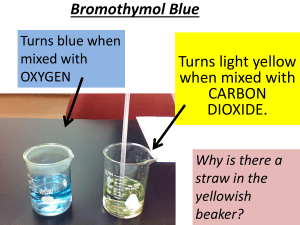
Carbon Dioxide Consumption in Drought-Tolerant vs. Non-Drought-Tolerant Plants Lyndsey Bennett Department of Biological Sciences Saddleback College Mission Viejo, CA 92692 Abstract Global warming is a growing concern for Earth, and rising levels of carbon dioxide in the atmosphere is a major contribution. With many parts of the country falling into a state of drought, individuals are replacing their gardens with plants that consume less water. This experiment is to prove non-drought-tolerant plants are more effective at consuming carbon dioxide from the environment than are drought-tolerant plants. Six carbon dioxide (CO2) probes were individually placed into airtight terrariums containing one of three drought-tolerant plants of the same species, and one of three non-drought-tolerant plants of the same species. Concentration readings of CO2 levels were recorded hourly, on three separate occasions. Over twenty-four hours, drought-tolerant plants consumed on average 361.1 ± 23.06 SEM parts per million (ppm) of CO2, while non-drought-tolerant plants consumed on average 804.78 ± 17.87 SEM ppm, proving to be significantly more efficient (p = 5.54 x 10-5). These results predict that levels of CO2 in the atmosphere will increase at a greater rate than currently occurs due to a lower abundance of efficient CO2 eliminating non-drought-tolerant plants. Introduction Two of the most critical concerns regarding today’s changing environment include the apprehension for potential drought in many regions around the world, as well as increasing levels of carbon dioxide (CO2) residing in the atmosphere and contributing to global warming (Beardsley, 2008). These issues go hand in hand, as limited water availability affects plants’ capacity to convert existing atmospheric carbon dioxide into oxygen through photosynthesis, thereby leaving higher amounts of CO2 to disrupt the Earth’s ozone layer (Beardsley, 2008). According to the National Centers for Environmental Information, nearly 18 percent of the United States was in a moderate to extreme drought at the end of October 2015. In response to this issue, an increasing number of individuals are replacing their typical, plush lawns and gardens with drought-tolerant plants, in an effort to conserve water. While these steps may result in lower water consumption, possibly alleviating immediate drought concerns, greater long-term damage in regards to raised CO2 levels in the atmosphere may result. Water is a vital component in photosynthesis (Osterhout and Haas, 1918). Less water, as during periods of drought, means less carbon dioxide uptake from the environment. The commonly accepted chemical reaction for photosynthesis shows the interaction between water and carbon dioxide: 6CO2 + 6H2O (+ light energy) → C6H12O6 + 6O2. This experiment was conducted to determine whether replacing non-droughttolerant plants with drought-tolerant plants is likely to result in a change of carbon dioxide levels in the atmosphere, thereby contributing to further and more rapid erosion of Earth’s ozone. It is expected the non-drought-tolerant plants are significantly better at converting carbon dioxide into oxygen through photosynthesis than are drought-tolerant plants. Materials and Methods Six carbon dioxide consuming plants were purchased from Lowe’s Hardware Store in San Clemente, CA; three of which were a drought-tolerant species of succulent Graptosedum, and three that were of the non-drought-tolerant species Epipremnum. Each of the plants was enclosed in an airtight terrarium made of a clear plastic Tupperware, then covered with clear saran wrap, and taped airtight. Each container was then positioned outside in direct sunlight. Using the Xplorer GLX Pasport, the carbon dioxide levels of each container was immediately analyzed upon their placement into the sunlight. For the next twenty-four hours the Xplorer GLX Pasport recorded carbon dioxide levels hourly to determine the change in carbon dioxide concentration. This experiment was conducted at the researcher’s house in San Clemente, CA on three