ppt - eweb.furman.edu
advertisement

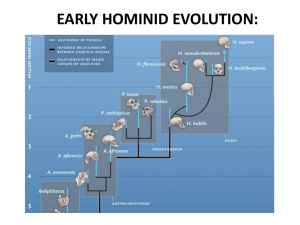
IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: Linnaean Classification Hominidae Pongidae Hylobatidae Apes = primates (grasping hands, binocular vision) with no tails IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: Phylogenetic Classification Hylobatidae Hominidae IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: 1-4% difference Hylobatidae Hominidae IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: How can the small genetic difference account for the dramatic differences that occur between these species? IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: 2. Patterns: a. Morphological and Behavioral Homo sapiens Chimps, Gorillas Larger Head/body ratio smaller Smaller jaw/head ratio larger Shorter limb/body ratio longer Less hair more hair Better learning poorer learning IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: 2. Patterns: a. Morphological and Behavioral Differences correlate with developmental differences Juvenile Primate Adult Primate Larger Head/body ratio smaller Smaller jaw/head ratio larger Shorter limb/body ratio longer Less hair more hair Better learning poorer learning IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: 2. Patterns: a. Morphological and Behavioral Big head, short limbs Small head, long limbs IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: 2. Patterns: a. Morphological and Behavioral b. Genetic Developmental genes can have profound effects on the final morphology of the organism. Human Chimp Primate Developmental Trajectory IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: 2. Patterns: a. Morphological and Behavioral b. Genetic What are some of these genetic differences? The HAR1 RNA molecule. - not a coding RNA; probably regulatory - nearby genes associated with transcriptional regulation and neurodevelopment are upregulated in humans. - only 2 changes in sequence between chicks and chimps; 18 between chimps and humans. “HAR” stands for “human accelerated region” – changing more rapidly than drift can explain. why? Selection. -Changes result in a profound change in RNA structure and, presumably, binding efficiency. Beniaminov A, Westhof E, and Krol A. 2008. Distinctive structures between chimpanzee and human in a brain noncoding RNA. RNA 14:1270-1275. Beniaminov A et al. RNA 2008;14:1270-1275 IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: 2. Patterns: a. Morphological and Behavioral b. Genetic c. Fossils Chimpanzee Human Sahelanthropus tchadensis Sahelanthropus tchadensis – discovered in Chad in 2001. Dates to 6-7 mya. Only a skull. Is it on the human line? Is it bipedal? Probably not (foramen magnum). Primitive traits, as a common ancestor might have. Ardipithecus ramidus: 4.3-4.5 mya. Discovered in 1994 by Haile-Sailasse, Suwa, and White, with the most complete fossils were not described until 2009. Arboreal, but facultatively bipedal. Grasping toes. IV. The Domain Eukarya E. Fungi F. Animals G. Human Evolution 1. Overview: 2. Patterns: a. Morphological and Behavioral b. Genetic c. Fossils Sahelanthropus tchadensis Chimpanzee Human Homo sapiens Competitive contraction? Radiation of Bipedal Hominids Innovation: Bipedality Australopithecus afarensis: 2.8-3.9 mya. A femur discovered in 1973 by Donald Johansson suggested an upright gait, confirmed by his discovery in 1974 of the ‘Lucy” specimen. Also, the Laetoli prints (found by Mary Leakey) were probably made by A. afarensis, and in 2006, a juvenile A. afaresis was found. Kenyanthropus platyops: 3.2-3.5 mya – Discovered by Meave Leakey’s team at Lake Turkana; most dispute it warrants another genus, and some even include it in A. afarensis. Australopithecus garhi: 2.5-2.6 mya; discovered by Asfaw and White in 1996, but the skull below was discovered by Haile-Selasse in 1997. The tooth morphology is a bit different from A. afarensis and A. africanus, being much larger than even the robust forms. There are associated stone tools! Australopithecus sebida: 1.9 mya, describe in 2010 by LE Berger; it has many characteristics like A. africanus, but also similar to genus Homo. Australopithecus africanus: 2-3 mya, discovered by Raymond Dart in South Africa in 1924 – the ‘Taung child’. Then, in 1947, Robert Broom found a skull he classified as Plesianthropus, but was grouped with A. africanus. Paranthropus aethiopicus: 2.5-2.7 mya, discovered by Alan Walker and Richard Leakey, the “black skull” is one of the most imposing hominid fossils there is! Aside from the high cheekbones and the sagittal crest, it has similar proportions to A. afarensis and is probably a direct descendant. It probably gave rise to the “robust” lineage of Paranthropus. Paranthropus boisei: 1.2-2.6 mya. Discovered by Mary Leakey in Olduvai Gorge in 1959, it was originally classified as Zinjanthropus and nicknamed “Zinj” or “nutcracker man” because of the large grinding molars. Paranthropus robustus: 1.2-2.0 mya. Discovered in South Africa in 1938 by Robert Broom. Competitive contraction? Radiation of Bipedal Hominids Innovation: Bipedality Homo habilis: 1.4-2.3 mya, discovered by Louis and Mary Leakey, in association with stone tools. “Handy man”. Longer arms and smaller brain than other members of the genus. Homo rudolphensis: 1.9 mya; Discovered by Richard and Meave Leakey’s team. Different from H. habilis, yet a contemporary. Either may be ancestral to recent Homo. Homo georgicus: 1.7 mya; the oldest hominid fossils found outside of Africa – found in Dmanisi, Georgia, in 1999. Thought to be a potential intermediate between H. habilis and H. ergaster/H. erectus. Homo ergaster (H. erectus): 1.3-1.8 mya, the most complete fossil hominid skeleton was discovered in 1984 by Alan Walker who called it “Turkana Boy”. Some consider this species intermediate to H. habilis and H. heidelbergensis/H. sapiens, leaving H. erectus as a distinct Asian offshoot of the main line to H. sapiens. However, most paleontologists suggest that H. ergaster is the African ancestor – even a chronospecies or population - of H. erectus, which is ancestral to more recent Homo species. Homo erectus: 0.2-1.8 mya; originating in Africa, but then leaving for Asia (Peking and Java Man). Discovered in Java by Eugene Dubois in 1891. Certainly one of the most successful hominid species in history; perhaps lasting as relictual species on islands in Indonesia as: Homo floresiensis: 94,000-13,000 years, discovered by Mike Mormood on the island of Flores. Shoulder anatomy is reminiscent of H. erectus, but could be an allometeric function of the small size (3 ft). Homo cepranensis: 350,000-500,000 years old; discovered by Italo Biddittu in 1994 in Italy. It is just a skull cap, but seems to be intermediate between H. erectus and H. heidelbergensis. Homo antecessor: 800,000-1.2 mya; fossils from 20 individuals found in Spain in 1994-5; may be H. heidelbergensis or an intermediate between it and H. ergaster. Homo heidelbergensis: 250-600,000 in Europe and Africa; ancestral to H. neaderthalensis and H. sapiens; may have buried their dead. Homo rhodesiensis: 125-300,000; may be H. heidelbergensis or intermediate to it and H. sapiens. Homo neaderthalensis: 30,000-150,000; first discovered in 1829. Descended from H. heidelbergensis. Homo sapiens idaltu: 160,000 – oldest Homo sapiens fossil – found in Africa in 2003… afar valley. Competitive contraction? Radiation of Bipedal Hominids Innovation: Bipedality