38
showed a local production of ANGII in VICs.
In ventricular myocytes there are vacuoles filled with ANGII 34 . If these vacuoles are also present in VICs this could explain the transient rise of ANGII found by Sadoshimaet al
34
and not in our study.
Another reason could be that ANGII is re-located intracellular in the valves by endocytosis 39-
42
. There is a nuclear AT
1
receptor
35,37
and Li et al
37
found that intracellular ANGII induces
TGFβ1. It is therefore possible that the decrease in serum levels is due to the fact that it is used intracellular. To find out if this happens the ELISA should be done on homogenized samples.
The question then remains whether an increase in intracellular ANGII would trigger an increase in TGFβ1. This should be tested by measuring TGFβ1 and by adding angiotensin receptor blockers (ARBs). A next step could be to use both ARBs and serotonin type 2 receptor blocker to see if the decrease of TGFβ1 is enhanced, compared to only serotonin type 2 receptor blocker or ARBs.
The ELISA measures free ANGII and not bound ANGII. For ANGII to activate its second messenger it binds to a receptor. When it is bound to AT receptors the ELISA does not measure it. The ANGII concentration in the media could go down because it is bound to the receptors and these receptors are upregulated. Goette et al
43
showed that in tissue samples from human patients with atrium fibrillation the AT
1
receptor is downregulated to about 50%, but the AT
2
receptors are upregulated to 500%. Only these valves were diseased valves and it is not sure if the upregulation already takes place within 24/72h. Kijima et al
44 showed in their study that strain of cardiac myocytes leads to upregulation of AT
1
receptor. It could be a reason why there is a significant drop in ANGII when strain goes up.
Thus these results do not exclude a role for ANGII in DMVD, since there was a significant difference between the strain groups. More tests are needed to explain the decrease in ANGII levels in the medium and understand the influence of ANGII on DMVD.
The ELISA for serotonin on the other hand showed that serotonin levels increased in response to increasing valve strain. The serotonin levels of 20 and 30% were significant higher than the serotonin levels of 0 and 10% strain. So we can conclude that strain on the valves induces elevation of serotonin. Since we isolated the valve from circulation we can conclude that the elevation of serotonin comes from local production. This conclusion is supported by the local presence of TPH1 7,30 which could be blocked by TPH inhibitor.
Serotonin provides for various actions via G protein coupled activation of second messengers, which leads among other things leads to the expression of TGFβ1 and therefore development of DMVD.
7,26-28
Higher levels of serotonin induce higher levels of TGFβ1 as shown by the significant effect of serotonin receptor blockade on TGFB1 levels. Therefore it can be said that (pathologic) strain-induced expression of TGFβ1 is dependent on serotonin.
In summary, angiotensin II decreases in cultured mitral valves in response to elevated cyclic tensile strain and more research is needed to examine the relevance of this decrease on
DMVD. Serotonin on the other hand increases in response to elevated strain and a serotonin receptor blocker decreases the abundance of TGFβ1 in cultured mitral valves. These results could point to possible therapeutic strategies to slow progression of degenerative mitral valve disease.
References
1.
LUNG, B., VAHANIAN, A. (2011) Epidemiology of valvular heart disease in the adult, Nat Rev Cardiol, 8 : p162-172.
2.
ATKINS, C., BONAGURA, J., ETTINGER, S., FOX, P., GORDON, S.,
HAGGSTROM, J., HAMLIN, R., KEENE, B., LUIS-FUENTES, V., STEPIEN, R.
(2009) Guidelines for the diagnosis and treatment of canine chronic valvular heart disease, J Vet Intern Med, 23 : p1142-1150.
3.
PELLERIN, D., BRECKERS, S., VEYRAT, C. (2002) Degenerative mitral valve disease with emphasis on mitral valve prolapse, Heart, 88 Suppl 4: iv20-28.
4.
SCHOEN, F.J. (2005) Cardiac valves and vavular pathology: Update on function, disease, repair, and replacement, Cardiovasc Pathol, 14 : p189-194.
5.
