Mechanisms driving nonnative plant
advertisement

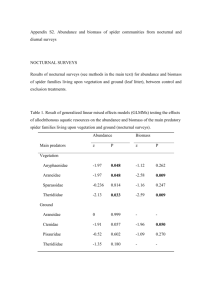
Mechanisms driving nonnative plant-mediated change in small mammal populations and communities Dan Bachen Master’s Student Department of Ecology, Montana State University And Andrea Litt Assistant Professor Department of Ecology, Montana State University Project duration: 2 years Total cost: 74,120 Project Summary/Abstract Nonnative plants can dramatically alter the habitat of native animals through changes in structure and composition of vegetation and the availability of food resources. Range expansion by the non-native grass: cheatgrass (Bromus tectorum L.) is a particularly acute threat for the sage-steppe ecosystem in Southwestern Montana. As climate is predicted to change over the next century, rangelands in Montana are predicted to become more hospitable to this invasive grass. Although previous research has documented changes in small mammal communities and populations concurrent with nonnative plant invasion, we know little about what mechanisms drive these changes. This project will seek to identify these mechanisms and their effects on three native species: deer mouse (Peromyscus maniculatus), sagebrush vole (Lemmiscus curtatus) and montane vole (Microtus montanus). Four primary changes are associated with cheatgrass invasion: increase in vegetation density in open spaces between shrubs, increase in litter, changes in available seed resource and changes in edible vegetation. I will first quantify these changes by analyzing the vegetation of an invaded area. Then I will develop treatments that mimic and isolate each of these factors. These treatments will be applied to randomly selected plots on the Gravely-Blacktail WMA. Using standard Capture Mark Recapture methods I will the estimate abundance and species diversity and compare these metrics to control plots to quantify the effect of each treatment. Data from these treatments will be combined several small studies looking at body condition and predator avoidance to suggest mechanisms driving changes in mammal abundance and community composition. Identifying how cheatgrass changes sage steppe and how these changes affect native species will provide critical information for managing for the conservation and persistence of some of Montana’s native small mammals. Bachen 2 Introduction: Nonnative plants can create considerable challenges for native animals by altering the structure and composition of vegetation. Small mammals are particularly sensitive to changes in vegetation structure and depend on plants and seeds for most, if not all, of their dietary needs. Many researchers have documented changes in community composition, population structure, and abundance of small mammals when an ecosystem is invaded by nonnative plants, such as spotted knapweed (Centaurea maculosa Lam.) and cheatgrass (Bromus tectorum L.). Range expansion by cheatgrass is a particularly acute threat for Southwestern Montana; as climate is predicted to change over the next century, rangelands in Montana are predicted to become more hospitable to this invasive grass (Bradley 2009). In the sage-steppe ecosystem, invasion by cheatgrass in combination with fire can lead to conversion from a shrub-land to annual grassland (Mack 1981). Succession of this nature is well-documented in the Great Basin where an estimated 17 million acres have been invaded (Kaufman 2007). Studies of small mammals have documented decreases in abundance associated with invasion; in one case, total abundance of small mammals in an invaded area was only 16% of that in a native sagebrush area (Ostoja 2009). Although previous research has documented changes in small mammal communities and populations with nonnative plant invasion, we know little about what mechanisms drive these changes. Some possibilities include: increases in litter cover and depth may make foraging more difficult, reductions in bare space between native vegetation and increased vegetation density may make movement more difficult, and changes in food resources (vegetation and seeds) may result in reduced reproductive activity. My study will seek to identify and isolate these mechanisms and quantify their effect on the community and populations of native species of small mammals. Bachen 3 This project will provide critical information for conservation and persistence of some of Montana’s native small mammals. Invasion by cheatgrass is correlated with disturbances, including grazing and fire, which are unlikely to decrease in the future. A disturbed ecosystem, in combination with the increasing vulnerability of Montana’s rangelands to invasion due to climate change, creates the very real potential that nonnative plants will increase in prevalence and drive concomitant changes in small mammal populations. These animals occupy basal positions in the food web and perform valuable ecosystem functions; therefore changes in their populations could affect numerous other species in the ecosystem. Understanding the impacts of nonnative plants on small mammals is therefore critical for management of the sage-steppe ecosystem. Literature Review Invasive species are now considered one of the primary causes of degradation of native landscapes (Vitousik 1996) and are considered a major cause of the extirpation and extinction of many species (Simberloff 2005, Young 2011). Climate change may exacerbate problems associated with the invasion of non-native species. A warming climate may change the ranges of exotic species (Bradley 2009, Walther2007), offer the opportunity for new introductions (Bradley 2009, Walther 2009), give nonnative species a competitive advantage (Fey 2011, Janion 2010), alter the ecology of native species (Schiffman 1994, Pearson 2008, Mack 1981) and increase the rate of invasion (Huang 2011). There is evidence that most of these species are not driving transformations of native communities but rather are symptoms of the larger climate change phenomenon (MacDougal 2005, Didham 2005). Cheatgrass is an invasive annuals grass found throughout the western United States (Young 1973). Introduced from Eurasia sometime in the nineteenth century, buy the 1980s it had become so prevalent that a paper published by Mack in 1981 described it as “the most ubiquitous and in many rangelands the dominate plant species” (Mack 1981). Knap estimated in 1996 that “Approximately 20% of the Bachen 4 Sagebrush-steppe (in the Great Basin Desert) vegetation zone is dominated to the point where establishment of native perennial species is nearly impossible” (Knap 1996). Fortunately in Montana establishment has not reached these levels, but the threat is projected to increase in the future (Figure 1, Bradley 2009) Cheatgrass is a passenger in Montana’s ecosystems, requiring disturbance such as grazing and road corridors to invade (Getz 2008). In the future we can expect to add climate change to this list of disturbances, opening up new areas for this invasive grass to propagate, establish and thrive. Alteration of Montana’s sage-steppe could be problematic for our native species of rodents due to their reliance on local vegetation for cover and food (Grant 1977). Cheatgrass has been documented to drastically change local vegetation and reduce native plant diversity (Reider 2010) which has an impact on the community structure and abundance of local rodents (Ostoja 2009). Experiments have shown that the increased density of vegetation associated with cheatgrass invasion has the potential to impede the movement of rodents and lizards possibility making them more venerable to predation (Newbold 2005, Reider 2009). Unfortunately other changes associated with cheatgrass invasion are not well documented. Approach: How does the invasion of nonnative plants into sage steppe ecosystems alter its small mammal community structure and species abundance? My primary question is relatively complex so to adequately address it I created a sequence of questions to address in a logical manner that will identify the mechanisms by which invasive plants influence local mammal communities. Question 1: Does cheatgrass alter the structure of sage-steppe vegetation and the quality and quantity food resources within the ecosystem? If so what are the magnitude of its effects? That cheatgrass has the potential to change vegetative communities and structure is addressed in the literature (see review) Bachen 5 but information quantifying these changes is lacking. To address this lack of information I will implement a small study to determine the increase in litter, increase in vegetation density between shrubs, change in seed resource and change in vegetation resource. Question 2: Is there evidence that small mammal communities and species abundance are influenced by the structure of their habitat’s vegetation and by the available food resources? This has already been established in the literature through studies showing correlation between changes in the local environment and subsequent changes in the local mammals. Question 3: Can native plant communities be manipulated in such a way that the alterations mimic those induced by the presence of non-native plants? Answering this question will require building on the knowledge gained in question 1 to develop treatments that mimic and isolate one of the for factors. This requires creativity to think of a way to simulate isolated factors in a way that ensures that the factor focused on is the only difference between treatment and control. Once developed, these treatments will be tested for their ability to mimic aspects of cheatgrass by applying them to 10 meter square study plots and comparing them to aspects of a real invasion. In this way I will be able to check and make alterations until I have developed an accurate way to isolate and mimic certain effects of cheatgrass invasion (Figures: 7 development of treatments,8 adjustment of control, 9 summary of treatments). Question 4: Can the consequences of manipulating the physiognomy of the vegetation and the available food resources be quantified in small mammal community structure and species abundance? If so what are the effects? I will answer this by applying the treatments developed in question 3 to randomly selected study plots within the sage steppe ecological system on the WMA. All treatments will be paired with a control plot and replicated. For each plot I will estimate the abundance of each species and the community structure. An analysis of the