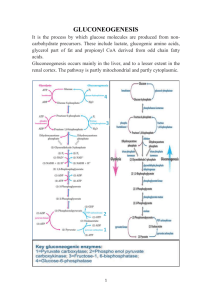
GLUCONEOGENESIS, GLYCOGEN SYNTHESIS & DEGRADATION
advertisement

MIA KUSMIATI Departemen BIOKIMIA FK UNISBA The stimulation of gluconeogenesis by high energy charge and high concentrations of citrate and acetyl-CoA is counterintuitive. Gluconeogenesis is active in the fasting state. the energy for gluconeogenesis is supplied by fatty acid oxidation. During overnight fast: 90 % gluconeogenesis hepar, 10% gluconeogenesis kidney Prolonged fasting: kidney becomes major glucose producing organ (40% total glucose production) Synthesis of glucose from pyruvate utilizes many of the same enzymes as Glycolysis. Three Glycolysis reactions have such a large negative DG that they are essentially irreversible. Hexokinase (or Glucokinase) Phosphofructokinase Pyruvate Kinase. These steps must be bypassed in Gluconeogenesis. Two of the bypass reactions involve simple hydrolysis reactions. (1), Glucokinase (2), phosphofructokinase; (3), pyruvate kinase; (4), pyruvate carboxylase; (5), phosphoenolpyruvate (PEP)-carboxykinase; (6), fructose-1,6-bisphosphatase; (7), glucose-6-phosphatase STIMULATION INHIBITION A, Substrate flow during fasting and in the well-fed state, and the effects of hormones on the amounts of glycolytic and gluconeogenic enzymes. Regulation of enzyme synthesis and degradation is the most important long-term (hours to days) control mechanism. In most cases, the hormone acts by changing the rate of transcription (insulin) B, Short-term regulation of glycolysis and gluconeogenesis by reversibly binding effectors and by: - Phosphorylation/dephosphorylation - Allosteric and competitive effects - phosphorylation. Only pyruvate kinase and phosphofructo-2kinase/fructose-2,6-bisphosphatase are regulated by cAMP-dependent phosphorylation. Synthesis and degradation of fructose-2,6bisphosphate, the most important regulator of phosphofructokinase and fructose-1,6-bisphosphatase. This regulatory metabolite is synthesized and degraded by a bifunctional enzyme that combines the kinase and phosphatase activities on the same polypeptide. cAMP-induced phosphorylation inhibits the kinase activity and stimulates the phosphatase activity of the bifunctional enzyme. , Phosphorylation; , dephosphorylation; , allosteric effect; , stimulation; , inhibition Lactat Pyruvate Glycerol Αlfa keto acid (oxaloacetat, a ketoglutarat) 7 glycolytic Rx are irreversible & are used in the synthesis of glucose from lactat or pyruvate: Carboxylation of pyruvate: A. biotin is a coenzyme Allosteric regualtion B. Transport of oxaloacetate to the cytosol C. Decaboxylation of cytosolic oxaloacetate D. Dephosporilation of Fructose 1,6 biP fructose 6P E. Isomerisasi Fructose 6P Glucose 6P F. Convert glucose 6P free glucose Glucose-6-phosphatase 6 CH OPO 2 2 3 5 O H 4 OH H OH 3 H H 2 CH2OH 1 OH OH glucose-6-phosphate O H H H2O H OH H + Pi H OH OH H OH glucose Hexokinase or Glucokinase (Glycolysis) catalyzes: glucose + ATP glucose-6-phosphate + ADP Glucose-6-Phosphatase (Gluconeogenesis) catalyzes: glucose-6-phosphate + H2O glucose + Pi Glucose-6-phosphatase 6 CH OPO 2 2 3 5 O H 4 OH H OH 3 H H 2 CH2OH 1 OH OH glucose-6-phosphate O H H H2O H OH H + Pi H OH OH H OH glucose Glucose-6-phosphatase enzyme is embedded in the endoplasmic reticulum (ER) membrane in liver cells. The catalytic site is found to be exposed to the ER lumen. Another subunit may function as a translocase, providing access of substrate to the active site. Phosphofructokinase 6 CH OPO 2 2 3 O 5 H H 4 OH 6 CH OPO 2 2 3 1CH2OH ATP HO 3 OH H fructose-6-phosphate O ADP 2 5 Pi H2O 1CH2OPO32 H H HO 3 OH 4 OH 2 H fructose-1,6-bisphosphate Fructose-1,6-biosphosphatase Phosphofructokinase (Glycolysis) catalyzes: fructose-6-P + ATP fructose-1,6-bisP + ADP Fructose-1,6-bisphosphatase (Gluconeogenesis) catalyzes: fructose-1,6-bisP + H2O fructose-6-P + Pi Bypass of Pyruvate Kinase: Pyruvate Kinase (last step of Glycolysis) catalyzes: phosphoenolpyruvate + ADP pyruvate + ATP For bypass of the Pyruvate Kinase reaction, cleavage of 2 ~P bonds is required. DG for cleavage of one ~P bond of ATP is insufficient to drive synthesis of phosphoenolpyruvate (PEP). PEP has a higher negative DG of phosphate hydrolysis than ATP. Pyruvate Carboxylase PEP Carboxykinase O O O O C ATP ADP + Pi C C C O CH3 C O pyruvate O GTP GDP CH2 HCO3 O O C CO2 O oxaloacetate C OPO 32 CH2 PEP Bypass of Pyruvate Kinase (2 enzymes): Pyruvate Carboxylase (Gluconeogenesis) catalyzes: pyruvate + HCO3 + ATP oxaloacetate + ADP + Pi PEP Carboxykinase (Gluconeogenesis) catalyzes: oxaloacetate + GTP PEP + GDP + CO2 Pyruvate Carboxylase PEP Carboxykinase O O O O C ATP ADP + Pi C C C O CH3 GTP GDP O C C CO2 C O pyruvate O CH2 HCO3 O