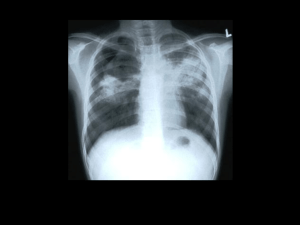
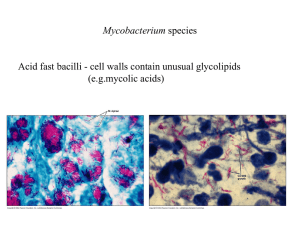
M. tuberculosis
advertisement

Thank you for viewing this presentation. We would like to remind you that this material is the property of the author. It is provided to you by the ERS for your personal use only, as submitted by the author. 2012 by the author “TB and M/XDR-TB: from clinical management to control and elimination” ERS School TB disease and infection: Do we have real news? Martina Sester Department of Transplant and Infection Immunology Saarland University; Germany May 23-26, 2012 in Bucharest, Romania Overview – Facts and news…* • Worldwide epidemiology of tuberculosis • M. tuberculosis infection: continuum from latency to active disease – Implications for diagnosis of M. tuberculosis infection • Host-pathogen interactions – Role of innate immunity • The vaccine pipeline *an immunologist´s view on 2011´s news… Tuberculosis – the facts • 7. position of leading causes of deaths • 1/3 of the world's population could be infected • > 80% can be cured • prevention can be > 90% effective Global tuberculosis control: WHO report 2011 Tuberculosis – the facts • • • • • 1.45 million people died in 2010 due to TB equally to 3800 deaths per day 8.8 million new cases of TB in 2010 Global incidence rate of 128/100 000 Most cases occurred in – Asia (59%) and – Africa (26%) WHO report 2011 Estimated TB incidence rates 2010 WHO report 2011 The global burden of TB in 2010 in relation to HIV co-infection All forms of TB HIV-associated TB Estimated number of cases 8.8 Mio (8.5–9.2 Mio) 1.1 Mio (13%)* (1.0–1.3 Mio) *82% of TB cases among people living with HIV originate from the African region Estimated number of deaths 1.45 Mio (1.2–1.5 Mio) 0.35 Mio (0.32–0.39 Mio) WHO report 2011 Estimated HIV prevalence in new TB cases 2010 WHO report 2011 Trends in TB incidence rates Lawn and Zumla (2011) Lancet 378: 57 Overview – Facts and news… • Worldwide epidemiology of tuberculosis • M. tuberculosis infection: continuum from latency to active disease – Implications for diagnosis of M. tuberculosis infection • Host-pathogen interactions – Role of innate immunity • The vaccine pipeline TB disease and infection - definitions TB disease • Detection of M. tuberculosis and/or clinical symptoms compatible with tuberculosis Latent infection with M. tuberculosis (LTBI) • Presence of an immune response in a skin test or an IFN-g release assay (IGRA) • Absence of clinical symptoms Natural course of M. tuberculosis infection M. tuberculosis Immunosuppression exposure 1% infection Progression Latency 90% Latency Years after contact Recent contacts Low TB High TB prevalence prevalence Successful treatment LTBI Old healedTB/LTBI TB TB disease 5% 2-5% Live bacilli? Immunosuppression Chemoprophylaxis efficient Protective immunity Never TB Bacterium extinguished? Chemoprophylaxis not necessary Prevalence of latent infection with M. tuberculosis and risk for progression Horsburg and Rubin (2011) N Engl J Med 364:1441 Immunodiagnosis of latent M. tuberculosis infection antigens/ peptides APC PPD ESAT-6/CFP-10/TB7.7 Negative controls Positive controls, i.e. mitogens PHA/SEB Skin test IFN-g release assay T cell cytokine induction activation/ cytokine induction cytokine induction ELISA ELISPOT assay QuantiFERON TB gold T.SPOT.TB Flow-cytometry activation marker cytokine induction IGRA cytokine PPV and NPV of immune-based assays for the development of tuberculosis Test PPV NPV TST 2.3-3.3 99.7 QFT-G-IT 2.8-14.3 99.8 T-SPOT.TB 3.3-10.0 97.8 Diel et al. (2011) Eur Respir J 37: 88 Sensitivity and specificity of immunebased assays to diagnose active TB Test Sensitivity Specificity TST 0.65 0.75 0.80 0.79 extrasang. 0.48 0.82 0.59 0.82 QFT-G-IT blood 0.81 T-SPOT.TB extrasang. 