differences of these metrics will be made for each treatment and its control. This will determine the effect of each factor. Bachen 6 Question 5: Is there a plausible explanation for how the treatments affected (or didn’t affect) the community structure and species abundance of the small mammals? Answering this will allow me to more confidently attribute changes in abundance and community structure to the altered habitat factors. Adequately addressing each response will require identification of a mechanism driving differences between treatment and control. Information to determine these mechanisms may be found in the literature or require further small studies (see figure 10). If a decrease in abundance for one of the species is seen with increased density of faux grass treatment in the interspace, it may be driven by reduced maximum sprint speed and therefore reduced ability to effectively escape predation. Decrease could also be driven by reduced ability to forage effectively for food (seeds and or arthropods). Two tests should provide insight: Maximum sprint speed trials in native vegetation and faux grass vegetation and body condition as a function of average weight by age class. If abundance decrease is driven by predation I would expect to find a mean reduction in sprint speed, but no difference in body condition. If the decrease is driven by a reduction in foraging ability, I would expect to see a decrease in body condition but relatively similar sprint speeds. If tests confirm a reduction in body condition and sprint speed a combination of both factors may be occurring. A decrease in abundance seen with the litter treatments may be due to the same mechanisms as with an increase in vegetation density in interspace. The protocol for testing each mechanism would be the same as stated above, body condition and sprint trials. Likewise the hypothesis would be that litter impedes ability to forage and litter impedes ability to escape from predators. Population fluctuation due to addition of potentially edible grass and reduction of native forbs could be explained through simple mechanisms. An increases in abundance would suggest that animals are consuming the grass and it is good forage and compensating for reduction in native forbs. This will be tested by comparing body condition of animals caught during CMR trapping between plots as well as Bachen 7 determining palatability of grass though laboratory trials comparing body condition of animals given a native diet and comparing them to an invasive diet. If body condition is found to be similar or increased in animals fed on the invasive diet this would support cheatgrass being a high-quality food source even at the detriment of native forbs. If a decrease in abundance is seen between treatment and control plots on the WMA and is concurrent with decreased body condition in the field and in the laboratory this would suggest the cheatgrass does not provide high-quality forage which does not compensate for the reduction in native forbs. If no change is seen this would suggest that cheatgrass compensates for loss of forbs but does not increase food resource. Finally differences in abundance in the seed treatment could be explained in much the same way as the edible vegetation and tested in a similar manner. Increase in abundance for a species would indicate cheatgrass seed is a high-quality food resource, this would be confirmed by looking at body condition in the field and laboratory. If addition of seed produces no significant change in abundance, than cheatgrass seed is either implantable or a poor food resource. This would also be confirmed by body condition analysis. If all questions are systematically answered than a causative link should be able to be drawn from the alterations of vegetative structure and food resources caused by cheatgrass to the effects have on small mammal communities and species abundance, and the mechanisms driving these changes inferred. Measurement Methods Vegetative sampling will be conducted using two processes to measure both vertical and horizontal structure. To quantify vertical structure I will use the SideLook Program (Zehm 2003). This program analyzes profile pictures of .3 square meters of vegetation taken against a solid background. The pixels containing plant materials are quantified in various ways by row and column allowing analysis of features like average vegetation height and average amount of space between vegetation at ground Bachen 8 level. Using this program almost completely removes of observer bias and allows a more sophisticated analysis of structure. To assess the percent cover of vegetation (horizontal structure) of my study plots I will sample 1 meter square quadrats (Daubenmire 1959). Each quadrat will be assessed as to the percent cover of bare ground, low bryophyte, forbs, grass, invasive plants and shrubs. I will quantify edible vegetation and seeds as concentration of biomass per hectare. The percent cover estimates will be combined with clipping and weighing of vegetation to produce estimates of the biomass of edible vegetation available to small mammals on the study plots. Seeds will be counted in small random sub-plots and the densities per meter squared extrapolated to the