O oxaloacetate OPO 32 CH2 PEP Contributing to spontaneity of the 2-step process: Free energy of one ~P bond of ATP is conserved in the carboxylation reaction. Spontaneous decarboxylation contributes to spontaneity of the 2nd reaction. Cleavage of a second ~P bond of GTP also contributes to driving synthesis of PEP. Pyruvate Carboxylase uses biotin as prosthetic group. N subject to carboxylation O C HN NH CH CH H2C CH S lysine biotin H H3N+ C CH2 CH2 CH2 CH2 NH3 COO lysine residue (CH2)4 O O C NH C (CH2)4 CH NH Biotin has a 5-C side chain whose terminal carboxyl is in amide linkage to the e-amino group of an enzyme lysine. The biotin & lysine side chains form a long swinging arm that allows the biotin ring to swing back & forth between 2 active sites. O O O P -O O O C O OH carboxyphosphate C C O N NH lysine residue CH CH H2C CH S (CH2)4 O O C NH C (CH2)4 CH carboxybiotin NH Biotin carboxylation is catalyzed at one active site of Pyruvate Carboxylase. ATP reacts with HCO3 to yield carboxyphosphate. The carboxyl is transferred from this ~P intermediate to of a ureido group of the biotin ring. Overall: biotin + ATP + HCO3 carboxybiotin + ADP + Pi N At the other active site of Pyruvate Carboxylase the activated CO2 is transferred from biotin to pyruvate: carboxybiotin + pyruvate biotin + oxaloacetate O O O -O C C C C O CH3 pyruvate O N H2C O CH S (CH2)4 C NH R O C C C O CH2 HN NH biotin CH CH H2C C O carboxybiotin CH CH O O NH O oxaloacetate CH S (CH2)4 O C NH R Pyruvate Carboxylase (pyruvate oxaloactate) is allosterically activated by acetyl CoA. [Oxaloacetate] tends to be limiting for Krebs cycle. Glucose-6-phosphatase glucose-6-P glucose Gluconeogenesis Glycolysis pyruvate fatty acids acetyl CoA oxaloacetate ketone bodies citrate Krebs Cycle When gluconeogenesis is active in liver, oxaloacetate is diverted to form glucose. Oxaloacetate depletion hinders acetyl CoA entry into Krebs Cycle. The increase in [acetyl CoA] activates Pyruvate Carboxylase to make oxaloacetate. The main stores of glycogen in the body: 1. Liver to mantain the blood glucose level 2. Skeletal muscleto serve as a fuel reserve for synthesis of ATP during muscle contraction Glycogen is a branched polymer of between 10,000 and 40,000 glucose residues held together by α-1,4 glycosidic bonds Glucose-6-phosphate is isomerized to glucose-1-phosphate by phosphoglucomutase. Glucose-1-phosphate then reacts with uridine triphosphate (UTP) to form UDP-glucose. UDP is attached to C-1 of glucose, and it is therefore this carbon that forms the glycosidic bond. The bond between glucose and UDP is energy rich Metabolic fates of glycogen in the liver (A) and in muscle (B). Note that the liver possesses glucose-6-phosphatase, which forms free glucose both in gluconeogenesis and from glycogen. This enzyme is not present in muscle tissue. Glycogen breakdown serves different purposes in liver and muscle. The liver synthesizes glycogen after a carbohydrate meal and degrades it to free glucose during fasting. The glucose-6-phosphate from glycogen breakdown is cleaved to free glucose by glucose-6-phosphatase. The liver releases this glucose into the blood for use by needy tissues, including brain and blood cells Skeletal muscle synthesizes glycogen at rest and degrades it during exercise. Muscles cannot produce free glucose because they have no glucose-6-phosphatase. Because glycogen degradation produces glucose-6-phosphate without consuming any ATP, anaerobic glycolysis from glycogen produces three rather than two molecules of ATP for each glucose residue. The phosphorylation state of the enzymes is regulated by hormones and their second messengers. 1. Insulin stimulates glycogen synthesis both in the liver and in skeletal muscle. It ensures that excess carbohydrate is stored away as glycogen after a meal. 2. Glucagon stimulates glycogen degradation in liver but not muscle during fasting when the blood glucose level is low. 3. Norepinephrine and epinephrine are powerful activators of glycogen breakdown both in muscle and liver. They mobilize glycogen when glucose is needed to fuel muscle contraction. A, Hormonal effects on the phosphorylation of the glycogen-metabolizing enzymes by protein kinases in the liver. ER, endoplasmic reticulum; GSK3, glycogen synthase kinase-3; B, Hormonal effects on the dephosphorylation of the glycogenmetabolizing enzymes by protein phosphatase-1, and the effects of allosteric effectors. Note that the hormones affect glycogen synthase and glycogen phosphorylase through the protein kinases and the protein phosphatase (phosphatase-1) that regulate their phosphorylation state. , Allosteric effects; , phosphorylation; , dephosphorylation; , activation; , inhibition.