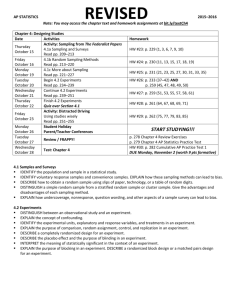
Document 12361897
advertisement

Project acronym: MIDAS Grant Agreement: 603418 Deliverable number: Deliverable 4.2 Deliverable title: Gap analyses of existing data to determine what future sampling is required and to provide support for ecological modelling from selected study regions Work Package: WP4 Date of completion: 14 December 2014 BIOGEOGRAPHY AND CONNECTIVITY IN DEEP-SEA HABITATS WITH MINERAL RESOURCE POTENTIAL A GAP ANALYSIS Table of Contents Executive Summary 1 3.1.5 CCFZ: Nodules 24 Current state of knowledge 3 3.1.6 CCFZ: Gaps to our knowledge 26 1. Introduction 7 KNOWLEDGE GRID for CCFZ 28 1.1 Aims and objectives of study 7 3.2 SMS – MIDAS areas from the Mid Atlantic Ridge 29 1.2 Study areas 7 3.2.1 Patterns of biogeography 30 1.3 Methodology approach undertaken in this study. 7 3.2.2 MAR-Vent systems: Gaps to our knowledge 30 Knowledge Grids 8 3.3 Non-active vents and non-vent hard substratum sites 31 2. General considerations - rarity 9 3.3.1 Current Knowledge: Biogeography of major faunal groups 31 2.1 Rarity in deep-sea samples 9 KNOWLEDGE GRID for MID ATLANTIC RIDGE - Vent systems 32 2.2 The rarity-endemicity issue 9 3.3.2 MAR – non-vent and off vent environments: Gaps to knowledge 33 2.3 Rarity, sampling and appropriate scales of study 9 Cnidarian megafauna–Case study 3 34 2.4 How to deal with rare species in a mining environment? 9 KNOWLEDGE GRID for MID-ATLANTIC RIDGE - Non-vent environments 35 2.5 Discussion and Conclusions 13 3.4 Methane hydrate – Svalbard margin and Black Sea 36 Rarity and deep-sea sampling: Case study 1: 14 3.4.1 Svalbard margin. 36 3. Study areas: current biogeographic knowledge and gaps 17 KNOWLEDGE GRID for SVALBARD MARGIN 37 3.1 CCFZ – sediment nodule fields 17 3.4.2 Black Sea 38 3.1.2 Physical and biogeochemical gradients in the CCFZ 17 4. Modelling 39 3.1.3 Scales of habitat heterogeneity 18 5. General discussion 39 3.1.4 Molecular approaches 18 6. Conclusions 40 3.1.5 Patterns of biodiversity in the CCFZ 20 6. References 41 Macrofauna – Case Study 2 21 MIDAS: WorkPackage 4– deliverable 4.2. This report should be quoted as: Paterson,G.L.J., Menot, L., Colaço, A. Glover, A.G., Gollner, S., Kaiser, S., Gebruk, A.V., Janssen, A., Silva, M. C. A., Janssen, F., Sahling, H., Felden, J., Martinez, P.A. 2014. Biogeography and connectivity in deep-sea habitats with mineral resource potential–gap analysis. Deliverable 4.2. MIDAS. 45pp. 12 December 2014 BIOGEOGRAPHY AND CONNECTIVITY IN DEEP-SEA HABITATS WITH MINERAL RESOURCE POTENTIAL: A GAP ANALYSIS Executive Summary Determining the environmental impact of mineral extraction depends on the knowledge, information and data available. The deep sea remains our least explored and also our largest environment on the planet. It follows that a considerable level of knowledge will be required to assess and manage sustainably exploitation of resources found in the deep sea. This report is an investigation identifying the fundamental gaps in our understanding of the distribution of the organisms which inhabit the various deep-sea habitats currently being explored as potential sites for mineral extraction. Understanding the biogeography of deep sea biota, the spatial distribution of life, is an essential first step in assessing the risks that extraction will pose. Knowing the distribution of an organism is at the heart of assessing the likelihood that human activities will ultimately lead to its extinction. In this report we undertake a gap analysis of current biogeographic knowledge of four sites where deep-sea mining may take place: massive sulphide deposits on the Mid-Atlantic Ridge; nodule fields in the Central Equatorial Pacific, methane hydrate fields in the areas off Svalbard in the Arctic Ocean and in the Black Sea. Gap analyses in this context is taken to mean an assessment of the current understanding of biogeography, what data are available to assess distribution patterns and what information needs to be gathered to address any gaps. The study focuses on the benthic fauna, looking at the various size groups and the information known. The pelagic system is not dealt with. We present three case studies which highlight different issues related to the biogeography of deep-sea organisms. The first deals with rarity, the widespread observation that a sizable proposition of the fauna collected is represented by species with only one or two individuals. An analysis indicates that while increased sampling can provide greater information on whether species are really rare or merely undersampled, practically no amount of sampling will be able to solve this riddle. Ultimately it will be for regulators to decide whether the risk of extinction of rare species is acceptable. The second case study attempts to use existing data to see if it is possible to determine whether the fauna associated with the nodules fields of the Pacific is widespread or changes with distance. The initial results based on macrofaunal polychaetes and meiofaunal nematodes indicated that the species assemblage changes with distance. If this is supported by further study then it would mean that extensive mining in one area may well pose a higher risk to the species which occur. It is therefore important to know the local distribution of species. However, the case study also indicated that the high numbers of rare species may bias the results so further study and sampling is needed. The third case study looks at what is known about epifaunal cnidarian species which are widespread but occur at low densities. One of the species is the octocoral Corallium, which occurs on non active and non-vent