Lec. 18
advertisement

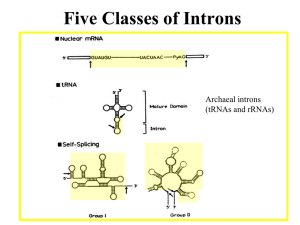
RNA Splicing • RNA splicing is the removal of intervening sequences (IVS) that interrupt the coding region of a gene • Excision of the IVS (intron) is accompanied by the precise ligation of the coding regions (exons) Discovery of Split Genes (1977) • P. Sharp and R. Roberts - 1993 Nobel Prize in Physiology & Medicine • Discovered using R-loop Analysis – Cloned genomic DNAs of a few highly expressed nuclear genes (e.g., hemoglobin, ovalbumin), and certain Adenoviral genes were hybridized to RNA fractions and visualized by EM – Loops form from RNA annealing to the template strand and displacing coding strand of DNA Genomic DNA fragment containing a Globin gene was annealed to large heterogenous nuclear RNA (hnRNA), which contained globin mRNA precursors. Template strand coding Dotted line is RNA DNA Fig. 14.3a template strand Coding strand When genomic globin gene was annealed to cytoplasmic mRNA (which contained mature globin mRNA) got an internal loop of single-stranded DNA (= spliced out intron). Fig. 14.3b Intron Classes & Distribution 1. Group I - common in organelles, nuclear rRNA genes of lower eukaryotes, a few prokaryotes 2. Group II - common in organelles, also in some prokaryotes and archaea 3. Nuclear mRNA (NmRNA) - ubiquitous in eucaryotes 4. Nuclear tRNA- some eucaryotes Relationships of the 4 Intron Classes 1. Each has a distinctive structure. 2. The chemistry of splicing of Groups I, II and NmRNA is similar – i.e, transesterification reactions 3. The splicing pathway for Group II and nuclear mRNA introns is similar. 4. Splicing of Groups I, II and possibly NmRNA introns are RNA-catalyzed Self-Splicing Introns 1. Some Group I & II introns self-splice in vitro in the absence of proteins - are “ribozymes. 2. Conserved secondary structure but not primary sequence. 3. Require Mg2+ to fold into a catalytically active ribozyme. 4. Group I introns also require a guanosine nucleotide in the first step. Tetrahymena rRNA Group I Intron • First self-splicing intron discovered by T. Cech’s lab in 1981 • In the 26S rRNA gene in Tetrahymena • First example of a catalytic RNA! • Nobel Prize in Chemistry to T. Cech and S. Altman (showed that RNase P was a true “turnover” riboenzyme in vivo), 1989 Group I splicing mechanism GOH – guanosine nucleotide, guanosine will work because the phosphates don’t participate in the reaction. The 3’ terminal G of the intron is nearly 100% conserved. In vivo, GTP probably used. Fig. 14.47 Cr.LSU intron: 2ndary structure of a group I intron Old style drawing Newer representation Exon seq. in lower case and boxed Shows how splice sites can be brought close together by “internal guide sequence”. Conserved core 5’ splice site RNA structures seen in Group I introns 1. G U pairs 2. Stacked helices 3. Long-range base pairings (P3 and P7, also form last) 4. Triple helices (3 strands); (P4-P6 junction area) 3-D Model of Tetrahymena rRNA Intron Catalytic core consists of two stacked helices domains: 1. P5 – P4 – P6 –P6a (in green) 2. P9 – P7 – P3 – P8 (in purple) The “substrate is the P1 – P10 domain (in red and black), it contains both the 5’ and 3’ splice sites. A two-metal ion mechanism for group I intron splicing (Second Step) M. R. Stahley et al., Science 309, 1587 -1590 (2005) Guanosine binding site of Group I Introns 1. Mainly the G of a G-C pair in the P7 helix of the conserved core. 2. Highly specific for Guanosine (Km ~20 μM). 3. Also binds the 3-terminal G of the intron in the second splicing step. Splicing Factors for Self-Splicing Introns • Some Group I and many Group II introns can’t self-splice in vitro (need protein factors) • Even self-splicing introns get help from proteins in vivo – Based on fungal (yeast and Neurospora) mutants deficient in splicing of mitochondrial introns (respiratory-deficient) Protein Splicing Factors for Group I (and Group II) Introns • 2 types: 1. Intron-encoded - promote splicing of only the intron that encodes it 2. Nuclear-encoded - Splice organellar introns Split Genes of Yeast Mitochondria From Phil Perlman Proteins encoded within these introns: Mat – maturase (promotes splicing) Endo – DNA endonuclease, promotes intron invasion • Nuclear-encoded splicing factors function by: 1. Promoting correct folding of the intron (cbp2) - CBP2 promotes folding of a cytochrome b intron (bI5) 2. Stabilizing the correctly folded structure (cyt-18) a) Cyt-18 promotes splicing of a number of Mt Group I introns in Neurospora. b) Cyt-18 is also the Mt tyrosyl-tRNA synthetase, dualfunction protein. c) Evolved from the tyrosyl-tRNA synthetase by acquiring a new RNA-binding surface. Alan Lambowitz Figure 5. Models of CYT-18/ΔC424-669 with Bound RNA Substrates(A) Dimeric CYT-18/ΔC424-669 with the T. thermophilus tRNATyr (orange) docked as in the T. thermophilus TyrRS/tRNATyr cocrystal structure (Yaremchuk et al. 2002; PDB ID: 1H3E). Subunits A (Sub. A; magenta) and B (Sub. B; blue) are defined as those that bind the tRNA acceptor and anticodon arms, respectively. Side chains at positions that did or did not give specific EPD-Fe-induced cleavages in the ND1 intron are shown in space-filling representations colored yellow and black, respectively.(B) Stereoview of dimeric CYT-18/ΔC424-669 with docked ND1 intron RNA. The model is based on optimized fit to directed hydroxyl radical cleavage data summarized in Figure 4B. The ND1 intron RNA (residues 27–182) is shown as a green ribbon, with purple balls indicating phosphate-backbone protections from full-length CYT-18 protein (Caprara et al., 1996a), and red ribbon segments indicating EPDFe cleavage sites. The C-terminal domain of T. thermophilus TyrRS (yellow) is shown positioned on subunit B as in the T. thermophilus/tRNATyr cocrystal structure (Yaremchuk et al., 2002).