Spatially structured food webs in a coloured environment Sara
advertisement

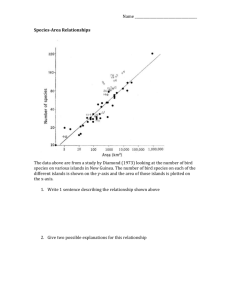
Spatially structured food webs in a coloured environment Sara Gudmundson, Frida Lögdberg and Uno Wennergren* Department of Theoretical Biology, Linköping University, Sweden, *Correspondence: Email: unwen@ifm.liu.se Introduction Species rich natural food webs contain complex interaction patterns evolved through historical processes in dynamic environments (May 1973). Food webs are known to endure unstable environments for several generations (Vasseur & Fox 2007). However, theoretical studies predict that large complex food webs should be unstable and extinction-prone because of high connectance, many modes of oscillation and positive feedback loops (May 1974; Tilman 1999; Green & Sadedin 2005; Borrvall & Ebenman 2008). The inability of explaining benefits of diversity implies that theory lacks important components regulating population dynamics. Furthermore, the stability analysis itself may not be sufficient. In this study, we add coloured environmental variation and spatial structure and investigate effects on the stability of food webs. In addition of using the most common stability measure, we perform a more thorough analysis concerning variation of densities. Food web stability is often measured as variability, which usually is calculated as the coefficient of variation, standard deviation divided by the mean (McCann 2000). Decreased variability implies decreased population variance which is likely to lower extinction risk (Lande 1993; McCann 2000). Addition of stabilizing mechanisms in models has been shown to facilitate food web complexity. For instance, network connectivity incorporating a mix of weak and strong links of interactions can inhibit oscillatory subsystems limiting species richness (Polis 1991; McCann et al. 1998). However, measurements of the coefficient of variation, CV, may not be enough for determining food webs able to withstand stresses. An increase in stability, measured as CV, can either imply an increase in mean density or a decrease in variance. A population consisting of just a few individuals can misleadingly be seen as robust to stresses as long as its variance is low in comparison to its mean. To address the risk of misinterpreting results of stability, we have evaluated mean and variance one by one in addition to variability measurements of food web biomass and species abundances. The environmental variance, measured in impact and frequency of extreme weather events, is increasing (Easterling et al. 2000). The change in climate is likely to cause increased variability and extinction risk of ecological systems (Lande 1993; Halley & Dempster 1996; Ripa & Lundberg 1996; Ripa & Lundberg 1996; Kaitala et al. 1997; Fontaine & Gonzalez 2005). When investigating the effect of environmental variation, it is important to consider different magnitudes of variance. Another important property of environmental variation is its correlation in time. Variation found in nature is considered to be the best represented by pink 1/f noise (Caswell & Cohen 1995; Halley 1996; Ripa & Lundberg 1996; Cuddington & Yodzis 1999). It describes correlations in many different scales and does not priorities between timescales of disturbances (Halley 1996). In order to investigate the effect of environmental variation on food webs, we incorporate 1/f noise with different magnitudes of variance and redness. Landscapes are known to hold different biotic and abiotic conditions giving rise to spatially separated subpopulations inhabiting patches. Dispersal between subpopulations enables reestablishment of extinct patches which can prolong the whole population’s time to extinction (Engen et al. 2002; Liebhold et al. 2004; Greenman & Benton 2005). In order to investigate stabilizing properties of dispersal between subpopulations, we position our food web in six spatially separated patches. All subpopulations are connected with dispersal. When modelling subpopulations in a variable environment, one also has to specify each subpopulations environmental response. Do all subpopulations respond the same to the environmental variation or do they have different responses? The answer will affect how subpopulation densities fluctuate in relation to each other. This property is called synchrony and can also be measured between species. Synchrony between species has been shown to have a substantial effect on food web stability and extinction risk. Asynchronous consumers coupled with uncorrelated environmental fluctuations can improve food web stability (1/CV) by dampening oscillations between resource and consumers (McCann et al. 1998; Vasseur & Fox 2007). High synchrony between species implies a lower species extinction risk than during asynchronous response (Borrvall & Ebenman 2008). We have also measured synchrony between species, according to (Vasseur & Fox 2007). In addition, we have measured the correlation between species and their environmental variation. By adding this measure we aim to increase the understanding of how common environmental fluctuations can affect species synchrony. Density regulation, environmental autocorrelation and dispersal are known to affect local population dynamics and should be included in investigations regarding population dynamics and extinction risks (Engen et al. 2002). We have used a food web withholding a mix of strong and weak links of interaction and investigated the effects of coloured environmental variation and dispersal between spatially separated