occasions; 29 October, 2015, 2 November, 2015, and 5 November, 2015. A one-tailed, unpaired t-test was run to assess the mean intake of carbon dioxide from each species, to test the hypothesis that non-drought-tolerant plants consume more carbon dioxide than do drought-tolerant plants. Results An overall significant difference in the rate of carbon dioxide consumption was observed over a period of three trials between the drought-tolerant and the non-droughttolerant plants, with non-drought-tolerant plants proving to be more efficient at carbon dioxide consumption (p= 5.54 x 10-5) (Figure 4). Day one of recording carbon dioxide concentrations in the plants’ terrarium, nondrought-tolerant plants averaged a rate of consumption of 829.33 ± 55.35 SEM parts per million (ppm) of CO2. Drought-tolerant plants averaged a rate of 406.33 ± 39.55 SEM of CO2. All six plant environments reached the greatest level of CO2 around 2:00 in the morning, when plant respiration was complete. CO2 levels begin dropping at this point when all plants began to consume the CO2 for photosynthesis. 1200 Non-Drought-Tolerant 1 1000 Non-Drought-Tolerant 2 800 Non-Drought-Tolerant 3 600 Drought-Tolerant 1 400 Drought-Tolerant 2 Drought-Tolerant 3 200 2:00 Noon 10:00 8:00 6:00 4:00 2:00 Midnight 10:00 8:00 6:00 4:00 0 2:00 Concentration of Carbon Dioxide (ppm) Comparison of CO2 Uptake in Drought-Tolerant vs. NonDrought-Tolerant Plants Day 1 Time (Hour) Figure 1. Comparison of three drought-tolerant vs. three non-drought-tolerant plants over twenty-four hours. Non-drought tolerant plants are significantly better at consuming carbon dioxide (one-tailed, unpaired t-test; p = 2.05 x 10-4). Day two of recording carbon dioxide concentrations in the plants’ terrarium, nondrought-tolerant plants averaged a rate of consumption of 815. ± 65.09 SEM parts per million (ppm) of CO2. Drought-tolerant plants averaged a rate of 346.3 ± 13.48 SEM of CO2. All six plant environments reached the greatest level of CO2 around 2:00 in the morning, when plant respiration was complete. CO2 levels begin dropping at this point when all plants began to consume the CO2 for photosynthesis. COncentration of Carbon Dioxide (ppm) Comparison of CO2 Uptake in Drought-Tolerant vs. NonDrought-Tolerant Plants Day 2 1200 Non-Drought-Tolerant 1 1000 Non-Drought-Tolerant 2 Non-Drought-Tolerant 3 800 Drought-Tolerant 1 600 Drought-Tolerant 2 Drought-Tolerant 3 400 200 0 Time (Hour) Figure 2. Comparison of three drought-tolerant vs. three non-drought-tolerant plants over twenty-four hours. Non-drought tolerant plants are significantly better at consuming carbon dioxide (one-tailed, unpaired t-test; p = 1.07 x 10-3). Day three of recording carbon dioxide concentrations in the plants’ terrarium, non-drought-tolerant plants averaged a rate of consumption of 770. ± 31.48 SEM parts per million (ppm) of CO2. Drought-tolerant plants averaged a rate of 330.67 ± 25.51 SEM of CO2. All six plant environments reached the greatest level of CO2 around 2:00 in the morning, when plant respiration was complete. CO2 levels begin dropping at this point when all plants began to consume the CO2 for photosynthesis. Concentration of Carbon Dioxide (ppm) 1200 Comparison of Average CO2 Uptake in Drought-Tolerant vs. Non-Drought-Tolerant Plants Day 3 1000 Non-Drought-Tolerant 1 Non-Drought-Tolerant 2 Non-Drought-Tolerant 3 Drought-Tolerant 1 Drought-Tolerant 2 Drought-Tolerant 3 800 600 400 200 0 Time (Hour) Figure 3. Comparison of three drought-tolerant vs. three non-drought-tolerant plants over twenty-four hours. Non-drought tolerant plants are significantly better at consuming carbon dioxide (one-tailed, unpaired t-test; p = 2.05 x 10-3). Figure 4 compares the average of each group of plants for each of the three days shows there non-drought-tolerant plants are significantly better at absorbing CO2 from the environment (p = 5.54 x 10-5). On average, non-drought-tolerant plants consumed 804.78 ± 17.87 SEM ppm of CO2, while drought-tolerant plants consumed 361.1 ± 23.06 SEM ppm CO2. Concentration of Carbon Dioxide (ppm) Comparison of Average