0.88 blood summary of pooled values Sester, Sotgiu et al. Eur Respir J (2011), 37: 100 New experimental tests LTBI • Antigen different from the commercial RD1 peptides • Markers different from IFN-g • Readouts different from ELISA or ELISPOT • Biological sample different from blood More details in the following talk: IGRA testing to diagnose TB disease and infection. What is new in clinical practice and for programmatic management? - D. Goletti, M Sester Diagnosis of active tuberculosis • • • • • Patient history Chest X-ray Culture Acid-fast bacilli staining Nucleic acid amplification testing New experimental tests active tuberculosis • • • • Assays for childhood tuberculosis Assays for smear negative tuberculosis Faster assays Improved NAAT tests (i.e. Xpert MTB/RIF assay) More details in the following talk: The new horizons of molecular diagnosis: do we still need conventional microbiology? - D. Cirillo Overview – Facts and news… • Worldwide epidemiology of tuberculosis • M. tuberculosis infection: continuum from latency to active disease – Implications for diagnosis of M. tuberculosis infection • Host-pathogen interactions – Role of innate immunity • The vaccine pipeline Pathogenesis and immune effector mechanisms Kaufmann (2010) Immunity 33: 567 Pathogenesis and immune effector mechanisms Macrophage activation Interplay IFN-g/VitD signaling Apoptosis/necrosis of Macrophages affects bacterial growth and Tcell priming Immunopathogenesis of IRIS Kaufmann (2010) Immunity 33: 567 Role of innate immunity • Controlling early pathogen growth • Instructing adaptive immunity without innate immunity M. tuberculosis load without adaptive immunity normal immunity infection time Role of innate immunity • Controlling early pathogen growth • Instructing adaptive immunity Fremond et al. (2004) J Clin Invest 114: 1790; Feng et al. (2005) J Immunol 174: 4185 Apoptosis versus necrosis • Apoptotic macrophages decrease bacterial load and accelerate T-cell priming • Necrotic macrophages increase bacterial load and slow down T-cell priming Divangahi et al. (2010) Nat Immunol 11: 751; Divangahi et al. (2009) Nat Immunol 10: 899 Apoptosis versus necrosis • Apoptotic macrophages decrease bacterial load and accelerate T-cell priming • Necrotic macrophages increase bacterial load and slow down T-cell priming Divangahi et al. (2010) Nat Immunol 11: 751; Divangahi et al. (2009) Nat Immunol 10: 899 Suppression of apoptosis as innate defence mechanism of virulent strains Interference with plasma membrane repair Decreased bacterial load Accelerated T-cell priming Increased bacterial load Delay in T-cell priming Divangahi et al. (2010) Nat Immunol 11: 751; Divangahi et al. (2009) Nat Immunol 10: 899; Behar et al. (2010) Nat Rev Microbiol 8: 668 Pathogenesis and immune effector mechanisms Macrophage activation Interplay IFN-g/VitD signaling Kaufmann (2010) Immunity 33: 567 Vitamin D deficiency and susceptibility to tuberculosis Vitamin D3 at start of antimicrobial treatment, and after 14, 28, and 42 days Martineau et al. (2011) Lancet 377: 242 Vitamin D deficiency and susceptibility to tuberculosis Vitamin D3 at start of antimicrobial treatment, and after 14, 28, and 42 days Median time to culture conversion • 36·0 days in the intervention group and • 43·5 days in the placebo group Adjusted hazard ratio 1·39, 95% CI 0·90–2·16; p=0.14. Martineau et al. (2011) Lancet 377: 242 Vitamin D deficiency and susceptibility to tuberculosis Effect of TaqI genotype Enhanced response with tt genotype (8.09, 95% CI 1.36–48.01; p=0.02) Tt genotype (0.85, 95% CI 0.45–1.63; p=0.63) TT genotype (1.13, 95% CI 0.60–2.10; p=0.71) Median time to sputum culture conversion • 36·0 days - intervention group • 43·5 days - placebo group Adjusted hazard ratio 1·39, 95% CI 0·90–2·16; p=0.14. Martineau et al. (2011) Lancet 377: 242 Active TB is associated with vitamin D deficiency Patients from Cape Town Effect of vitamin D deficiency is more pronounced in HIV infected patients Martineau et al. (2011) Proc Natl Acad Sci U S A 108: 19013 Seasonal variation in vitamin D status and tuberculosis notifications Martineau et al. (2011) Proc Natl Acad Sci U S A 108: 19013 Antimicrobial effect of vitamin D and T-cell derived IFN-g Fabri et al. (2011) Sci Transl Med 3: 104ra102 Antimicrobial effect of vitamin D and T-cell derived IFN-g Induction of autophagy Fabri et al. (2011) Sci Transl Med 3: 104ra102 Antimicrobial effect of vitamin D and T-cell derived IFN-g Induction of antimicrobial peptides Fabri et al. (2011) Sci Transl Med 3: 104ra102 Mechanistic link - vitamin D deficiency and HIV-induced immunodeficiency Antimicrobial effect via induction of antimicrobial peptides and autophagy Martineau et