entire plot. Mammal abundance and community composition will be assessed by trapping using Sherman live traps in 10 by 10 grids on each plot over 5 nights. Captured mammals will be measured, aged and sexed to gather valuable data on demographics and body condition. Individuals will be weighed with a pezo scale accurate to .1 grams. Sex will be determined visually in hand. Breeding condition will also be determined in a similar manner, examining individuals for evidence of swollen testicles (males) and swollen nipples or distended abdomen (females). Age will be determined by a combination of morphological cues (pillage, head: body ration) and weight. Upon capture individuals will be uniquely marked with double ear-tags for species with large enough ears to hold a tag (deer mice) or sharpie marker for those with ears to small (voles). Laboratory body condition analysis will be conducted in accordance with established Montana State University Animal Care and Use Committee guidelines. Animal housing and feeding will likewise conform to these guidelines. As deer mice are possible carriers of the Sin Nombre virus which causes Hantavirus pulmonary syndrome I will obtain laboratory animals from the Peromyscus Genetic Stock Center. Both species of voles are not carriers of this disease so I will obtain individuals from the local area. Randomly selected individuals will receive either of two treatment diets (cheatgrass seed or cheatgrass vegetation Bachen 9 with a realistic degree of supplemental food) or a standard laboratory diet. All will be measured on a daily basis prior to feeding using a pezo scale. A sample size of 15 individuals will be used in each category. Trials will be conducted over 1 month. If body condition for any individual decreases to critical levels it will be removed from the experiment and fed on a standard laboratory diet. Statistical Methods The analysis used in this project will focus on quantifying the differences or treatment effects between plots treated as discussed previously and controls. For all cases I will hypothesize that there is no difference or treatment effect, with the alternative hypothesis being that there is a treatment effect. For each comparison I will find a difference in means and the amount of variance for this parameter. Using the program R, I will run a Bootstrap analysis and generate 95% confidence intervals for the treatment effect or difference. This will allow me to see if there is a possibility of no effect (overlaps zero) and quantify the difference or effect. To quantify differences in vegetative structure and available food resources between treatment and control plots, output from program SideLook such as density by column and row, height, roughness, average hole size and others will be compared. The percent cover for each of the cover classes will also be compared. The analysis for the food resources will all be similar. Each resource will be measured, converted to a density for each plot then using an average weight for the unit converted to biomass (kg/ha). Analysis of abundance and species richness will be performed for the mammals captured. Abundance estimates will be made using program MARK for each species on each plot. There is the possibility that the amount of captures will be low enough to present problems accurately estimating species abundance. If this is the case I will aggregate the data to achieve an adequate sample size (Litt and Bachen 10 Steidel 2010). If enough captures are generated, species estimates will be done by age class, yielding a more accurate analysis of effect. Diversity will be considered the as the number of species caught by plot and averaged by plot by treatment. Data for demography of the populations of mammals on my study plots will be collected by assessing age class, sex and breeding status for all captures. This data will be adjusted for the possibility of unequal probability of detection. Calculation of this statistic will be done in conjunction with the analysis for abundance in MARK. Adequacy of Design Accurate estimation of the sources of variation in the estimated abundance of the plots can be used to prospectively test whether the proposed design is adequate and if not, optimize it. My objective in this analysis is to determine if my methods will be able to discriminate a biologically significant treatment effect (>2-3 animals) given the relatively small number of plots used. In this study there are two primary sources of variation that in addition to treatment effect cause differences in the number of animals estimated to be on each plot. The first source of variation is found in the accuracy of the estimates using the CMR method and its analysis. The second is the natural variation in animal numbers between plots; this is a function of naturally occurring differences in habitat and food resources due to each plot being relatively unique. Both of these sources of variation can confound the ability to discriminate a treatment effect. Variation in the estimation method is possible to estimate using a combination of the analysis software and R statistical package to run analysis of mock data to check accuracy. The natural variation between plots is next to impossible to estimate without