CO2 Uptake in Drought-Tolerant vs. Non-Drought-Tolerant Plants 1000 800 600 Non-Drought-Tolerant 400 Drought-Tolerant 200 0 1 2 Trial (Day) 3 Figure 4. Comparison of carbon dioxide uptake (ppm) in non-drought-tolerant plants vs. drought-tolerant plants. Non-drought-tolerant plants are significantly better at consuming carbon dioxide from the environment, averaging 804.78 ± 17.87 SEM ppm, while the drought-tolerant plants averaged a CO2 intake of 361.1 ± 23.06 SEM ppm (one-tailed, unpaired t-test; p = 5.54 x 10-5). Discussion Expectations that the drought-tolerant plants would be found less effective at absorbing carbon dioxide than the non-drought-tolerant plants were consistent with the results of the experiment, as shown in Figure 4 (p = 5.54 x 10-5). CO2 levels peaked in all terrariums around 2:00 in the morning (Fig. 1; Fig. 2; Fig. 3), after the plants completed respiration during periods of low light levels. The difference in each of the terrarium’s greatest level of CO2 concentration and lowest level of CO2 concentration shows the amount CO2 each plant is able to consume in parts per million (ppm) for photosynthesis. The differences in ability to consume CO2 are due to the varying about of stomata each type of plant holds. Nearly every terrestrial plant (one that grows on land) contains stoma, the pores through which carbon dioxide is consumed and converted to oxygen during photosynthesis, producing sugars necessary for plant survival and production (Kayvon, 2008). The commonly accepted equation for this chemical reaction is 6CO2 + 6H2O (+ light energy) → C6H12O6 + 6O2. Drought-tolerant and non-drought-tolerant plants have been found to contain vast differences in the number of stomata present on their leaves (Eckerson, 1920). These differences occur to genetic modifications made by the plants in response to the availability of water, a necessary component to partake in photosynthesis (Eckerson, 1920). Non-drought-tolerant plants respond to drought by altering their physical makeup, such as a reduction of leaf area, leaf wilting, as well as increased root growth and closure of stomata through the release of abscisic acid (ABA), a plant stress hormone (Kayvon, 2008). While these measures result in short term water use efficiency, there is also a reduction in carbon dioxide consumption for photosynthesis (Assmann, 2013). Drought-tolerant plants grow naturally in regions with limited water availability (Kayvon, 2008). These plants have evolutionarily modified their genetic makeup to improve their ability to survive in drought conditions (Assmann, 2013). Many develop leaves with a waxy coating to prevent water loss; others have lighter leaf color to reflect sunlight, or tiny hairs covering their exterior to reduce water transpiration (Assmann, 2013). Many of these plants also grow their roots laterally to increase water and nutrient uptake (Assmann, 2013). The greatest factor contributing to lower photosynthetic capability is the reduced number of stomata drought-tolerant plants develop (Edwards and Ogburn, 2012).. Stomata (stoma; plural) are the sites in plants where carbon dioxide is absorbed, and oxygen and water is released back into the environment (Edwards and Ogburn, 2012). Although the reduced carbon uptake of these drought-tolerant plants is the main focus of this experiment, the water cycle resulting from water released by the plant during the photosynthetic process is also an important factor. Lower quantities of water released from the plants leads to less moisture in the atmosphere for rainfall, leading to less moisture in the soil available for plant growth. When conditions are dry, stomata condense to prevent loss of water needed for survival (Lampard, et al., 2008). The smaller stoma in effect also reduces carbon consumption due to smaller surface area of the stoma opening for carbon consumption. According to the Unites States Environmental Protection Agency, carbon dioxide is the most emitted greenhouse gas due to human activity, claiming 82% of all the greenhouse gases individuals produce (NRC, 2010). Since the start of the