Supplementary Materials and Methods
advertisement

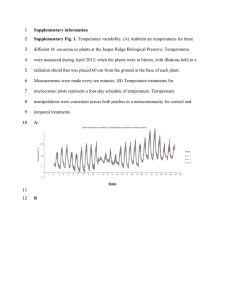
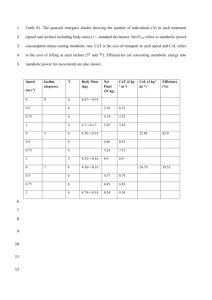
Electronic Supplementary Material for Warmer Ambient Temperature Depress Liver Function in a Mammalian Herbivore authored by Patrice Kurnath and M. Denise Dearing MATERIAL AND METHODS Temperature Treatments Mojave Desert temperature data were obtained from a meteorological station maintained by NOAA and the Western Regional Climate Center at Lytle Ranch, UT (37°09’N, 114°01’W) for the past 30 years (www.wrcc.dri.edu/cgi-bin/cliMAIN.pl?ut5252). The mean monthly temperatures here for May-September included the temperature treatments from the present study (20-29°C) and there was a consistent 10°C difference between mean maximum and minimum values. A second weather station in St. George, UT (37°12’N, 113°57’W) maintained by the USGS from 1928-1996 (http://mojave.usgs.gov/climate-history/index.html) was also referenced for additional daily maximum and minimum temperature values. Temperature treatments were not chosen to reflect the estimated changes in average surface temperature due to climate change. Rather, treatments were selected to represent temperatures within the thermal neutral zone (TNZ, 29°C) and just below the TNZ (21°C) for N. Figure S1. Relationship between metabolic rate and ambient temperature of three desert woodrats (Neotoma lepida). The thermal neutral zone extends from approximately 25-35°C, as indicated by the flat black line. Line made with lowess smoothing function in R (www.r-project.org). lepida (figure S1). Metabolic rates increase when ambient temperatures are outside the TNZ as mammals must expend energy to maintain thermal homeostasis. Based on studies with Neotoma and laboratory rodents, we predicted that mammalian herbivores will be able to handle the highest dose of toxins at temperatures slightly lower than the lower critical temperature of the TNZ [2, references therein]. Differences between the metabolic rates of N. lepida at 21°C and 29°C are unlikely to explain the differences observed in our liver function assay. If metabolic rates governed liver function, then the highest capacity for detoxification should be observed at extremely low ambient temperatures where metabolic rates are also highest. However, data from laboratory rodents does not support this prediction; animals had reduced toxin tolerance at ambient temperatures outside the TNZ (8°C) where metabolic rates are highest [3]. Data from our preliminary studies on metabolic rates and clearance times of woodrats did not reveal a correlation between these two variables (figure S2). Figure S2. Preliminary relationship between metabolic rates and clearance times after a long, 30-day acclimation period at two ambient temperatures (blue = 21°C, red = 29°C) in Neotoma lepida. Each point represents a single individual. Clearance times are indicative of liver function; longer times equate to reduced liver function. Table S1. Ambient temperature (mean + 1 s.e.) of animal rooms (cool or warm) after two acclimation periods. maximum (°C) minimum (°C) long, 30-day acclimation short, 3.5-hour exposure cool warm cool warm 22.8 + 0.2 21.1 + 0.2 29.4 + 0.2 27.8 + 0.2 20.6 + 0.9 20.0 + 0.5 28.9 + 0.2 28.3 + 0.3 Measuring Metabolic Rates The procedure was followed as per [14]. Woodrats in the TNZ study were housed at room temperature (23-25°C) for at least one month and the chamber temperature ranged from 16-36°C during data collection. Woodrats used for metabolic rate and liver clearance time comparisons were housed at either 21°C or 29°C for at least three weeks and the chamber temperature was maintained at either 21°C or 29°C, matching the acclimation temperature of the animal. REFERENCES 2. Dearing M.D. 2013 Temperature-dependent toxicity in mammals with implications for herbivores: a review. Journal of Comparative Physiology B 183(1), 43-50. (doi:10.1007/s00360-012-0670-y). 3. Keplinger M.L., Lanier G.E., Deichmann W.B. 1959 Effects of environmental temperature on the acute toxicity of a number of compounds in rats. Toxicology 1(2), 156-161. 14. McLister J., Sorensen J., Dearing M. 2004 Effects of consumption of juniper (Juniperus monosperma) on cost of thermoregulation in the woodrats Neotoma albigula and Neotoma stephensi at different acclimation temperatures. Physiological and biochemical zoology : PBZ 77(2), 305-312. (doi:10.1086/380211).