subpopulations. We aim to increase the understanding of what enables species diversity in food webs. Our results show that coloured environmental variation can change relative abundance of species, increasing food web robustness by increasing the density of the species with the smallest population size in a constant environment. Stabilizing effects of dispersal between spatially separated subpopulations show the importance of spatial and temporal variability for food web resistance. Method The diamond shaped food web contains four species. Two consumers share one resource and have one common predator (Fig. 1). The dynamics are described by a continuous-time differential equation system, modelled by Vasseur & Fox (2007) after McCann et al. (1998). Resources grow logistically and consumers and predator have natural background mortality. Consumption is limited by a type II nonlinear functional response (Yodzis & Innes 1992; McCann et al. 1998; Vasseur & Fox 2007). The biologically plausible parameter values have previously been used by Vasseur & Fox (2007) and McCann et al. (1998) (Table 1). The values are estimated from studies on species’ body mass versus metabolic and ingestion rate (Dickie et al. 1987; Yodzis & Innes 1992; McCann et al. 1998; Vasseur & Fox 2007). Resource gain and predator preference are set higher for C1 than for C2. C1 is the strongest resource competitor and preferred prey of P. The competition irregularity causes intrinsic asynchronous fluctuations of consumers. Species densities fluctuate in stable limit cycles in constant environment. Figure 1 The diamond shaped food web with differential equation system (modeled after McCann et al. 1998). P is the density of the top predator, C1 first consumer, C2 second consumer, R the resource species and Ωi,j, represent the trophic interaction strength between the species. Table 1 Parameter explanation and their values. The standard deviation, σenv, colour, γ, and cross-correlation, ρenv, of environmental variation are independent parameters affecting the mortality rates of the consumers. Uncorrelated, white, environmental variation was generated from a random normal distribution with zero mean and σenv2 variance. Fourier transform was used to generate coloured 1/f noise. The discrete Fourier transform of the coloured noise, P(ƒ), was scaled according to: 𝑃(𝑓) = |𝑋(𝑓)|2 𝑓 −𝛾 (1) where ƒ is frequency, X(ƒ) is the discrete Fourier transform of the previously generated white noise and the colour of P(ƒ) was determined by the value of the spectral exponent, γ, where γ = 0 gives white and γ > 0 gives red noise. After colouring the time series, inverse Fourier transform was used on P(ƒ) to generate the coloured environmental noise, envi(t). The food web model was integrated across a range of σenv, 0 to 0.6 in steps of 0.05, and γ, 0 to 0.6 in steps of 0.2. Environmental variation affects the two consumers’ mortality rates through an exponential filter (Gillooly et al. 2001; Vasseur & Fox 2007): MCi (t) = MCi (0)eenvi (t) (2) where MCi(t) is the mortality rate at time t, MCi(0) is the medial mortality rate, envi(t) is the environmental variation at time t for consumer i. In order to determine the effect of dispersal between spatially separated subpopulations, all measurements in our study were taken from one patch of 6 in the landscape. Patches, containing the food web, were either isolated or connected with the other patches by dispersal. Dispersal between subpopulations was governed by a mass-action mixing process without distance dependence. Subpopulations with dispersal were connected through a dispersal matrix (Caswell 2001 and Wennergren et al. 1995) (Table 2). Table 2 Dispersal matrix with six patches. dij represents the proportion of the subpopulation in patch i that migrates to patch j in one time step. 0 d21 d31 d41 d51 d61 d12 0 d32 d42 d52 d62 d13 d23 0 d43 d53 d63 d14 d24 d34 0 d54 d64 d15 d25 d35 d45 0 d65 d16 d26 d36 d46 d56 0 Migrating proportions, dij, were generated from a random normal distribution with mean (patches)-1 and variance 0.2*(patches)-1. The distribution was truncated by 0 and 1.2*(patches)-1. The same dispersal matrix was used for all four species in this study. Cross-correlation of environmental variation, ρenv, had values -1, 0 and 1. -1 represented negative correlation between species and positive correlation between subpopulations within species. Two perfectly negatively correlated time series of environmental variation was generated for each species, all subpopulations within species was affected by the same time series. ρenv = 0 represented independent environmental response of all subpopulations. Independent time series of environmental variation was generated for all subpopulations, independent of species. Finally, ρenv = 1 represented positive environmental response correlation between all subpopulations. In this case all subpopulations were affected by the same time series of environmental variation. Simulations were made in Matlab 7.5.0 (R2007b, The Mathworks, Natick, MA, USA) with 100 replicates and 3000 time-steps. Initial subpopulation densities where chosen on the uniform interval; 0.1 to 1.0. Extinction risk was calculated as the risk of populations decreasing below the extinction boundary 10-6 and by how many replicates that had all subpopulations staying above the extinction boundary until the end of the simulation. With dispersal, populations were considered to decrease below the extinction boundary when the sum of all subpopulations within species decreased below 10-6. Replicates with extinctions were