hard substratum sites. It is important that we gain a better understanding of the underlying factors governing epifaunal distribution in order to assess whether the search for and exploitation of one economic important resource could impact on another. Understanding the species distributions of this group would also provide information on what conditions are required for epifaunal species to settle and grow. General conclusions Spatial coverage of sampling programmes. The range of scientific, i.e. research-led, sampling programmes to the various areas differs quite markedly. There is little known about the biota of the methane hydrate fields off Svalbard, while other sites have been subject to varying degrees of study. Perhaps the best investigated are the MAR vent sites TAG – most visited, Lucky Strike – best studied and both have been visited on a fairly regular basis since their discovery in the late 1980s. The recurrent observation with all areas is that we lack good data on the distribution of biota on many scales from local (i.e. within habitats), regional (between habitats found within a site or claim area) and on a basin scale. Stratified, co-ordinated sampling programmes will be needed to address this gap. The road map suggested by German et al. (2011) highlights the need for academia and industry to continue to work together to address key issues such as extinction of rare species and potentially of isolating populations. Such collaborations will go some way towards addressing the gaps highlighted. Taxonomic coverage and resolution of different biota. Taxonomy is the key to understanding the distribution and biogeography of biota. Megafauna generally have been better studied and their taxonomy is at a more advance stage than for most of the other groups. Nevertheless, many megafaunal species, even well known ones, lack any molecular assessment. In general population studies aimed at determining connectivity have only been attempted with abundant species but for most species their populations are too small and widely dispersed to be able to collect enough individuals to undertake meaningful population genetic surveys. Modern genetic methods may be able to address this problem but such methods have not been deployed in current surveys. 1 Taxonomic resources, specifically trained people, continue to be a limiting factor despite the rise of molecular tools. The reasons for this have been the subject of much debate and will not be repeated here but within the deep sea one reason is the shear scale of the species richness that sampling reveals. For smaller macrofauna, meiofauna and taxa, such as Foraminifera, the numbers of species recovered from a sampling programme can be into the hundreds or thousands. In the past this has led to a more ecologically focused approach which has concentrated more on the identification of samples and much less on the classification and formal description of species. The result has been that formal taxonomy lags behind discovery and ecological analyses, creating a major gap in our biogeographic knowledge of deep-sea biota. And now, as we stand on the brink of commercial exploitation of deep-sea resources, we cannot say with any degree of certainty what the risk of extinction will be for even large charismatic megafaunal organisms. Table 1. RAG assessment of biogeographic knolwedge of MIDAS sites being explored for mineral respources. Vents Megafauna For some areas there is a considerable amount of data available in the scientific literature. Vents megafauna being the most studied. However, despite over four decades of research in some of the MIDAS study areas the data remain patchy and scarce. The Internatonal Seabed Authority have developed a central database for collating information which will serve as a much needed source of data for future studies. At all times strong data management will be 2 Svalbard seeps Black Sea N/a Meiofauna ? Microbial: Protista Microbial: Bacteria Microbial: Archaea Good knowledge with ability to make informed predictions s CCFZ N/a Macrofauna To reverse this both academic and commercial programmes need to collaborate and develop robust taxonomic pipelines which will result in consistent identification of species. Such an infrastructure will need to include both traditional morphological assessments as well as the application of genetic tools and will need to use web-based approaches to disseminate this information in a timely way. Decades of work in various different habitats has shown that without adequate resources to establish and run taxonomic pipelines, no further work on the taxonomy of species will be carried out. The critical data needed to assess risk of extinction will not be generated. Data availability. Off vents Useful data available but fundamental gaps remain Little or primary information only essential to ensure that data being produced are appropriate and fit for purpose. Collections of specimens from previous studies, both commercial and academic are scattered across a variety of institutions. It can be difficult to determine where the material is. Some form of metadata catalogue would assist in keeping track of important collections. As commercial exploration activities increase in intensity, good protocols will be needed to ensure that samples/specimens are stored in recognised collection-based institutions and made available for study. Again resourcing for this activity needs to be built into the commercial and academic programmes. Specimens are the only way to verify the presence of a species at a particular place in space and time and therefore are an important part of the environmental