AUPPERLE, H., MARZ, I., THIELEBEIN, J., SCHOON, H.A. (2008) Expression of transforming growth factor-beta1, -beta2 and -beta3 in normal and diseased canine mitral valves, J Comp Pathol, 139 : p97-107.
6.
OBAYASHI, K., MIYAGAWA-TOMITA, S., MATSUMOTO, H., KOYAMA, H.,
NAKANISHI, T., HIROSE, H. (2011) Effects of transforming growth factor-beta3 and matrix metalloproteinase-3 on the pathogenesis of chronic mitral valvular disease in dogs, Am J Vet Res, 72 : p194-202.
7.
DISATIAN, S., ORTON, E.C. (2009) Autocrine serotonin and transforming growth factor beta 1 signaling mediates spontaneous myxomatous mitral valve disease, J
Heart Valve Dis, 18 : p44-51
8.
VILLAR, A.V., COBO, M., LLANO, M., MONTALVO, C., GONZÁLEZ-
VÍLCHEZ, F., MARTÍN-DURÁN, R., HURLÉ, M.A., NISTAL, J.F. (2009) Plasma levels of transforming growth factor-beta1 reflect left ventricular remodeling in aortic stenosis.
PLoS One, 4 :e8476
9.
TAYLOR, P.M., BATTEN, P., BRAND, N.J., THOMAS, P.S., YACOUB, M.H.
(2003) The cardiac valve interstitial cell.
Int J Biochem & Cell Bio, 35 : p113-118
10.
VAN WAMEL, A.J., RUWHOF, C., VAN DER VALK-KOKSHOORN, L.J.M.,
SCHRIER, P.I., VAN DER LAARSE, A. (2002) Stretch-induced paracrine hypertrophic stimuli increase TGF-β
1 expression in cardiomyocytes . Mol
CellBiochem, 236 : p147–153
11.
FILIP, D.A., RADU, A., SIMIONESCU, M. (1986) Interstitial cells of the heart valves possess characteristics similar to smooth muscle cells.
Circ Res, 59 : p310-320
12.
BERK, B.C., FUJIQRA, K., LEHOUX, S. (2007) ECM remodeling in hypertensive heart disease. J Clin Invest, 117 : p568-575
13.
DISATIAN, S., EHRHART III, E.J., ZIMMERMAN, S., ORTON, E.C. (2008)
Interstitial cells from dogs with naturally occurring myxomatous mitral valve disease undergo phenotype transformation. J Heart Valve Dis, 17 : p402-412
14.
KOMORO, T. (1990) Re-evaluation of fibroblasts and fibroblast-like cells. Anat
Embryol, 182 : p103-112
15.
KATWA, L.C., RATAJSKA, A., CLEUTJENS, J.P.M., SUN, Y., ZHOU, G., LEE,
S.J., WEBER, K.T. (1995) Angiotensin converting enzyme and kininase-II-like
activities in cultured valvular interstitial cells of the rat heart. Cardiovasc Res, 29 : p57-64
16.
FAN, D., TAKAWALE, A., BASU, R., PATEL, V., LEE, J., KANDALAM, V.,
WANG, X., OUDIT, G.Y., KASSIRI, Z. (2014) Differential role of TIMP2 and
TIMP3 in cardiac hypertrophy, fibrosis, and diastolic dysfunction. Cardiovasc Res,
Epub
17.
QI, G., JIA, L., LI, Y., BIAN, Y., CHENG, J., LI, H., XIAO, C., DU, J. (2011)
Angiotensin II infusion-induced inflammation, monocytic fibroblast precursor infiltration, andcardiacfibrosisare pressure dependent. Cardiovasc Toxicol, 11 : p157-
167
18.
ZHANG, H.,WU, J.,DONG, H.,KHAN, S.A.,CHU, M.L.,TSUDA, T. (2014) Fibulin-
2 deficiency attenuates angiotensin II-inducedcardiac hypertrophyby reducing transforming growth factor-βsignalling. ClinSci (Lond) 126 : p275-88
19.