industrial revolution, human contribution to carbon dioxide released emissions have risen drastically, roughly 80% between 1970 and 2004 (Wild, 2010). This is due primarily to fossil fuel use and land desiccation (Wild, 2010). When gases such as carbon dioxide enter the atmosphere, they are trapped in the environment and prevent escape of heat radiating towards space, a phenomena known as the “greenhouse effect” (Gardiner, 2004). Carbon dioxide is believed to contribute to climate change as it resides in the atmosphere and does not respond to fluctuations in temperature (Gardiner, 2004). This is where plants play a crucial role in the regulation of our environment, as they are needed to adjust the levels of carbon dioxide fixed in the atmosphere. An abundance of droughttolerant plants means lowered photosynthetic activity, resulting in higher levels of carbon dioxide in the atmosphere, decreased water availability, and fewer plants rich with stomata. As a plant is growing, environmental conditions affect “stomatal-development” pathways, which determines the number of stoma the plant will grow (Lampard et al., 2008). In an environment containing high levels of carbon dioxide and low levels of water, a plant will develop fewer stoma than it would in an environment rich in water availability and low in carbon dioxide (Edwards and Ogburn, 2012). Upon finding there to be a significant difference between drought-tolerant Graptosedum and non-drought-tolerant species Epipremnum in the consumption of carbon dioxide, further experiments should be conducted using a greater variety of drought-tolerant and non-drought-tolerant plants to obtain more comprehensive results. Studies should also be completed to find the total overall increasing rate of net CO2 in the atmosphere, after accounting drought levels and percentage of non-drought-tolerant plants being replaced with drought-tolerant plants. Literature Cited "Drought - October 2015." Drought. Web. 19 Nov. 2015. <https://www.ncdc.noaa.gov/sotc/drought/201510>. "US Environmental Protection Agency." EPA. Environmental Protection Agency. Web. 20 Nov. 2015. <http://www.epa.gov/>. Assmann, Sarah M. "Natural Variation in Abiotic Stress and Climate Change Responses in Arabidopsis : Implications for Twenty-First-Century Agriculture." International Journal of Plant Sciences (2013): 3-26. JStor. The University of Chicago Press. Web. 20 Nov. 2015. Beardsley, Timothy M. "An Epidemic with Global Consequences." Bioscience 58.6 (2008): 475. JStor. Oxford University Press on Behalf of the American Institute of Biological Sciences. Web. 20 Nov. 2015. Eckerson, Sophia H. “The Number and Size of the Stomata”. Botanical Gazette 46.3 (1908): 221–224. Web. 20 Nov. 2015. Edwards, Erika J., and R. Matthew Ogburn. "Angiosperm Responses to a Low-CO 2 World: CAM and C 4 Photosynthesis as Parallel Evolutionary Trajectories." International Journal of Plant Sciences (2012): 724-33. JStor. Web. 20 Nov. 2015. Gardiner, Stephen M. "Ethics and Global Climate Change." ETHICS Ethics 114.3 (2004): 555-600. JStor. Web. 20 Nov. 2015. Lampard, G. R., C. A. Macalister, and D. C. Bergmann. "Arabidopsis Stomatal Initiation Is Controlled by MAPK-Mediated Regulation of the BHLH SPEECHLESS." Science 322.5904 (2008): 1113-116. PubMed. Web. 20 Nov. 2015. NRC (2010). Advancing the Science of Climate Change. National Research Council. The National Academies Press, Washington, DC, USA. Osterhout, W. J. V., and A. R. C. Haas. "Dynamical Aspects of Photosynthesis." Proceedings of the National Academy of Sciences 4.4 (1918): 85-91. JStor. National Academy of Sciences. Web. 19 Nov. 2015. Wild, Martin, and Edgar Schmucki. "Assessment of Global Dimming and Brightening in IPCC-AR4/CMIP3 Models and ERA40." Clim Dyn Climate Dynamics (2010): 1671-688. Print.
 0
0
advertisement
Related documents







Download
advertisement
Add this document to collection(s)
You can add this document to your study collection(s)
Sign in Available only to authorized usersAdd this document to saved
You can add this document to your saved list
Sign in Available only to authorized users