Electronic Supplementary Information 1b: The functional cartography
advertisement

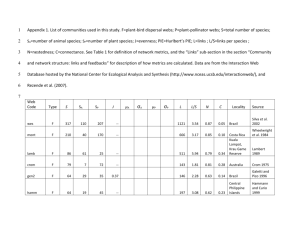
Electronic Supplementary Information 1a: Summarized description of the dataset Study system ID Fig. 1 1 2 3 4 5 6 7 1 Alien plant species Impatiens glandulifera Carpobrotus affine acinaciformis Multiple species Multiples species Opuntia sp. Opuntia sp. Impatiens glandulifera Geographic region Central Germany Northeastern Spain (Mainland) Argentina (Southern Andes) Mauritius Northeastern Spain (Mainland) Spain (Islands of Menorca & Tenerife) Southern England Habitat type No. of networks Publication Riparian 2 1 Mediterranean shrubland 6 2 Forest 8 3,4 Heathland 2 5,6 Mediterranean shrubland 6 1 Rocky coast, shrubland 8 7 Ruderal 8 8 Bartomeus, I., Vilà, M. & Steffan-Dewenter, I. (2010). Combined effects of Impatiens glandulifera invasion and landscape structure on native plant pollination. J. Ecol., 98, 440–450. 2 Bartomeus, I., Vilà, M. & Santamaría, L. (2008). Contrasting effects of invasive plants in plant-pollinator networks. Oecologia, 155, 761–770. 3 Morales, C.L. & Aizen, M.A. (2006). Invasive mutualisms and the structure of plant–pollinator interactions in the temperate forests of north-west Patagonia, Argentina. J. Ecol., 94, 171–180. 4 Aizen, M.A., Morales, C.L. & Morales, J.M. (2008). Invasive mutualists erode native pollination webs. PLoS Biol., 6, 396–403. 5 Kaiser-Bunbury, C.N., Memmott, J. & Müller, C.B. (2009). Community structure of pollination webs of Mauritian heathland habitats. Perspect. Plant Ecol. Evol. Syst., 11, 241–254. 6 Kaiser-Bunbury, C.N., Muff, S., Memmott, J., Müller, C.B. & Caflisch, A. (2010). The robustness of pollination networks to the loss of species and interactions: a quantitative approach incorporating pollinator behaviour. Ecol. Lett., 13, 442–452. 7 Padrón, B., Traveset, A., Biedenweg, T., Diáz, D., Nogales, M. & Olesen, J.M. (2009). Impact of alien plant invaders on pollination networks from oceanic and continental islands. PLoS ONE, 4(7), e6275. 8 Lopezaraiza-Mikel, M.E., Hayes, R.B., Whalley, M.R. & Memmott, J. (2007). The impact of an alien plant on a native plant–pollinator network: An experimental approach. Ecol. Lett., 10, 539–550. Electronic Supplementary Information 1b: The functional cartography methodology For the calculation of modularity M and the number of modules we used the method of functional cartography (software NETCARTO) based on simulated annealing (Guimerà & Amaral 2005a). Simulated annealing is a stochastic optimization technique that combines multivariate optimization and statistical mechanics to maximize M (see Kirkpatrick et al. 1983; Guimerà & Amaral 2005a,b; Olesen et al. 2007 for further details). For each network, NETCARTO calculates an index of modularity (M): I s ks 2 M 2I s1 I NM where Nm is the number of modules, Is is the number of links between species within module s, I is the number of links in the network, and ks is the number of links of all species in s. Hence, M increases with increasing link density within modules and decreasing connectedness between different modules. The number of modules of a network represents the optimal number of partitions when M is maximized (Guimerà & Amaral 2005a,b; Dupont & Olesen 2009). NETCARTO is among the most accurate module-identifying algorithms currently available, yielding nearly the same partitions in different runs and thus highly robust results (Danon et al. 2005; Guimerà & Amaral 2005a,b). The SA-algorithm also assigns each species to a topological role in a network based on the within-module degree zi (the standardized number of links a species i is connected with others in its own module) and the among-module connectivity ci (measuring how connected a species i is to all modules) (Guimerà & Amaral 2005a; Olesen et al. 2007). The parameter zi is calculated as: zi k is k s ks where kis is the number of links of species i to other species in its module si,ks and ks are mean and standard deviation of within-module links (k) for all species in module si. The parameter ci is calculated as: k ci 1 is k NM s 1 2 i where ki is the number of links of species i. Thus, c = 0 if all links of species i are within its own module. If the links of species i are distributed evenly among modules, c 1. Olesen et al. (2007) modified the original role assignment criteria of Guimerà & Amaral (2005a) and tailored them specifically to plant–pollinator networks, such that each plant and pollinator species can play one of the following four roles according to their position in the zc-parameter space: (1) network hub (z > 2.5, c > 0.62); (2) module hub (z > 2.5, c ≤ 0.62); (3) connector (z ≤ 2.5, c > 0.62); or (4) peripheral (z ≤ 2.5, c ≤ 0.62). A network hub is not only highly linked to species of its own but also species of other modules, making it important for the connectivity of among species in both, its own module and the entire network (Olesen et al. 2007). A module hub plays an important role in its own module, increasing its coherence. A connector species is important for among-module connectivity, and consequently network coherence, but plays an inferior role within its own module. Peripheral species have all or most of their relative few interactions within their own module, playing a topologically inferior role in the network (Olesen et al. 2007). References Danon, L., Díaz-Guilera, A., Duch, J. & Arenas, A. (2005). Comparing community structure identification. J. Stat. Mech. Theor. Exp., P09008, 1–10. Dupont, Y.L. & Olesen, J.M. (2009). Modules and roles of species in heathland pollination networks. J. Anim. Ecol., 78, 346–353. Guimerà, R. & Amaral, L.A.N. (2005a). Functional cartography of complex metabolic networks. Nature, 433, 895–900. Guimerà, R. & Amaral, L.A.N. (2005b) Cartography of complex networks: modules and universal roles. Journal of Statistical Mechanics, article no. P02001, 1–13. Kirkpatrick, S., Gelatt, C.D. & Vecchi, M.P. (1983). Optimization by simulated annealing. Science, 220, 671– 680. Olesen, J.M., Bascompte, J., Dupont, Y.L. & Jordano, P. (2007). The modularity of pollination networks. Proc. Natl. Acad. Sci. USA, 104, 19891–19896.