ROSENKRANZ S. (2004) TGF-beta1 and angiotensin networking in cardiac remodeling. Cardiovasc Res, 63 : p423-432
20.
EVERETT, A.D., TUFRO-MCREDDIE, A., FISHER, A., GOMEZ, R.A. (1994)
Angiotensin receptor regulates cardiac hypertrophy and transforming growth factorbeta 1 expression . Hypertension, 23 : p587-592
21.
KUPFAHL, C., PINK, D., FRIEDRICH, K., ZURBRUGG, H.R., NEUSS, M.,
WARNECKE, C., FIELITZ, J., GRAF, K., FLECK, E., REGITZ-ZAGROSEK, V.
(2000) Angiotensin ii directly increases transforming growth factor beta1 and osteopontin and indirectly affects collagen mrna expression in the human heart.
Cardiovasc Res, 46 : p463-475
22.
GRAY,M.O.,LONG,C.S.,KALINYAK,J.E.,LI,H.T.,KARLINER,J.S.
(1998) Angiotensin II stimulates cardiac myocyte hypertrophy via paracrine release of TGF-beta 1 and endothelin-1 from fibroblasts. Cardiovasc Res, 40 : p352–363
23.
CAMPBELL, S.E., KATWA, L.C. (1997) Angiotensin II stimulated expression of transforming growth factor-beta 1 in cardiac fibroblasts and myofibroblasts.
J Mol
Cell Cardiol, 29 : p1947–1958
24.
FISHER, S.A., ABSHER, M.
(1995) Norepinephrine and angiotensin II stimulate secretion of TGF-beta by neonatal rat cardiac fibroblasts in vitro.
Am JPhysiol, 268 : p910–917
25.
LEE,A.A.,DILLMANN,W.H.,MCCULLOCH,A.D.,VILLARREAL,F.J.
(1995) Angiot ensin II stimulates the autocrine production of transforming growth factor-beta 1 in adult rat cardiac fibroblasts . J Mol Cell Cardiol, 27 : p2347–2357
26.
JIAN, B., XU, J., CONNOLLY, J., SAVANI, R.C., NARULA, N., LIANG, B.,
LEVY, R.J. (2002) Serotonin mechanisms in heart valve disease i: Serotonin-induced up-regulation of transforming growth factor-beta1 via g-protein signal transduction in aortic valve interstitial cells.
Am J Pathol, 161 : p2111-2121
27.
WALTENBERGER, J., LUNDIN, L., OBERG, K., WILANDER, E., MIYAZONO,
K., HELDIN, C.H., FUNA, K. (1993) Involvement of transforming growth factor-beta in the formation of fibrotic lesions in carcinoid heart disease . Am J Pathol, 142 : p71-
78
28.
LACERDA, C.M.R., KISIDAY, J., JOHNSON, B., ORTON, E.C. (2012) Local serotonin mediates cyclic strain-induced phenotype transformation, matrix degradation, and glycosaminoglycan synthesis in cultured sheep mitral valves. Am J
Physiol, 302 : p1983-1990
29.
HUTCHESON, J.D., SETOLA, V., ROTH, B.L., MERRYMAN, W.D. (2011)
Serotonin receptors and heart valve disease – It was meant 2B.
Pharmacology &
Therapeutics, 132 : p146-157
30.
DISATIAN, S., LACERDA, C., ORTON, E.C. (2010) Tryptophan hydroxylase 1 expression is increased in phonotype-altered canine and human degenerative myxomatous mitral valves.
J Heart Valve Dis, 19 : p71-78
31.
KRISHNAMURTHY, G., IOTH, A., BOTHE, W., SWANSON, J.C., KUHL, E.,
KARLSSON, M., CRAIG MILLER, D., INGELS, N.B. Jr (2009) S tress-strain behavior of mitral valve leaflets in the beating ovine heart. J Biomech, 42 : p1909-
1916
32.
