Chapter 8 Population genetics II
advertisement

Self-fertilization and sibling mating most
extreme forms of inbreeding, but matings
between more distant relatives (e.g.
cousins) has same effect on frequency
of homozygotes, but rate is slower.
F = Coefficient of inbreeding: probability
that two alleles in an individual are
identical by descent (both alleles are
copies of a particular ancestor’s allele in
some previous generation).
F increases as relatedness increases.
If we compare heterozygosity of inbred
population Hf with that of a random mating
population Ho relationship is
Hf = Ho (1-F)
Anytime F>0 frequency of heterozygotes is
reduced and frequency of homozygotes
naturally increases.
Calculating F. Need to use pedigree
diagrams.
Example: Female is daughter of two halfsiblings.
Two ways female could receive alleles
that are identical by descent.
Male
Female
Female
Male
Male
Fig 6.27a
Half-sibling mating
Fig 6.27b
Total probability of scenario is 1/16 + 1/16
= 1/8.
Inbreeding increases frequency of
homozygotes and thus the probability
that deleterious alleles are visible to
selection.
In humans, children of first cousins have
higher mortality rates than children of
unrelated individuals.
Each dot on
graph
represents
mortality
rates for a
human
population.
Fig 6.28
Mortality rate
for children
of cousins
consistently
about 4%
higher than
rate for
children of
non-relatives.
In a study of 2760 individuals from 25
Croatian islands Rudan et al. found a
strong positive relationship between high
blood pressure and the inbreeding
coefficent.
Inbreeding depression also documented
in studies of wild animals.
E.g. Great Tit. Two studies show that
survival of inbred nestlings is lower than
that of outbred individuals and that
hatching success of inbred eggs is lower
than that of outbred eggs.
Fig. 6.30
Migration: movement of alleles between
populations.
Migration can cause allele and
genotype frequencies to deviate from
Hardy-Weinberg equilibrium.
Consider Continent-Island migration
model.
Migration from island to continent will
have no effect of continental allele
frequencies. Continental population
much larger than island.
However continent to island migration
can greatly alter allele frequencies.
Lake Erie water snakes. Snakes range in
appearance from unbanded to strongly
banded.
Banding caused by single locus: banded
allele dominant over unbanded.
Mainland: almost all snakes banded.
Islands many snakes unbanded.
Unbanded snakes have selective
advantage: better camouflage on
limestone rocks. Camouflage very
valuable when snake is young.
Fig 6.6
If selection favors unbanded snakes on
islands why aren’t all snakes unbanded?
Migration introduces alleles for banding.
Fig 6.7
A unbanded, B+C some banding, D strongly banded
Migration of snakes from mainland
makes island populations more like
mainland.
This is general effect of migration:
Homogenizes populations (making them
resemble each other).
Genetic drift results from the influence of
chance. When population size is small,
chance events more likely to have a
strong effect.
Sampling errors are very likely when small
samples are taken from populations.
Assume gene pool where frequency A1 =
0.6, A2 = 0.4.
Produce 10 zygotes by drawing from
pool of alleles.
Repeat multiple times to generate
distribution of expected allele
frequencies in next generation.
Fig 6.11
Allele frequencies much more likely to
change than stay the same.
If same experiment repeated but
number of zygotes increased to 250 the
frequency of A1 settles close to
expected 0.6.
6.12c
Founder Effect: when population
founded by only a few individuals allele
frequencies likely to differ from that of
source population.
Only a subset of alleles likely to be
represented and rare alleles may be
over-represented.
Silvereyes colonized South Island of New
Zealand from Tasmania in 1830.
Later spread to other islands.
http://photogallery.canberra
birds.org.au/silvereye.htm
6.13b
Analysis of microsatellite DNA from
populations shows Founder effect on
populations.
Progressive decline in allele diversity from
one population to the next in sequence
of colonizations.
Fig 6.13 c
Norfolk island Silvereye population has
only 60% of allelic diversity of Tasmanian
population.
Founder effect common in isolated
human populations.
E.g. Pingelapese people of Eastern
Caroline Islands are descendants of 20
survivors of a typhoon and famine that
occurred around 1775.