pilot study data, which we do not have. Similar to natural variation between plots, treatment effect is hard to estimate without a pilot study, although other Bachen 11 research has found the holistic effect of cheatgrass invasion reduces abundance by up to 84% (Ostoja 2009). The primary driver of accuracy in estimating abundance when using analysis software such as MARK is the probability of capture per individual per night. As this set of parameters is unknown and can change over time I estimated the accuracy of estimates for three different models showing different types of trap response and also tested these models with a range of probabilities (Figure 4). I found an acceptable degree of variation for most probabilities with a homogeneous response. Exploring heterogeneity of response I found that a moderate degree of trap attraction or trap avoidance produced wider confidence intervals. This analysis elucidated the need for an adequate amount of traps to optimize the capture probability. As the animals themselves dictate the degree of heterogeneity in trap response there is little I can do to influence this to increase accuracy. As stated in the first paragraph estimation of the natural variation in abundance by plot is impossible without pilot study data. This is problematic as this variation could mask a treatment effect. Fortunately other research on small mammals has been conducted in a similar environment with similar methods that I can use not to estimate the variation due to differences in abundance by plot but to estimate this variation in combination with the variation due to analysis method. The study that provides this useful information was done by Maron in the Blackfoot river valley in western Montana (Maron 2010). The local environment is listed as native grassland with the dominant shrub being big sage brush (Artemisia tridentate), this is descriptive of a sage-steppe and is relatively similar in structure to my study area (personal observation). The study explores predation effects on small mammal populations, but uses very similar methods to those I am proposing. 4 control plots which are 1 Ha in size are trapped with a grid of 100 Sherman traps over 5 nights. Data were collected three Bachen 12 times a year between 2003 and 2008 (additionally once in 2002 and twice in 2009). The paper contains a table with the summary of estimations of abundance and the estimated SE (Figure 3). I analyzed his results for both P. maniculatus and M. montanus. It appeared that there were 3 distinct levels of abundance for both species (low: mean 1-2; medium: mean 3-6; large: 7-17). Small mammal populations are often cyclic in nature so this variation is not surprising. Interestingly variation increases with increase in estimated abundance. The variation found at low and medium levels would be acceptable to discriminate a biologically relevant difference in populations (>3 individuals/HA), the amount of variation found at high levels could potentially mask a relevant treatment effect. I believe there are two possible explanations for this phenomenon. The first is that capture probability of individuals decreases with an increase in populations as traps in optimum locations are filled. This seems unlikely because the ratio for highest population estimated to traps used was no more than 1individual to 4 traps. If this is the source of variation increasing density of traps may be a way to counter this source of variation. The second possibility is that an overall increase in abundance is not manifested evenly across the landscape and the variation is inherent in the samples. I can do nothing to correct for this. It is difficult to identify whether estimation error or plot difference drive the variation in this data. Without knowing which model was used to generate the estimates I don’t know whether heterogeneity of trap effect was present. Also I can only guess at the probability of capture. If I wanted to compare all possible variation with variation seen I would have possible combinations and no way to discriminate which was correct. To summarize this analysis my proposed methods will produce adequate results. Through analysis of the CMR method it is clear that in almost all cases estimates at all capture probabilities are adequate but the accuracy increases with capture probability. This would support using as many traps as possible to Bachen 13 ensure the maximum capture probability possible. The optimal number of plots is similar to this the more I use, to some degree the more accurate I will be in my estimations of treatment effect. Unfortunately since I am conducting this experiment in the real world the optimal number of plots to use is not the maximum due to logistical constraints. I could not possibly allocate my resources to do more than 8 plots per treatment and 8 per control for each treatment. Ideally I would just do 4 plots for each. The analysis showed that in all cases there was little practical difference between the ability to discriminate a difference of 2 individuals at low, medium or high between 4, 6 or 8 plots. In most cases 4 plots did about as well as 6 or 8 at distinguishing a difference of 1 except at medium levels of abundance where going to at least 6 plots would double the percent chance of determining a statistically significant difference (Fig 4). Based on the analysis I will increase my number of plots to 6. Timetable See Attached (Figure 5) Budget See Attached (Figure 6) Qualifications See Attached (Figure 11) Bachen 14 Literature Cited Blank, R. R., T. Morgan and F. Allen. 