only analysed in respect to extinction risk. The first quarter of the simulated time series was excluded from analysis to avoid initial transients. Mean, variance and stability of patch density, species density and food web biomass, consumer synchrony and extinction risk were calculated for each of the combinations of varied parameters. Food web biomass was the sum of all subpopulations. Stability was measured as density variability: 1 𝜇𝑖 = 𝐶𝑉 𝜎𝑖 (3) where CV is the coefficient of variation, σi the standard deviation and μi the mean of population i’s density time series (Vasseur & Fox 2007). Consumer synchrony was calculated through: 𝜌𝐶 = 1 𝑁𝜎𝐶1 𝜎𝐶2 𝑁 ∑(𝐶1 (𝑡) − 𝜇𝐶1 ) (𝐶2 (𝑡) − 𝜇𝐶2 ) (4) 𝑡=1 where N is time series length, σi standard deviation and μi mean of consumer species i’s time series (Vasseur & Fox 2007). Consumer synchrony and environmental variation was calculated as equation (4), when ρenv =1, in order to evaluate the impact of environmental variation on each consumer. Results The magnitude of environmental variance was of great importance for food web stability and extinction risk. Low-to-moderate variance lowered variability of biomass and all species densities, except the resource, whereas higher variance destabilises the system (Fig. 2a, d, Fig. 3a). The standard deviation of environmental variation, σenv, generating maximum stability, was species specific. C1 and P gained their maximum stability from higher σenv than C2 and R. The same pattern was found for each value of cross-correlation of environmental variation, ρenv. Reddening of the environmental variation decreased the stabilising effect of low-tomoderate σenv and enhanced the destabilising effect of higher σenv. In addition, it lowered the σenv values generating maximum stability (Fig. 2d). Dispersal had minor affect during correlated environmental variation (Fig. 3). However, during uncorrelated environmental variation, the stabilising effect of low-to-moderate σenv was enhanced and the destabilising effect of higher σenv was reduced with dispersal (Fig. 2d, Fig. 3). Studies on time series of biomass and species abundances revealed that addition of dispersal between subpopulations resulted in maintenance of intrinsic dynamics during moderate σenv. The stable limit cycles where not as apparent in isolated patches during the same environmental variance (Fig. 4). Mean food web biomass decreased and biomass variance increased with increasing σ env (Fig. 2e, f), regardless of ρenv. However, a constant environment did not give the lowest variance in biomass. Low-to-moderate σenv actually resulted in a minor decrease in biomass variance. Measurements on time series of species densities showed that the value of σenv affected the relative abundance of species (Fig. 2b). Mean density of the species with the smallest population in constant environment, C1, increased with increased σenv (Fig. 2c). In contrast to C1, high σenv decreased mean density and resulted in a major increase in variance for the largest species in constant environment, C2. Mean density of R increased where as the mean of P decreased with increased σenv. Reddening of the environmental variation enhanced the effects of increased σenv on biomass (Fig. 2e, f) and each species. The same change in relative species abundance occurred, but for lower values of σenv. Dispersal coupled with uncorrelated environmental variation reduced the effects of increasing σenv on food web biomass (Fig. 2e, f) and species densities. Figure 2 Stability, mean and variance for species population densities and food web biomass with environmental fluctuation strength, σenv and uncorrelated environmental variation, ρenv=0. Left column; measurements on species population density with white environmental variation of γenv=0, without dispersal. P is predator, C1 first consumer, C2 second consumer and R resource. Right column; measurements on food web biomass with coloured environmental variation of γenv=0-0.6, without and with (crosshatch lines) dispersal. Figure 3 Stability of food web biomass with standard deviation of environmental variation, σenv and cross-correlation of environmental variation, ρenv. a) isolated patch b) patch connected by dispersal. Figure 4 System responses to continual synchronous point perturbations with standard deviation of environmental variation, σenv= 0.3 and uncorrelated environmental variation, ρenv=0. The patch that is connected by dispersal with the other patches maintains the intrinsic dynamics of the food web. Subpopulation extinction risk increased with increased σenv, regardless of the value of ρenv. ρenv = -1 gave the highest extinction risk whereas ρenv =1 gave the lowest. A similar pattern was found for each species, where C2 showed the highest sensitivity to increased σenv. Reddening of the environmental variation increased population extinction risk where as dispersal coupled with uncorrelated environmental variation reduced the risk of extinction. Both consumers become increasingly negatively correlated with their environmental variation during low-to moderate σenv. However, results differed for σenv values above 0.3. The negative correlation between C1 and the environmental variation continued to increase while the negative correlation between C2 and environmental variation started to decrease for higher σenv. Reddening of the environmental variation amplified the effect where as dispersal coupled with uncorrelated environmental variation decreased the effect of increased σenv. The pattern of differences in correlation was retained for all different scenarios