al. (2011) Proc Natl Acad Sci U S A 108: 19013 Fabri et al. (2011) Sci Transl Med 3: 104ra102 Pathogenesis and immune effector mechanisms Immunopathogenesis of IRIS Kaufmann (2010) Immunity 33: 567 HIV-associated IRIS Immune reconstitution inflammatory syndrome • May occur in up to 30% of HIV infected patients after start of ART • Tissue destructive inflammation • Microbial co-infections as risk factor • Recovering CD4 T cells as immediate effectors? • Pathological T cell responses? Mouse model for lymphopenia-induced IRIS IRIS develops in context of • Chronic microbial infection • CD4 T cell deficiency Barber et al. (2012) Nat Rev Microbiol 10: 150 Model for IRIS involving a dysregulated innate immune response Barber et al. (2012) Nat Rev Microbiol 10: 150 Overview – Facts and news… • Worldwide epidemiology of tuberculosis • Infection cycle • M. tuberculosis infection: continuum from latency to active disease – Implications for diagnosis of M. tuberculosis infection • Host-pathogen interactions – Role of innate immunity • The vaccine pipeline Characteristics of successful vaccines CMV Rappuoli & Aderem (2011) Nature 473: 463 BCG vaccine • Developed 1921 • 120 Mio doses administered/year • Provides 80% protection against severe and disseminated disease in children • 50% risk reduction in adults (0-80% efficacy) – Genetic divergence – Differences in T-cell response Vaccine candidates • Subunit and live viral vectors – Antigens: ESAT-6, TB10.4, Ag85A, Ag85B, Mtb32 and 39 and fusions thereof – Adjuvants: IC31, AS01, AS02, CAF01 – Live viral vectors: Adenovirus, Vaccinia • Live attenuated or killed bacteria – rBCG – M. tuberculosis – M. vaccae Adjuvants used for fusion proteins • Serve to improve immunogenicity • i.e. ligands for pattern recognition receptors Kaufmann (2011) Lancet Infect Dis 11: 633 Vaccines – where they should act • Pre-exposure vaccination • Post-exposure vaccination • Therapeutic vaccination Kaufmann (2011) Lancet Infect Dis 11: 633 Therapeutic vaccine candidates Pre-/post-exposure vaccines Most advanced vaccine candidates 12 candidates have reached clinical trials (to replace BCG) (after BCG) Animal models: Prevention of tuberculosis, no eradication of M. tuberculosis Kaufmann (2011) Lancet Infect Dis 11: 633 Most advanced vaccine candidates 12 candidates have reached clinical trials (to replace BCG) (after BCG) Animal models: Prevention of tuberculosis, no eradication of M. tuberculosis Kaufmann (2011) Lancet Infect Dis 11: 633 Most advanced vaccine candidates 12 candidates have reached clinical trials (to replace BCG) (after BCG) Animal models: Prevention of tuberculosis, no eradication of M. tuberculosis Kaufmann (2011) Lancet Infect Dis 11: 633 Most advanced vaccine candidates 12 candidates have reached clinical trials (to replace BCG) (after BCG) Animal models: Prevention of tuberculosis, no eradication of M. tuberculosis Kaufmann (2011) Lancet Infect Dis 11: 633 Post-exposure vaccines Effective vaccine for pre- and post-exposure H56 vaccine • Early antigen Ag85B • Early antigen ESAT-6 • Latency antigen Rv2660c – expressed during starvation Vaccination of mice Aagaard et al. (2011) Nat Med 17: 189 Effective vaccine for pre- and post-exposure H56 vaccine • Pre-exposure 6 weeks after challenge 24 weeks after challenge • T cells induced are polyfunctional Aagaard et al. (2011) Nat Med 17: 189 Effective vaccine for pre- and post-exposure H56 vaccine • Post-exposure 35 weeks p.i. blood 35 weeks p.i. spleen Aagaard et al. (2011) Nat Med 17: 189 Effective vaccine for pre- and post-exposure H56 vaccine • Post-exposure 2 vaccinations 2 vaccinations 2 vaccinations 2 vaccinations 35 weeks p.i. blood 35 weeks p.i. spleen 3 vaccinations 2 vaccinations Analysis between 23 and 43 weeks p.i. Aagaard et al. (2011) Nat Med 17: 189 Future candidates? • Up to now, no vaccine candidate has achieved sterilising immunity Conclusions • Understanding the continuum from latency to active disease will lead to improved diagnosis of patients at risk and targeted therapy • Knowledge of the role of innate immunity will lead to improved understanding of host-pathogen interactions • Rationale vaccine design has achieved success, but clinical studies are still proceeding at slow pace