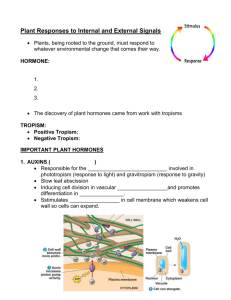
PHYTOHORMONES A phytohormone is an organic compound that is synthesized in one part of a plant and translocated to another part where, in low concentrations (1μM or less), it causes a physiological response. By doing so, phytohormones regulate various physiological processes of plant growth and development. In chronological order of their discovery, these include: 1. Ethene/ethylene 2. Auxins (indole-3-acetic acid – IAA) 3. Gibberellins (GA) 4. Cytokinins 5. Abscisic acid (ABA) ETHENE/ETHYLENE Ethylene is another class of hormones with a single representative. It is a simple gaseous hydrocarbon with the chemical structure H2C=CH2. Ethylene is apparently not required for normal vegetative growth, although it can have a significant impact on the development of roots and shoots. Ethylene appears to be synthesized primarily in response to stress and may be produced in large amounts by tissues undergoing senescence or ripening. It is commonly used to enhance ripening in bananas and other fruits that are picked green for shipment as well. Ethylene Synthesis In seedlings, it is produced at the shoot apex. This may be as a result of the high amounts of auxins present in this region, because auxins stimulate ethylene formaton. • Nodes of dicot seedling stems produce much more ethylene than internodes do. • Roots release relatively small amounts of this gas (auxin treatment causes the rate to rise). • Production in leaves generally rises slowly until the leaves become senescent and abscise. • Flowers also synthesize ethylene, and this gas often causes their senescence and abscission. • In many fruits, little ethylene is produced until just before ripening, when the concentration of this gas rises from almost undetectable clearly detectable amounts. • Mechanical effects such as gently rubbing a stem or leaf, increased pressure, pathogenic microorganisms, and insects increase ethylene production. AUXINS Although originally discovered in relation to growth, auxin influences nearly every stage of a plant's life cycle from germination to senescence. The morphology of a plant depends on the directed movement of auxin via the polar transport system, which maintains both basic shootroot polarity and polarized outgrowth throughout development. IAA plays an important role in stem elongation and is also necessary for growth of fruits. It is commonly assumed that it is essential for growth of leaves and flowers, but there is little evidence for this. It stimulates root growth but at low concentrations. Auxins of all types stimulate many kinds of cells to produce ethylene, which retard elongation of both roots and stems. As such, IAA at high concentrations inhibits root growth through ethylene formation. IAA also stimulates adventitious root development on stems through stimulation of cell division of an outer layer of phloem. In most species, the apical bud exerts an inhibitory influence (apical dominance) upon the lateral buds, preventing their development. This extra production of undeveloped buds has definite survival value, for if the apical bud is damaged or removed by a grazing animal, a lateral bud will then grow out and become the leader shoot. Another dominant effect of the shoot apex is to cause branches below to grow out somewhat horizontally. The inhibitory effect of the apical bud and/or shoot apex is due to IAA, either directly or indirectly through formation of another inhibitory compound or even by causing lack of some nutrient or stimulatory hormone. BIO 3103 – Plant Physiology 4 Apart from these effects, IAA functions in phototropism and geotropism as well as in delaying abscission of leaves, flower and fruits. However, through ethylene stimulation, IAA may promote senescence and abscission as well as fruit ripening, flowering in plant species which characteristically do so under the influence of ethylene, and femaleness in dioecious flowers. It has a marked effect on nucleic acid synthesis, stimulating faster production of RNA and DNA, hence proteins. Auxin Synthesis IAA is structurally similar to the amino acid tryptophan and is synthesized from it. This occurs by removal of the amino group and the terminal carboxyl group from tryptophan’s side chain. The enzymes necessary for this conversion are most active in young developing tissues, such as shoot meristems and young leaves and fruits. In these tissues, auxin contents are also highest, suggesting that IAA is synthesized there. It moves slowly in plants through other living cells and its transport is polar since it always occurs in a basipetal (baseseeking) direction in stems and an acropetal (apex-seeking) direction in roots. Additionally, it movement is an active process, requiring ATP to move against a concentration gradient. Because of its potency, plants have developed mechanisms to get rid of IAA when it is no longer needed. It can be combined with other molecules to form certain derivatives called “bound auxins” or it can be degradatively removed by Mn2+ -dependent oxidation resulting in loss the its carboxyl group. GIBBERELLINS Gibberellins were first discovered in Japan in studies with diseased rice plants that grew excessively tall. These plants often could not support themselves and eventually died from combined weakness and parasite damage. This phenomenon was later discovered by Kurosawa (in 1926) to be caused by a fungus, Gibberella fujikuroi. In the 1930s, Yabuta and Hayashi isolated the active compound, gibberellins, from the fungus. However, much interest was not initially given to this phytohormone until the 1950s due to preoccupation with IAA (which was also discovered around that time), lack of early contact with the Japanese, and then World War II. At least 50 gibberellins have now been discovered in fungi and