2011. Influence of plant invasion on seed chemistry of winterfat, green rabbitbrush, freckled milkvetch, indian ricegrass and cheatgrass. Natural Resources and Environmental Issues 16: article 19, 1-3. Bradley, B. A., M. Oppenhimer, and D. S. Wilcove. 2009. Climate change and plant invasions: restoration opportunities ahead? Global Change Biology 15: 1511-1521. Daubenmire R. 1959. A Canopy-coverage method of vegetation analysis. Northwest Science 33(1): 4364. Didham, R.K., J.M. Tylianakis, M.A. Hutchison, R.M. Ewers and N.J. Gemmell. 2005. Are invasive species the drivers of ecological change? Trends in Ecology and Evolution 20(9):470-474. Fey, S.B., and K.L. Cottingham. 2011. Linking biotic interactions and climate change to the success of exotic Daphnia lumholtzi. Freshwater Biology 56: 2196-2209. Getz, H.L. and Baker, W.L. 2007. Initial Invasion of cheatgrass (Bromus tectorum) into burned pinionjuniper woodlands in western Colorado. American Midland Naturalist 159:489-497 Grant, W.E., N.R. French and D.M. Swift. 1977. Response of a small mammal community to water and nitrogen treatments in a shortgrass prairie ecosystem. Journal of Mammalogy 58(4): 637-652. Huang , D., R. A. Haack and R. Zhang. 2011. Does global warming increase establishment rates of invasive alien species? A centurial time series analysis. PLoS ONE 6(9), e24733. doi:10.1371/journal.pone.0024733 Janion, C. H.P. Leinaas, J.S. Terblanche and S.L. Chown. 2010. Trait means and reaction norms: the consequences of climate change/invasion interactions ant the organism level. Evol. Ecol. 24:1365-1380. Kaufman, S. R., and W. Kaufman. 2007. Cheatgrass. Pages 191-192 in Invasive plants: a guide to identification and the impacts and control of common North American species. Stackpole Books, Mechanicsburg, PA, USA. Knapp, P. A. 1996. Cheatgrass (Bromus tectorum L.) dominance in the Great Basin Desert. Global Environmental Change 6(1):37-52. Bachen 15 Litt, A.R., and R.J. Steidl. 2009. Improving estimates of abundance by aggregating sparse capture-recapture data. Journal of Agriculture, Biological and Environmental Statistics 15(2): 228-247 MacDougal, A.S., and R. Turkington. 2005. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 86(1): 42-55. Mack, R. N. 1981. Invasion of Bromus Tectorum L. into western North America: an ecological chronicle. Agro-Ecosystems 7: 145-165. Maron, J.L., D.E. Pearson and R.J.Fletcher. Counterintuitive effects of large-scale predator removal on a midlatitude rodent community. Ecology 91(12):3719-3728. Newbold, T.A.S. 2005. Desert horned lizard (Phrynosoma platyrhinos) locomotor performance: the influence of cheatgrass (Bromus Tectorum). The Southwestern Naturalist 50(1):17-23. Nobis, M. 2005. SideLook 1.1 - Imaging software for the analysis of vegetation structure with true-colour photographs; http://www.appleco.ch. Ostoja, S. M., and E. W. Schupp. 2009. Conversion of sagebrush shrublands to exotic annual grasslands negatively impacts small mammal communities. Diversity and Distributions 25: 863-870 Pearson, D.E. and R.J. Fletcher. 2008. Mitigating exotic impacts: restoring deer mouse populations elevated by an exotic food subsidy. Ecological Applications 18 (2): 321-334. Reider, J.P., T. A. S. Newbold and S. M. Ostoja. 2010. Structural changes in vegetation coincident with annual grass invasion negatively impacts sprint velocity of small vertebrates. Biological Invasions 12: 2429-2439. Simberloff, D. 2005. Non-native species do threaten the natural environment!. Journal of Agricultural and Environmental Ethics 18:595-607. Schiffman, P.M. 1994. Promotion of exotic weed establishment by endangered giant kangaroo rats (Dipodomys ingens) in a California grassland. Biodiversity and Conservation 3:524-537. Vitousik, P.M., C.M D’antonio, L.L. Loope, and R. Westbrooks. 1996. Biological Invasions as Global Environmental Change. American Scientist 84(5): 468-478. Walther, G.R., E.S. Gritti, S. Berger, T. Hickler, Z. Tang and M.T. Sykes. 2007. Palms tracking climate change. Global Ecol. Biogeogr. 16: 801-809. Bachen 16 Young, J.A. and R.A. Evans.1973. Downy Brome: Intruder in the plant succession of big sagebrush communities in the Great Basin. Journal of Range Management 26(6): 410-415. Young, A.M., and B.M.H. Larson. 2011. Clarifying debates in invasion biology: a survey of invasion biologists. Environmental Research 111:893-898. Zehm, A., M. Nobis and A. Schwabe. 