auditing process. Current state of knowledge difficult to make convincing predictions as to the impact of mining on species. The RAG assessment (Red, Amber and Green) is based on analyses of current knowledge of the biogeography of different organism groups and the MIDAS sites (Table 1). It is meant as a quick assessment of the current state of understanding. Green status means that the taxonomic knowledge is robust and that it is possible or could be possible to make informed predictions as to the impact of mining. Red status means that the only sources of data and information are in primary scientific publications and that the coverage of the investigations undertaken are limited. It is not possible to make any realistic predictions on the impact of mining. Only the megafauna associated with vent sites have been well enough studied for an informed assessment of their biogeography which could be used to assess mining impacts on species populations. Most of the other faunal groups and areas require considerably more research before we are able to make informed assessments of impacts. Amber status indicates that study of the particular taxonomic group at a particular site is more comprehensive, more than just primary data are available. But there are significant gaps in biogeographic knowledge such that it is 3 BIOGEOGRAPHIC KNOWLEDGE GAPS WHICH IMPACT ON ENVIRONMENTAL ASSESSMENT AND SUSTAINABLE MANAGEMENT 1) Spatial coverage of sampling programmes. Gap Mitigation Actions Priority Information is insufficient for comprehensive analyses of most taxa likely to be affected by mineral extraction irrespective of site. Focused biological cruises coupled with stratified sampling programme across a range of spatial scales. Better co-ordination between teams undertaking exploratory work and research projects. Coverage is patchy dependant on site and taxon. Emphasis on obtaining a better taxon sampling using a range of gears. High to Medium Insufficient information or data to determine linkages between different habitats, such as sea mounts and other hard rock surfaces. Sampling across different habitats not just those of immediate economic interest. Medium In large habitats such as abyssal plains and MAR off vent and non-vent environments, too little is known of the pattern of distribution at different spatial scales which is the first step towards identifying the underlying ecological drivers . Targeted research including multidisciplinary cruises comparing Massive Sulphide Deposits (MSD) with surrounding hard substratum habitats. High to Medium In areas where mining claims have already been established – MAR and CCFZ – there is little incentive for the different contractors to co-ordinate and share data, although some now do. Regulatory bodies should play a more constructive role in co-ordinating between contractors and establish protocols for the sharing of non-commercial scientific data between contractors, academia and NGOs. High to Medium High 2) Taxonomic coverage and resolution of different faunistic elements. Taxonomic knowledge needs to be developed as in only a few groups is there sufficient biogeographic information. Ensure taxonomic standards across all actors, particularly contractors, though workshops, create a taxonomic data portal, encouraging publication of taxonomic results, develop a taxonomic clearing mechanism which links taxonomic resources with teams needing specialist support. High Molecular approaches linked to morphology and morphotypes need to be applied. Molecular approaches need to be built into the taxonomic data pipeline. Linking molecular data to morphotypes, where applicable, will enable previous studies and the taxa collected to be linked more unequivocally to future identifications. High Developing infrastructure such as a clearing mechanism and a data sharing portal will support taxonomic efforts and maximise existing resources. High to medium Resolving OTU identifications, particularly intercalibrating between different studies and between species identified using molecular methods will maximise the taxonomic resources and improve resolution. High Better co-ordination between taxonomists, ecologists and the contractors is needed to maximise existing taxonomic resources. Resolution of most smaller elements of the biota, including the microbial, is at the level of (M)OTU. This is a major impediment to understanding the distribution of biota. 4 3) Data availability Gap Mitigation Actions Priority Considerable amounts of data may be ‘hidden’ within grey literature – contract reports or unpublished surveys. Such data are effectively unavailable. Create a data portal to release non-commercial scientific data. The ISA would be the obvious organisation to undertake this for the areas with mining claims. Medium Specimens, here considered the primary element of biogeographic data, are also scattered, often difficult to access. A metadata catalogue or holdings indicating what material is held where and what its status is in terms of identification would be a useful resource. Medium Intercalibration of specimens is difficult and there is no mechanism or incentive for contractors to join with researchers to produced unified datasets. Develop appropriate taxonomic infrastructure as outlined above. Fund taxonomic intercalibration workshops to produce unified datasets and publishable taxonomic keys and descriptions. High to medium MIDAS sites Clarion–Clipperton Fracture Zone nodule fields CCFZ: High resolution studies to determine spatial patterns of species distributions. It is important to understand how species are distributed across different habitats within claim areas, between claim areas and in the APEIs. Focused biological cruises by contractors and the research community. Co-ordinated exchange of information. Ensure the use of the ISA database. High CCFZ: Lack of basin-wide species level datasets. Encourage contractors to make non-sensitive data available via ISA. High CCFZ: Lack of taxonomic consistency. 