tested. Consumer synchrony increased with increased σenv, regardless of ρenv. Reddening of the environmental variation enhanced the effect where as dispersal coupled with uncorrelated environmental variation reduced the synchronising effect of increased σenv. Discussion Natural food webs include complex interactions between species in different trophic levels and between each species and its environment. Low-to-moderate white environmental variation can increase food web stability (1/CV), by interrupting initial consumer asynchrony (Vasseur & Fox 2007). Our study confirms these results (Fig. 2a, d, Fig. 3). Consumer synchronisation caused a shift in total resource predation pressure affecting resource density. The shift in resource density caused another quick consumer response. Quick dynamic responses from consumers and resource dampened predator fluctuations and decreased biomass variance. For isolated patches, the lowest level of variability occurred during positive correlation of environmental variation, ρenv (Fig. 3a). Similar results have been found in a model where environmental variation affects the growth rates of all species in a food web (Borrvall & Ebenman 2008). Higher values of σenv decreased food web stability by increasing the variability of species densities. However, measuring food web stability by variability can be misleading without additional independent studies on mean and variance. The positive effect of low-to-moderate environmental variation can be questioned because of the resulting decrease in mean food web biomass and increased extinction risk. A decreased mean has negative effects on population persistence, such as increased effects of demographic stochasticity and catastrophes (Lande 1993). However, our independent studies of species mean densities reviled an environmental variation induced shift in relative abundance of species. It contributed to an increase of the species with the smallest population in a constant environment (Fig. 2b). The shift in relative abundance of species was caused by the intrinsic dynamics of the food web. It is important to have in mind that food web structure and choice of model parameter will affect the degree of sensitivity to different kinds of environmental variation (Greenman and Benton 2005). In our model, C2 has a lower ingestion rate than C1. This means that C1 has a better ability to take advantage of its resource than C2 during high environmental variance. Despite the increase in available resources, C1 was still affected by a high predation pressure from P limiting its density increase. Differences between the consumers were apparent in the correlation between each consumer and the environmental variation. The negative correlation between C1 and the environmental variation increased continuously with increased σenv where as the negative correlation between C2 and the environmental variation started to decrease after reaching a threshold value of environmental variance. P’s population decreased with increased σenv. Even though P prefers C1, it was negatively affected by the drastic density decrease of the originally large C2 population. A density increase of the smallest population, C1, will have positive effect on food web persistence. Food webs withholding species with large populations will have a smaller total risk of suffering from catastrophes and demographic stochasticity than food webs withholding small populations. In addition to measures of stability, we have investigated how different magnitudes of environmental variance affect the extinction risks in our model food web. As expected from earlier studies (Lande 1993, Engen et al. 2002), extinction risk for each species in the food web increased with increased σenv. Lowered mean densities and increased variance increased the risk of populations reaching extinction boundaries. C2:s poor resource tracking abilities gave C2 the highest extinction risk at high σenv, despite being the largest population in a constant environment. The result was caused by C2:s high density variance (Fig. 2c). Isolated subpopulations with uncorrelated and negatively correlated environmental variation had higher mean extinction risks than isolated subpopulations withholding a positively correlated environmental variation. Results may be explained by decreased variance in species densities with increased correlation (Borrvall & Ebenman 2008). Results of shifts in relative abundance with increased σenv indicate that addition of stress factors, such as catastrophes and demographic stochasticity, may affect the relationship between extinction risk and environmental variance. Moderate environmental variation may decrease the risk of extinction by increasing the density of the species with the lowest population in a constant environment. Further studies including these mechanisms may further clarify the effect of environmental variation and the importance of multiple measures when analysing food web stability and extinction risk. Environmental variation found in nature is considered to be positively correlated in time (Caswell & Cohen 1995; Halley 1996; Ripa & Lundberg 1996; Cuddington & Yodzis 1999). Red variation is dominated by low frequencies which result in bad/good conditions being retained for several time steps. Red environmental variation gives populations more time to respond to differences in their environment, increasing the probability of environmental fluctuation tracking (Ripa & Lundberg 1996). The stabilising power of low-to-mediate environmental variation was reduced and extinction