SACKS, M.S., ENOMOTO, Y., GRAYBILL, J.R., MERRYMAN, W.D.,
ZEESHAN, A., YOGANATHAN, A.P., LEVY, R.J., GORMAN, R.C., GROMAN,
J.H. III (2006) In-vivo dynamic deformation of the mitral valve anterior leaflet. Ann
Thorac Surg, 82 : 1369-1377
33.
BALANCHANDRAN, K., SUCOSKY, P., JO, H., YOGANATHAN, A.P. (2009)
E levated cyclic stretch alters matrix remodeling in aortic valve cusps: implications for degenerative aortic valve disease. Am J Physiol Heart Circ Physiol, 296 : pH756-
H764
34.
SADOSHIMA, J., XU, Y., SLAYTER, H.S., IZUMO, S. (1993) Autocine release of angiotensin II mediates stretch-induced hypertrophy of cardiac myocytes in vitro .
Cell, 75 : p977-984
35.
KUMAR, R., THOMAS, C.M., YONG, Q.C., CHEN, W., BAKER, K.M. (2012) The intracrine renin-angiotensin system . Clin Sci, 123 : 273-284
36.
YAMADA, H., FABRIS, B., ALLEN, A.M., JACKSON, B., JOHNSTON, C.I.
MENDELSOH, A.O. (1991) Localization of angiotensin converting enzyme in rat heart. Circ Res, 68 : p141-149
37.
LI, X.C., ZHUO, J.L. (2008) Intracellular ANG II directly induces in vitro transcription of TGF-β1, MCP-1, and NHE-3 mRNAs in isolated rat renal cortical nuclei via activation of nuclear AT
1a
receptors. Am J Physiol Cell Physiol, 294 : p1034-1045
38.
KATWA, L.C, TYAGI, S.C., CAMPBELL, S.E., LEE, S.J., CICILA, G.T., WEBER,
K.T. (1996) Valvular interstitial cells express angiotensinogen and cathepsin D, and generate angiotensin peptides. Int J Biochem Cell Bid, 28 : p807-821
39.
GONZALEZ-VILLALOBOS, R., KLASSEN, R.B. ,ALLEN, P.L., NAVAR, L.G.,
HAMMOND, T.G. (2005) Megalin binds and internalizes angiotensin II. Am J
Physiol Renal Physiol, 288 : p420-427
40.
LI, X.C., HOPFER, U., ZHUO, J.L. (2009) AT1 receptor-mediated uptake of angiotensin II and NHE-3 expression in proximal tubule cells through a microtubule-dependent endocytic pathway. Am J Physiol Renal Physiol, 297 : p1342-1352
41.
KATS, VAN J.P., LANNOY, DE L.M., JAN DANSER, A.H., MEEGEN, VAN
J.R., VERDOUW, P.D., SCHALEKAMP, M.A.D.H. (1997) Angiotensin II type 1
(AT
1
) receptor–mediated accumulation of Angiotensin II in tissues and its intracellular half-life in vivo. Hypertension, 30 : p42-49
42.
KATS, VAN J.P., DANSER, A.H.J. MEEGEN, VAN J.R., SASSEN, L.M.A.,
VERDOUW, P.D., SCHALEKAMP, M.A.D.H. (1998) Angiotensin production by the heart: a quantitative study in pigs with the use of radiolabelded angiotensin infusions. Circ, 98 : p73-81
43.
GOETTE, A., ARNDT, M., RÖCKEN, C., SPIESS, A., STAACK, T., GELLER,
J.C., HUTH, C., ANSORGE, S., KLEIN, H.U., LENDECKEL, U. (2000) Regulation of Angiotensin II Receptor Subtypes During Atrial Fibrillation in Humans.
Circulation, 101 : p2678-2681
44.
KIJIMA, K., MATSUBARA, H., MURASAWA, S., MARUYAMA, K., MORI,
Y., OHKUBO, N., KOMURO, I., YAZAKI, Y., IWASAKA, T., INADA, M.
(1996) Mechanical stretch induces enhanced expression of angiotensin II receptor subtypes in neonatal rat cardiac myocytes.
Circ Res, 79 : p887-897