2003. Multiparameter analysis of vertical vegetation structure based on digital image processing. Flora 198(2): 142-160. Bachen 17 Appendix Figure 1. Map from Bradley et al. 2009 depicting increased threat of invasion of cheatgrass due to a changing climate. Map on right highlights location of Gravelly-Blacktail WMA. Figure 2. Estimates of abundance of deer mice and montane voles from Maron et al. 2010. Bachen 18 Figure 3. Three graphs illistrating the possible variation in abundance estimates with changing capture probability. The first illistrates even probability across 5 nights, the second simulates trap attraction where probability of capture increases each night as animals prefertually returen to traps. The third simulates trap aversion where animals are less likely to return to traps after capture. Hom ogenious Trap Response Difference from true N 30 25 20 15 10 5 0 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 Probability of detection Trap Aversion (probability decreasing .1/night) 16 Diffence from true N 14 12 10 8 6 4 2 0 0.4 0.5 0.6 0.7 0.8 0.9 1 Starting probability Bachen 19 Figure 4. These graphics show the ability to correctly detect a treatment effect as a function of the variation for low , medium and high densities based on Maron 2010 and number of plots per treatment at increasing treatment effects. The X axis is treatment effect in number of animals, the Y axis is the proportion of times the treatment effect is detected. Note the variation found at medium densities and a treatment effect of 1. Bachen 20 Figure 5. Timeline of project Timing 2011 2012 Activity Fall Begin vegetation surveys to quantify differences between invaded and non-invaded sites Winter Develop any specialized equipment needed for treatments Spring Continue surveys to quantify vegetation differences Begin treatments Summer/Fall Continue surveys to quantify vegetation differences Small mammal trapping 2013 Spring Continue treatments Summer Small mammal trapping Fall Complete thesis and publish findings Figure 6. Total Project Budget Funding Source Item Cost Montana State University Salary $60,000 Traps In-kind Gas 3,000 Trapping supplies 1,000 Lab animals 625 Animal equipment 400 Various small grants/MSU Technician salary MT Fish, Wildlife and Parks Field housing In-kind Vehicle In-kind Field assistance In-kind Plant mimics Sterilized cheatgrass seed 700 0 Hay pellets 150 Straw 120 Total Cost Bachen 8,325 $74,120 21 Figure 7. This flow chart shows my process for developing treatment that mimic factors of interest, and ensure that the treatments create conditions similar to those found in invaded ecosystems. This example uses litter as the factor of interest. Figure 8. This illustrates the adjustment of the control to isolate one factor in a case where the treatment causes a difference between two factors. Bachen 22 Figure 9. Chart displays the process of implementation for 4 treatments starting with quantifying the factor on an invaded plot. Action/Treatment Litter Interspace Seed Edible Vegetation Quantify On Invaded Plot Measure litter depth and density Measure density of cheatgrass in interspace Measure seeds Measure vegetation density, convert to Kcal/ha Develop Treatments Apply straw Produce and apply monofilament grass plant replicas Addition of unviable seed Herbicide to reduce forbs, apply straw pellets Adjust Control Add supplemental feed none needed none needed none needed Implement Implement Implement Implement Bachen 23 Figure 10. Potential mechanisms that could cause treatment effect and additional experiments performed to support each mechanism Potential Causal Mechanism mechanisms Litter Interspace Seed Edible vegetation - Increased difficulty of foraging - Impediment of movement leading to increase in predation -Cheatgrass seeds provide better/ worse quality of forage - Cheatgrass provide better/ worse quality of forage -Movement trials comparing sprint speed in fake grass and native environment -Diet preference study - Diet preference study - Body condition study on treatment and control plots -Body condition - Demographic analysis - Demographic analysis - Impediment of movement leading to increase in predation Additional - Body condition study experiments on treatment and to support control plots mechanisms - Demographic analysis -Movement trials comparing sprint speed in litter and native Environment study on treatment and control plots Figure 11. Relevant coursework: STAT 511, 512 Methods for Data Analysis (2 semesters) Statistical Sampling (1 semester) Research Methods (1 semester) Wildlife Habitat Relationships(1 semester) Applied Population Ecology (1 semester) Human Dimensions in Fish and Wildlife Management (1 semester) Community Ecology (1 semester) Relevant work experience: Research Assistant, Montana State University Aug 2011 – Present Design and conduct experiments to identify causes of changes in small mammal communities invaded by nonnative plants Bachen 24 Field Zoologist/Research Assistant: Montana Natural Heritage Program May-Nov 2008, May 2009-Aug 2011 Senior crewmember on two-person team conducting diversity monitoring surveys for reptiles, amphibians, and small mammals, including bats Worked unsupervised in remote locations across Montana, following and interpreting protocols as necessary to collect and manage data Responsible for entering, correcting and error checking data from the project and for processing other types of data for the Heritage Program Data Processing Technician, United States Forest Service Sep-Dec 2010 Worked as an independent contractor and processed records of nesting raptors collected over a 20 year-period in southeastern Montana, formatted information to append to the Montana Natural Heritage Program Point Observation Database Bachen 25