One survivor was heterozygous carrier of
a recessive loss of function allele of
CNGB3 gene.
Codes for protein in cone cells of retina.
4 generations after typhoon
homozygotes for allele began to be
born.
Homozygotes have achromotopsia
(complete color blindness, extreme light
sensitivity, and poor visual acuity).
Achromotopsia rare in most populations
(<1 in 20,000 people). Among the 3,000
Pingelapese frequency is 1 in 20.
High frequency of allele for
achromotopsia not due to a selective
advantage, just a result of chance.
Founder effect followed by further
genetic drift resulted in current high
frequency.
Effects of genetic drift can be very strong
when compounded over many
generations.
Simulations of drift. Change in allele
frequencies over 100 generations. Initial
frequencies A1 = 0.6, A2 = 0.4. Simulation
run for different population sizes.
6.15A
6.15B
6.15C
Populations follow unique paths
Genetic drift has strongest effects on small
populations.
Given enough time even in large
populations genetic drift can have an
effect.
Genetic drift leads to fixation or loss of
alleles, which increases homozygosity and
reduces heterozygosity.
6.15D
6.15E
6.15F
Genetic drift produces steady decline in
heterozygosity.
Frequency of heterozygotes highest at
intermediate allele frequencies. As one
allele drifts to fixation number of
heterozygotes inevitably declines.
The Hardy-Weinberg model provides an
idealized picture of how allele
frequencies and genotype frequencies
are expected to change over time in a
large population.
The Wright-Fisher model is a similar model
but applies to small populations.
The W-F model retains the assumptions of
the H-W model except for population
size and in the model only a small
sample of gametes are drawn at
random from the gene pool.
The small sample drawn mimics the
effects of drift because allele
frequencies in the sample can differ a lot
from the starting gene pool.
We know that genetic drift leads to a loss
of heterozygosity over time.
Alleles going to fixation naturally reduce
the diversity of alleles in the population
and without allelic diversity
heterozygosity must decline.
In a Wright-Fisher population expected
heterozygosity declines on average by a
factor of 1/2N per generation, where N is
population size.
When N is large 1/2N is very small so we
expect heterozygosity to decline slowly.
Conversely, with a small population 1/2N
is large and heterozygosity will decline
quickly as a result of drift.
Recall that F = Coefficient of inbreeding:
probability that two alleles in an
individual are identical by descent (both
alleles are copies of a particular
ancestor’s allele in some previous
generation).
Also recall that two alleles can be
identical in their genetic sequence (e.g.
both A1), but if they did not come from
the same shared ancestor they are not
considered identical by descent.
Consider a gamete pool to which all
parents contribute equally and from
which offspring are produced by
drawing two gametes at random.
With a population of N parents each
contributing 2 alleles there is a total pool
of 2N alleles.
Under these circumstances the
probability of an offspring having two
alleles derived from the same parental
copy is 1/2N.
{Note the probability is not 1/2N * 1/2N
because the first allele can be any allele
and there is then a 1/2N chance it is the
same as that first one}
The parental generation had a level of
heterozygosity that we call Hparental.
Thus the offspring generations
heterozygosity is
Hoffspring = [1 – 1/2N] * Hparental
See box 8.2 for derivation
Buri (1956) established 107 Drosophila
populations.
All founders were heterozygotes for an
eye-color gene called brown. Neither
allele gives selective advantage.
Initial genotype bw75/bw
Initial frequency of bw75 = 0.5
Followed populations for 19 generations.
Population size kept at 16 individuals.
What do we predict will occur in terms of
allele fixation and heterozygosity?
In each population expect one of the
two alleles to drift to fixation.
Expect heterozygosity to decline in
populations as allele fixation
approaches.
Distribution of frequencies of bw75 allele
became increasingly U-shaped over
time.
By end of experiment, bw75 allele fixed in
28 populations and lost from 30.
Fig 6.16
Frequency of heterozygotes declined
steadily over course of experiment.
Declined faster than expected because
effective population size was smaller
than initial size of 16 (effective refers to
number of actual breeders; some flies
died, some did not get to mate).
Fig 6.17
Effective population size refers to the
number of breeders in a population.