1) Create taxonomic clearing system and centralise taxonomic depository. 2) Training . 3) Taxonomic exchange programmes to improve capacity among actors. High CCFZ: Nodule fauna–is it endemic to nodules or found on other metal rich habitats such as seamount crusts? Sample seamounts within and beyond CCFZ. High CCFZ: Epifauna found in the CCFZ– does it also occur on hard surface areas such as seamounts? Focused sampling of megafaunal groups, particularly for molecular analyses. Develop cruise programmes to visit both types of habitat. Many claim areas also have a range of seamounts so targeted cruises could be planned.. High CCFZ: Virtually nothing is known about protistan taxa such as ciliates, flagellates etc. The importance of this obvious gap in knowledge is also unknown. Molecular eDNA may be the only way to determine the biodiversity of such groups. Determining their role in the ecosystem is also problematic and may depend on molecular approaches. High to medium CCFZ: Microbial diversity and distribution. Considerably more sampling is needed to understand the distribution of bacteria and archaea across the CCFZ. Their function both in nutrient and metal cycling is important and needs to be elucidated. High to medium CCFZ: Identifying the drivers for biogeographic patterns in the CCFZ Integrating ecological data, e.g. productivity, with phylogenetic information from key species to identify the historical-evolutionary processes and the ecological drivers. High to medium 5 MAR: Hydrothermal vents sites Gap Mitigation Actions Priority MAR: There have been few studies focussed on meiofauna diversity and distribution – mostly in the Pacific and some from reduced sediments. Targeted sampling – within mussel beds and tube thickets. High to medium MAR: Little is known about microbial protistan biodiversity. Sampling hard surfaces is difficult and new collectors may need to be engineered to optimise collection of these taxa. Targeted sampling across various habitats and zones within and between vents. High to medium MAR: Increase molecular taxonomic knowledge and population studies of all taxa. Establishing standard sampling and processing protocols to be used by academic research and contractors. High MAR: Greater sampling coverage along the MAR and within the claim areas is needed to be able to establish biogeographic patterns and distributions at different scales. Co-ordinate research and exploration programmes and encourage academic and contractor collaboration. High MAR: Off-vent and non-vent areas MAR: There is no coherent understanding of the distribution of fauna from the sediment habitats away from active vents and how it relates to sediment fauna from surrounding abyssal and continetal habitats. Stratified sampling combining AUV, ROV and tethered samplers from the claim areas and from areas outside on the MAR High MAR: There is little understanding of the ecological drivers of the biogeographic patterns of species associate with both hard rock and sediment habitats. Targeted multidisciplinary research projects focused on off-vent sites, bringing together academia and contractors. High to medium MAR: molecular data is not available for most mega, macro and meiofauna which impedes the ability to determine connectivity and distribution. Establish molecular sampling/subsampling as a standard protocol for future work. High Svalbard: Methane hydrates Svalbard: Intiatial investigations point to similarities between the Svalbard sites and those from the Håkon Mosby mud volcano and Norwegian margin. However, different chemosynthetic sites often have unique combinations of species so more intensive study is needed to detemine the relationship and distribution patterns of the fauna. 6 Comprehesive sampling, including for molecular analyses will provide Medium 1. Introduction This study assesses the current state of the art in relation to our knowledge of the distribution and biogeography of the fauna from areas rich in deep-sea mineral resources. The aim is to identify gaps, suggest future research priorities and to inform monitoring strategies being developed for these areas. 1.1 Aims and objectives of study Knowledge of the distribution of organisms within and between the sites of potential commercial interest and the surrounding region is crucial in developing management plans. Without such knowledge it becomes almost impossible to determine the impact of resource extraction on a regional scale. In effect how likely is it that such activities lead to the permanent extinction of species? 2) Taxonomic coverage and resolution of different faunistic elements. The aim is to determine whether all the ecological actors had been sufficiently analysed. This aspect also included whether there were any genetic analyses. Here we assess whether taxonomic coverage and resolution is sufficient to analyse biogeographic patterns and 1.2 Study areas The main areas under consideration are the Mid-Atlantic ridge hydrothermal systems , the Clipperton–Clarion Fracture Zone polymetallic nodule fields in the Central Pacific, methane hydrate fields in the Arctic and Black Sea. All of these areas are likely to be regions where commercial exploitation of mineral resources is planned. Figures 1 and 2 show the study sites. 