Fish411 Research paper 145KB Dec 02 2013 08
advertisement

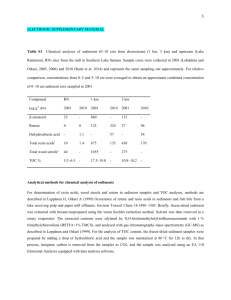
Introduction: Within recent years, the breadth and severity of ocean acidification (OA) has begun to be better understood. As carbon emissions continue to rise, the world’s oceans act as sink for the carbon dioxide. As these levels rise, the ecological and financial implications become more severe (Range et al., 2011; Kleypas et al., 2006). While OA is becoming much more well understood, literature on environmental effects on bivalve optics is limited. The current literature on scallop optics tends to concentrates on the biomechanics of the eye itself (see Land, 1964). Scallops are an ecologically and economically valuable family of organisms, both as prey species for other aquatic organisms as well as for scallop aquaculture. Despite the lack of genomic data on the scallop (family Pectinidae), current research is looking to illuminate the functions of the various phototransduction pigments and where they stand in genetic relatedness to each other (see Kojima et al., 1997; Pairett, 2013). We aim to look at the beginning of this cascade, specifically with the protein opsin, to attempt to draw out the implications of decreasing ocean pH on scallop vision. Experimental Procedures: We collected 43 specimens of pink scallops (Chlamys rubida) off the coast of Friday Harbor, San Juan Island, Washington. After the collection the specimens acclimatized in flow tanks for one week before being placed into experimental setup. Initially, a light/no light condition was set up for preliminary data. Then, using a 2X2 experimental design, four tanks were used for four separate conditions— normal pH/light, low pH/light, normal pH/no light, low pH/no light—the scallops were allowed to acclimatize to each condition for a period of 3 days. Tissues were extracted from the gill, mantle, and eye. It should be noted that due to the extraction methods used and the close adherence of eye to mantle, the eye samples contained small amounts of mantle tissue. Tissue samples were preserved at -80° C for processing. The frozen tissue samples were processed for RNA extraction using the protocol established for the University of Washington Fisheries 441 course. The tissue was homogenized with TriReagent® RNA isolation reagent to facilitate the precipitation of RNA. A sequence of ethanol rinses and centrifuge cycles produced a RNA sample from each tissue which were each quantified with NanoDrop®. After RNA isolation, the samples were processed for cDNA again using the University of Washington FISH 441 protocol. The preliminary test was run using qPCR using the primers set out by Serb et al. (2013). As discussed below, the qPCR was ineffective with the primers used therefore the primary samples were processed with conventional PCR using the protocol outlined by Serb et al. Reagents used and amounts include 12.5uL green master mix, 1uL primer, 2.5uL MgCl2, 5uL nuclease free water, and 3uL cDNA. Results: We did not see proper annealing of primers in either qPCR or our relaxed condition PCR. After RNA extraction from the whole tissue, the sample was quantified using NanoDrop® and all samples showed high concentrations and purity (averaging 450.7 ng/uL; 260nm/280nm: 1.89). The primers used were not species specific due to the lack of diversity in available scallop genomics. When no amplification was seen in the initial qPCR, we opted to use the less expensive PCR with a more relaxed temperature setting in an attempt to facilitate annealing. As Fig. 1 indicates, bands were seen at ~150bp and the negative control showed no bands, however we expected our gene product to show at ~550bp. Discussion: While the results proved less than desirable, they ultimately shed light on the current state of knowledge on OA and its effects on important oceanic species. Research continues to build the knowledge base on OA, however there yet remains many holes in our understanding of the full effects of OA. The primers used were not species specific thus as new genetic evidence becomes available it may reveal unique aspects of the scallop genome. With this new evidence, interesting aspects of the scallops’ ability for cope with environmental stressor may become known. In turn, these genetic indicators may reveal information on ways to better manage, from an anthropogenic perspective, the effects of OA on in-danger species. The margin for error within the complex processes that make up PCR is relatively high considering the approach we took to processing our samples. The primers we used were pulled from recently published literature and were not specific to the species we were testing. As more evidence mounts for genetic variability among species, the gene specific primers that become available may prove more effective. The protocol used for the PCR procedure followed that set out by Serb et al. however we used an available gotaq master mix which had a preset proportion of taq, taq buffer, and dNTPs. Therefore were unable to perfectly replicate the procedure as set out by Serb et al. in our experiment. Human error is certainly a consideration when looking at results like ours. While this is certainly a possibility, it is unlikely. All our samples showed the same result, i.e. bands at ~150bp, and the negative control showed no bands. This, as well as our NanoDrop results, lend to concluding there was little contamination in the samples. We need to spend more time on this project in order to elicit more concrete results. A larger sample size and more time to process those samples is required for any hope of significant findings. The appropriate primers and optimization sequence for PCR need to be teased out through experimentation to determine first whether opisn is present in our sample tissues, and then to determine the effects of environmental stressors on opsin levels. Once these conditions are solved then the bigger picture implications can be better understood. It would be advisable to look at the effects of OA on developing scallop larvae to further understand the implications on lowering ocean pH on bivalve organisms.