In many cases population size and
effective population size may be quite
different if most individuals don’t breed, if
a few individuals produce most of the
offspring or if there is a strongly biased
sex ratio.
Templeton et al. (1990) Studied Collared
Lizards in Ozarks of Missouri
Desert species occurs on remnant pieces
of desert-like habitat called glades.
http://en.wikipedia.org/wiki/F
ile:Collared-lizard.jpg
Human fire suppression has resulted in
loss of glade habitat and loss of
crossable savannah habitat between
glades. Areas between glades
overgrown with trees.
Based on small population sizes and
isolation of collared lizard populations
Templeton et al. (1990) predicted strong
effect of genetic drift on population
genetics.
Expected low genetic diversity within
populations, but high diversity between
populations.
Found expected pattern. Genotype
fixation common within populations and
different genotypes were fixed in different
populations.
Lack of genetic diversity leaves populations
vulnerable to extinction.
Found >66% of glades contained no lizards.
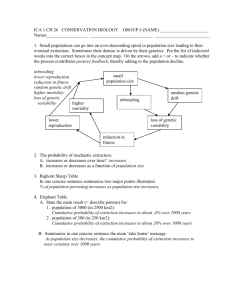
What conservation measures could be
taken to assist Collared Lizard
populations?
Repopulate glades by introducing
lizards.
Burn oak-hickory forest between glades
to allow migration between glades.
Another way in which populations may
be exposed to the effects of drift is if the
population experiences a bottleneck.
A bottleneck occurs when a population
is reduced to a few individuals and
subsequently expands. Even though the
population is large it may not be
genetically diverse as few alleles passed
through the bottleneck.
Simulation models show a bottleneck
can dramatically affect population
genetics.
Next slide shows effects of a bottleneck
on allele frequencies in 10 replicate
populations.
The northern elephant seal (which
breeds on California and Baja California)
was hunted almost to extinction in the
19th century. Only about 10-20
individuals survived.
Now there are more than 100,000
individuals.
The northern elephant seal population
should show evidence of the bottleneck.
Two studies in the 1970’s and 1990’s that
examined 62 different proteins for
evidence of heterozygosity found zero
variation.
In contrast, similar studies of southern
elephant seals show plenty of variation.
More recent work that has used DNA
sequencing has shown some variation in
northern seals, but still much less than in
southern elephant seals.
Examination of museum specimens
collected before the bottleneck have
shown much more variation in these
specimens than in current populations,
which shows that the population was
much more genetically diverse before
the bottleneck.
In small populations inbreeding may be
unavoidable.
Even with random mating, a small
population that stays small and receives no
immigrants will become inbred.
Major problem for rare species such as
California sea otters and northern elephant
seals.
Two hundred years ago Illinois covered
with prairie and home to millions of
Greater Prairie Chickens.
Steel plough allowed farmers to farm the
prairie. Acreage of prairie plummeted
and so did Prairie Chicken numbers.
Lesser Prairie Chicken
In 1960’s habitat protection measures
introduced and population increased
until mid 1970’s.
Then population collapsed. By 1994 <50
birds in two populations in Illinois.
Fig 6.3
Why did prairie chicken populations
decline even though available habitat
was increasing?
Prairie destruction reduced numbers of
birds and isolated the populations from
each other.
No migration between populations.
Small populations vulnerable to genetic drift
and inbreeding depression.
Accumulation of deleterious recessive
alleles (genetic load) can lead to extinction
of small populations.
Problem exacerbated when exposure of
deleterious mutations further reduces
population size and increases
effectiveness of drift. “Extinction vortex”.
Prairie chickens showed clear evidence
of inbreeding depression. Egg hatching
rates had declined dramatically by 1990
< 40% hatch rate.
FIG 6.31
Illinois Prairie chicken populations showed
less genetic diversity than other populations
and less genetic diversity than they had in
the past.
Illinois birds 3.67 alleles per locus rather than
5.33-5.83 alleles of other populations and
5.12 of Illinois museum specimens.
Conservation strategy?
In 1992 prairie chickens introduced from
other populations to increase genetic
diversity.
Hatching rates increased to >90%.
Population increased.