1.3 Methodology approach undertaken in this study. In this study we employ the term gap analyses to mean an assessment of the fundamental data need to provide information for management such that there is a good understanding of the distribution of species and what impact extraction of resources would have on species affected. The information required could be both factual as gained by analyses of samples or based on probabilistic models together with the appropriate error terms. The analyses undertaken followed the following steps: 1) Spatial coverage of sampling programmes in relation to the key areas under investigation. Here we assess gaps in the geographic coverage what has been termed the Hutchinsonian perspective Figure 1. MIDAS Study areas. Areas of particular focus for WP4: 14) CCFZ - nodules; 9-12) Mid-Atlantic Ridge - ; 1,2,5) Black Sea and N. Arctic – methane hydrates. 7 connectivity between populations. This has been called the Linnean perspective 3) Data availability. Given that in many of the study areas research is undertaken by different teams from different institutions and in some areas, such as the CCFZ, work is undertaken by different contractors we assess how easy it is to access data being produced. Assessing data coverage is a necessary first step in developing predictive models. Modelling is seen as a way of dealing with the large spatial scales present in some of the sites and the problems of adequately sampling in such an environment. Knowledge Grids As a means of summarising the understanding and availability of data, a knowledge grid has been developed for each of the main environments dealt with in this report. The grid is composed of different taxonomic groups as columns across the top–megafauna, macrofauna, etc.; with a series of subject categories as rows. The categories assessed were as follows: Taxonomy Taxonomic knowledge: this is an assessment as to what level of taxonomic information is available to undertake biogeographic studies. Taxonomic resolution: this is a measure of the taxonomic maturity. Are the taxa well classified and described? Or are they only classified as an Operational or Molecular Taxonomic Unit (OTU – MOTU), i.e. species 1, species b, etc. Keys: in particular whether the data are available in original primary sources or whether there are keys and large revisions or monographs available. Taxonomic information availability: in what form is the taxonomic information available–primary sources, scientific literature, on-line and what mechanisms, other than publication in the scientific literature, will help people exchange taxonomic information. Sampling data coverage Standards and protocols: are samples being taken using similar methods between the different teams undertaking surveys and primary research studies? Biogeography (in this study it is ecological aspects of biogeography which are the focus) Information and data availability: are there current, available biogeographic analyses with suitable data already available? Or if not are there any data with which to undertake relevant biogeographic analyses? The study focuses on the benthic fauna, looking at the various size groups and the information known. The pelagic system is not dealt with in this study. Sampling: what kind of sampling is currently generating taxon-based data? Sample coverage: how comprehensive are the sampling programmes – any particular gaps? Molecular sampling: are samples for molecular analyses being collected? Is this linked to morphological taxonomic analyses or purely molecular such as eDNA studies? Figure 2. MIDAS WP4 sites: CCFZ nodule fieds (image IFREMER); Hydrothermal chimney at Rainbow vent, MAR (image IMAR/DOP) ; MAR non-ventcoral garden Menez Gwen (image IMAR/DOP); Black Sea bacteria carbonate mounds (image AWI); Arctic methane seeps (image AWI). 8 2. General considerations - rarity In assessing biogeographic patterns, dealing with the question of rarity becomes a major challenge; one which is seldom really dealt with in the deep-sea literature. The long tail of rare species associated with samples is considered to be an emergent property of the deep-sea ecosystem. However, in trying to determine ecological impacts some regard as to this challenge has to be made. Effectively, the question will be asked – What is the likelihood that exploitation of mineral resources will lead to the extinction of species and that those species will be the rare or least abundant ones? Or is this rarity really an artefact due in part to the huge size of the ecosystem and our ability to sample it effectively? 2.1 Rarity in deep-sea samples In the following sections we discuss the implications of rarity on assessing exploitation of resources. Rarity and what it might mean in terms of biogeographic analyses and subsequent environmental management are discussed. 2.2 The rarity-endemicity issue A major characteristic of many species-rich habitats is that the assemblage comprises a large number of species represented by one or two individuals. In the deep sea benthic samples are characterised by a long ‘tail’ of such species (see Fig.2). In a study of hydrozoan and anthozoan ‘coral’ species found on the hard surface habitats around the Azores, Braga-Henriques et al. (2013) reported that 30% of the 164 species recorded had only one or two records (Fig 2). Collins et al. (2012) reported that 80% of the fauna recovered from the vent systems in the Manus Basin off Papua New Guinea were represented by 5 or fewer individuals. This long tail of species with few individuals is a major characteristic of deep-sea diversity. However in terms of assessing biogeography and connectivity the potential rarity of such species posses a major question in such studies - are such species actually rare or just undercollected? This is the ‘rarity’ problem (Gaston, 1994). Rarity in this context is not the same concept as endemism. Rare species can be widespread and endemic species can be abundant. The problems and challenges to defining rarity involve sampling intensity and variability in abundance – the higher the errors in measuring these factors the less confidence the rarity estimates return. 