risks where increased with increased redness. Maximum food web stability was found for lower σenv than during white environmental variation (Fig. 2d). Results are caused by reddened environmental variation causing larger time series variance differences than white variation (Greenman and Benton 2005). Cuddington and Yodzis (1999) have shown that reddening of variation can decrease mean persistence time in overcompensating single population models. However, our result showed that reddening of environmental variation amplified the shift in relative abundances of species, increasing the density of the species with the smallest population size. Reddening also increased consumer synchronisation which has been observed to reduce extinction risks in food webs without spatial structure (Borrvall & Ebenman 2008). Redness increasing the positive correlation between populations has also been shown in Greenman and Benton (2005). Reduced stabilizing effects and increased extinction risks caused by redness speak against the importance of environmental variation as an important stabilising property of food webs. However, addition of dispersal between subdivided populations clarifies the importance of spatial and temporal variability. Dispersal had a strong stabilising effect during uncorrelated environmental variation (Fig. 2d, Fig. 3, Fig. 4). Individuals from patches with good conditions were able to migrate to patches with poor conditions (Engen et al. 2002, Liebhold et al. 2004). The migration undermined consumer synchronisation and evened out destabilising effect of environmental variation. The equalising effect caused by dispersal had major implications for food web stability and extinction risks. The food web with dispersal affected by dark pink environmental variation was actually more stable than the food web in an isolated patch affected by white variation. Extinction risks were close to zero, during the interval of environmental variance and redness. However, higher σenv values generated similar destabilising effects of redness as in the case with isolated subpopulations. Kaitala et al. (1997) supports our results by showing that increased system complexity can reduce the effect of redness. Engen et al. (2002) showed that increasing dispersal between patches, withholding single species, results in longer time to extinction. Mass action mixing has no distance dependence. This infers similar probabilities of dispersal between all patches. The assumption can be far from dispersal found in nature. However, results from Petchey et al. (1997) showed minor differences in population persistence when comparing landscapes with global and local dispersal. Despite the lack of distance dependence, a minor increase in stability was observed in some patches when adding dispersal during correlated environmental variation. This effect can be explained by the variance in dispersal rate between patches caused by our dispersal matrixes generation method. The small variance in dispersal rates makes it possible for individuals in patches with larger dispersal rates to save other patches with lower dispersal rates at the cost of their original subpopulation density. Further studies on distance dependent dispersal withholding negative effects, such as additional death rates on dispersers, would further clarify the importance of dispersal between subdivided populations. Food web stability and extinction risk were measured at subpopulation level in this study. It is important to consider the differences between patch and landscape level when estimating food web resistance. The choice will have large effects on estimated extinction risks! When comparing species with and without dispersal during correlated environmental variation, the stability of the food web in the landscape without dispersal was much higher than the one with dispersal. Without dispersal, all subpopulations affected by weak variation, will fluctuate in their own phase, depending on initial densities. This asynchrony minimises the variance of the sum of all subpopulations which leads to larger landscape stability. With dispersal, patches that originally fluctuate in their own phase will eventually be more synchronised, preserving the variance in the sum of all subpopulations. Time lagged dispersal, more close to dispersal found in nature would decrease this synchronising effect. However, it is still important to think of these different scales both when investigating model food webs and when measuring populations empirically. The addition of coloured environmental variation and spatial structure had major implications for the stability and extinction risk of our model food web. Low-to-moderate coloured environmental variation had a stabilising effect by increasing the coefficient of stability and changing the relative abundance of species. However the stabilising effect was reduced with increased redness. An important implication of variation induced shifts in relative abundance is that present large population sizes may not give insurance towards future increase in environmental variance. Dispersal stabilised the system and lowered extinction risks during uncorrelated environmental response. Interaction pathways, exemplified in our study, have been shown to repeat at different resolutions, making food web stability scale invariant (McCann 2009). Our model can be seen as a building block for more complex food webs indicating that dispersal coupled by variability in space and time can be the missing component in theory explaining the resistance of diverse food webs.