plants. They are abbreviated GA, with a subscript such as GA1 , GA2 , GA3 , and so on, to distinguish them. GA3 has been studied much more than the others, because of its availability, and is often referred to as gibberellic acid. GA have the unique ability among plant hormones to stimulate extensive growth of intact plants by enhancing elongation. Many experiments have shown how GA stimulate growth of dwarf plants. However, gibberellins have little direct effect on root growth, and they inhibit adventitious root formation. In seeds, they are capable of overcoming dormancy and enhancing cell elongation so the radicle can push through the endosperm and seed coat. Additionally, they help to make food available to the embryo of seeds by stimulating formation of hydrolytic enzymes that break down reserve substances into more useful forms. Apart from this, gibberellins cause parthenocarpic (seedless) fruit development in some species. Gibberellins also induce flowering in some plants, especially those growing in cold environments or exposed to inadequate hours of light. It may delay senescence in leaves and fruits and participate in phototropism and geotropism. They induce maleness in dioecious flowers. The overall effects of gibberellins on plants are: 1) they stimulate cell division in the shoot apex, 2) they stimulate cell growth because they increase hydrolysis of starch, fructosans, and sucrose into glucose and fructose molecules, and 3) they sometime increase wall plasticity. Gibberellins Synthesis • Seeds of beans and many other dicots are rich sources of gibberellins. • Young leaves are the major sites of active gibberellin synthesis, just as they are for IAA. Mature leaves have little ability to synthesize either hormone type. • Roots also synthesize gibberellins in significant quantities, even though they have little effect on root growth. It has been suggested that gibberellins are formed in roots and move to the shoot via the xylem vessels. • Gibberellins are present in fruits, where they stimulate growth. CYTOKININS Sometime around 1920, Haberlandt discovered that an unknown compound present in vascular tissues of various plants stimulated cell division, causing cork cambium formation and wound healing in cut potato tubers. He also deduced that the wounded parenchyma cells produced another compound that participated in the division and healing process. In 1954, Skoog and colleagues found a highly active, purine-like compound that promoted cytokinesis in aged or autoclaved herring sperm DNA (named “kinetin). Although kinetin is not present in plants, Steward (still in the 1950s) later identified several related compounds (known as cytokinins) in coconut milk that stimulated cell division in carrot root tissues. When a mature but still active leaf is cut off, it begins to lose chlorophyll, RNA, proteins, and lipids from chloroplast membranes more rapidly than if it were still attached, even if it is provided with mineral salts and water through the cut end. This premature aging and senescence occur especially fast if the leaves are kept in darkness. Sometimes, adventitious roots form at the base of the petiole in dicot leaves, and then senescence of the blade is greatly delayed. This is due to formation of cytokinins in the newly formed roots and passage through xylem to the leaf. Cytokinins also promote lateral bud development, which may be dramatic enough to overcome apical dominance in some species such as tobacco. This effect is reversed in roots. Cytokinins are suggested to promote increased growth in seedlings that were germinated in the dark, even if the provided light energy is too low to allow photosynthesis. This is because cytokinins promote functional chloroplast development through formation of stroma lamellae. Cytokinins Synthesis • Cytokinins are relatively abundant in young fruits and seeds, in young leaves, and in root tips. It seems logical that they are synthesized there, but the possibility of transport from some other site cannot be dismissed. • For roots, synthesis is probably involved, because if the roots or stems are cut off and xylem exudates are collected from the remaining portions, cytokinins continue to be exuded up to four days. A conclusion is that root tips synthesize cytokinins and transport them through the xylem to all parts of the plant. This might explain their accumulation in young leaves, fruits, and seeds into which xylem transport is effective. Abscisic acid (ABA) Developmental and physiological effects of ABA Abscisic acid plays primary regulatory roles in the initiation and maintenance of seed and bud dormancy and in the plant's response to stress, particularly water stress. In addition, ABA influences many other aspects of plant development by interacting, usually as an antagonist, with auxin, cytokinin, gibberellin, ethylene, and brassinosteroids. The primary functions of ABA are (1) prohibiting precocious germination and promoting dormancy in seeds and (2) inducing stomatal closure and the production of molecules that protect cells against desiccation in times of water stress. Abscisic acid is the growth inhibitor hormone in plants. It is synthesized within the stem, leaves, fruits, and seeds of the plant. It acts as an antagonist to Gibberellic acid. It is also referred to as the stress hormone because it helps by increasing the tolerance of plants to different kinds of stress. Functions of Abscisic acid • Stimulates the closing of stomata in the epidermis • Favors in the development and maturation of seeds • Inhibits plant metabolism and seed germination • Mainly involved in regulating abscission and dormancy • Used as a spraying agent on trees to regulate the dropping of fruits • Induces dormancy in seeds and helps in withstanding desiccation and other unfavorable growth factors