2.3 Rarity, sampling and appropriate scales of study Sampling efficiency is a particular issue with deep-sea samples. In sediment environments such as the CCFZ, quantitative sampling devices, such as box corers, sample a small area of the ocean floor and as a result large numbers of replicates are needed to adequately sample one area. The intensity of the sampling required places a heavy burden on resources which is accentuated in some deep-sea settings where faunal abundance is very low. Use of the new generation of epibenthic sledges can collect considerably larger samples. They do not necessarily collect the same elements of the fauna compared with corers. Such samplers are also qualitative. Furthermore, in nodule areas, the material collected can be quite damaged. So though no apparatus will be representative for the entire fauna, a combination of samplers will probably provide more robust estimates of biodiversity. Rock environments such as MSD also are problematic in terms of survey and sampling. Extensive phototransects are available for the vent areas themselves but off-vent areas are less frequently surveyed. Collecting specimens requires intensive use of ROVs. It is, therefore, difficult to determine whether hard substrate fauna is rare or merely undersampled. This challenge was also noted by Barsa Henriques et al. (2013) for coral species around the Azores. They suggested that it was not yet possible to be certain that records of rare species were not artefacts of sampling despite 150 years of sampling and study; increasing the sampling area may well result in the discovery of more specimens of such species. Molecular approaches provide an opportunity to shed some light on connectivity between low abundance populations which may be considered rare. But even here if sufficient material is not collected then establishing rates of genetic exchange remains difficult if not impossible to determine. 2.4 How to deal with rare species in a mining environment? Rarity definitions are often based on a different scales, e.g. definitions such as 'locally common but globally restricted' are often used but must take into account skewed observations and temporal change. Species life cycles can lead to changes in population sizes. A more pragmatic approach may be to specify the actual area of habitat being used or could be used by the species rather than some arbitrary area encapsulating a range of habitats i.e. drawing a line round a geographic region. Modelling may support such attempts but more information on potential life styles, environmental variables and biogeochemical settings will be required (see Section 4. Modelling). These characterisations are difficult in the deep sea. As a first step it is possible to assess the potential distributions of species and therefore their status based on their occurrence in samples and the degree of sampling undertaken. 9 Figure 2a. A typical species abundance graph showing the long tail of macrofauna species with low numbers of individuals from the Domes A nodule site in the Clipperton-Clarion Fracture Zone, Central Pacific. Figure 2b. Nematode species rank abundance curve from the equatorial site of the EqPac study. Data from Brown (1998). Figure 2c. Records of Anthozoa recorded from around the Azores EEZ. The records are based on assessing literature records, fishing bycatch returns and new sampling. In this example the number of records indicate the rarity of the species such that 50 species have only been recorded from one sample. From Barga Henriques et al. (2013) Figure 2d. Records of holothurian and asteroid species recorded from the Mid-Atlantic Ridge. These records are based on samples taken during the MAR-ECO programme. Data from Gebruk (2008) and Dilman (2008). 10 Draft 11 Nov 2014 Table 2. Why it matters whether a species is rare. The impact of mineral extraction is compared against the different levels of species abundance. The consequences of species loss at different geographical scales are estimated. Species occurrence Local scales (i.e. claim area, vent site) Regional Scale Global scale Common Local extinction but populations elsewhere remain viable Some disruption to gene flow but no major losses No major losses Locally abundant and widespread As above As above but local regional extinction may occur changing the biogeography of the species No major losses Locally abundant widespread but rare in other localities1 Local extinction but recovery and population viability depends on where the species is most abundant Depending on population impacted the effect may be to push a species to non-viable levels Potential loss or reducing the population of species to low levels. Potentially placing species at risk. Locally rare but widespread Local extinction likely Regional extinction may occur changing biogeography Globally rare may result in local but not total extinction Local widespread, rare and restricted –endemic species Local patches may survive if widely dispersed within and beyond exploitation area Threat of total extinction. At regional scales it is a question of determining habitat preferences and distribution patterns Species extinction possible. Rare species Locally restricted could result in total extinction Extinction possible Extinction possible This is important in the context of local/claim area management. There may be regions of high richness with localised species which need to be preserved. Detailed sampling in the area is needed 1 11 2.5 Discussion and Conclusions There are two aspects to rarity which need to be considered. The first is whether extinction is a real possibility and therefore acceptable in terms of the sustainable management of mineral extraction. The second is that if it is deemed acceptable what potential impact could this have on ecosystem functioning. Even with advances in molecular approaches or applying modelling techniques, the rarity issue remains problematic and it perhaps best categorised as potentially unknowable. Pragmatically, given that research may be unable to resolve the issue of whether extinction will be caused, it becomes a matter for regulators to evaluate the risk. 12 Functionally what do such rare species contribute to ecosystem function? Given they are found in low abundance it is tempting to conclude that their contribution is negligible. This however may be inaccurate and a result of studying their distribution at an inappropriate scale (Jain, 2013; Mouillot et al 2013). The long tail of species with single occurrences may appear to contribute little apart from contributing to the high species richness characteristic of a deep-sea sample. However, the large number of such species may confer a degree of redundancy to the system (Naeem, 1998). How a reduction in the number of species would impact on the system's ability to recover or species to recolonize remains unknown. In other ecosystems rare species have been shown to have functional traits not found in the common species and therefore contribute to the functional diversity of the ecosystem. In some cases rare species may support highly vulnerable functions. Mouillot et al. (2013) suggest that such species may provide the ecosystem with the functional robustness to adapt to climate change or anthropogenic pressure. Table 3 summarises the discussions around rarity and how it relates to impact assessment and potential consequences following exploitation. Table 3. Rarity and its relevance at different scales following anthropogenic distrubance Locally rare Basin-scale rare Globally rare Local extinction threat – increase in population patchiness Loss of genetic diversity – potential impact on functioning and gene flow Serious threat of extinction Detailed sampling across the claim and preservation areas is essential to be able to determine whether refugia contain functioning populations Need to know the distribution of all size class elements at regional scales Difficult to establish as the likely distribution and occurrence are often unknown due to lack of appropriate sampling Knowledge Gap Insufficient knowledge of small-scale distributions and how this is related to scales of physical heterogeneity Insufficient molecular data to establish population genetics Insufficient information of biogeography and connectivity basin scale studies Understanding of species distributions and population connectivity is poor to non-existent Lack of synthetic studies bringing together information on distribution of different taxa Reasons for gap Detailed sampling is resource heavy There is no regulatory mandate to do this type of study Taxonomic and ecological expertise is in short supply Funding is not sufficient to support this intensive approach Concentration by different contractors of groups in selected areas Lack of exchange of data Molecular data are needed but often difficult to get for certain faunal elements Lack of data Large-scale studies, particularly taxonomic ones are time-consuming and require financial support Need taxonomic support and co-ordination Consequences Unable to determine whether refugia or set aside areas will provide sufficient protection Unable to determine basin scale impacts and to determine efficacy of regions of special interest. Unable to determine extinction levels on a global scale because of poor baseline data Local areas of refuge and no activity areas Regional reserved areas and no activity zones Restrictions on activity in zone where endemic species are found Impact Relevance to deep sea Current mitigation proposals 13 Rarity and deep-sea sampling—Case Study 1 An analysis of existing data is undertaken to assess categories of species distribution. The aim is to determine: 1) how sampling may affect the designation of rarity to species occurrence; 2) a more robust definition of rarity and its consequences if an area is likely to be exploited; 3) what steps might be take to reduce the impact of exploitation on such species. Table 1 summarises the impact that intensive exploitation would have on species with differing levels of occurrence at different scales. An attempt is made to provide a more rigorous definition of species occurrence to try to separate out different consequences of exploitation on their survival at different spatial scales. Method Four datasets from the CCFZ were studied – two datasets from sites east to west across the CCFZ (macrofaunal polychaetes) and two from sites south to north (one of polychaetes and one of meiofaunal nematodes). The east–west samples were from previous studies: Domes A, Echo and PRA, here termed CCFZ 1, and from three sites sampled during the Kaplan study (ISA technical Report 3, 2008), here termed Kaplan, in the east, centre and west (this latter site is very close to Domes A).South to North sites were from the EqPac programme were assessed and the numbers of species in the categories above were counted. Each species was assessed as to its occurrence in the samples from each site, whether it also occurred in different sites and if so how many. The relative abundance of each species was also assessed to provide some indication of whether the species occurred in the same relative density in all samples and sites. Based on its occurrence each species was classified according to the terms defined below. To assess whether sampling had any effect on the numbers of rare species, the numbers of species recorded with just one individual were counted in the individual samples, in the pooled samples for the whole site and pooled across 14 the whole study area. The results were then graphed against the number of samples. localities are seriously undersampled to address the issue of rarity and endemicity in the abyss. Definition of terms Taxonomy is also a potentially confounding factor, even when the study was conducted by one group or taxonomist. As a result it is not possible to amalgamate the CCFZ 1 identifications with those from Kaplan as different taxonomic teams carried out the identifications and there has not been the opportunity to undertake an intercalibration exercise. Damaged specimens and the sheer number of species can make it difficult to maintain a consistent taxonomy across many samples. To try to mitigate against this, a conservative approach has been taken in grouping the occurrence categories. Common: Found in many samples across a site in relatively high numbers (at least >5% in some samples) and also found in all sites studied with similar site-relative abundance. Locally abundant and widespread: Found in some samples within more than one site in abundance excess of 5% and also found in other geographically separate sites. Locally abundant widespread but rare in other localities: Found in relatively high abundance (>5%) in samples at one site but is also found in other sites but in much lower abundance and maybe locally rare. Locally rare but widespread: found in low numbers at any one site but found in a number of geographically separate sites. Local widespread, rare and restricted: Found in a number of samples across a site but in low abundance; not found outside the site. Rare: found only as single individuals in one or two samples within a single site. The relatively low number used for the percentage abundance is a reflection of the generally low abundances found in deep-sea systems. The exception being high productivity areas associated with vents. Caveats Domes A samples were potentially compromised by poor sorting and represent an impoverished estimate which might account for the very low numbers recorded in this site. The numbers of samples taken has a bearing on species discovered but it is not a linear relationship so it is difficult to be sure when the collectors curve would flatten out. Domes A samples (47 Spade Box Cores) when analysed indicated that between 30 samples would be sufficient (Paterson et al. 1997). This means that the other Results Effect of sampling There appears no obvious relationship between sampling intensity and the number of species defined as rare (Fig. 3a). Restricting the analyses by removing the Domes A data still shows that as expected there is high variation at the single sample level which drops as more samples are pooled (Fig. 3c). However, the current data suggest that both the mean number and relative percentage of rare species remains relatively high even when as many as 25 samples are pooled. CCFZ. Rare species with only one occurrence in any sample represented 48% of the total CCFZ fauna and if samples with only occurring locally (potential endemics?) species are included the proportion of the fauna restricted rises to 55% (Table 4). However it is interesting is that many of the species found are locally rare but are found in sites across the CCFZ sometimes they may be locally abundant (42.8% of the total number of species). So this implies that such species are in fact very widespread in occurrence and occur at low densities. Increasing sampling may detect more of such species but there is a diminishing return on this in terms of effort. A similar pattern can be seen in the EqPAC samples where rare and locally endemic species comprise approximately 65% of the fauna. Similarly the widespread species occurring in low numbers equate to approximately 34%. Rarity appears to be a conspicuous and consistent feature of the fauna of sediment habitats. Intensive sampling points towards the discovery that single occurrence species may actually be rare but have a wide distribution either in the local area or across a wider spatial extent. Despite this there remains a significant proportion of species whose occurrence is below the threshold of possible sampling intensity. It is highly unlikely that such species are the sole representatives remaining of their species given the high numbers which are consistently found in samples. Such a conclusion would imply either that that the deep sea is populated, indeed dominated, by species on the very brink of extinction; or that the abyssal fauna represents a sink and that the viable centre for the species is outside the abyss (Rex et al. 2005). However, with our current knowledge we are unable to determine the likely distributions of such species. Based on current data, the densities in the CCFZ of such rare species would be below 1 individual per 12.25 m2 based on samples taken across distances of over 2700 kilometres. The nematode data showed that there was a gradual reduction of rare species as samples are pooled and a greater area is sampled (Fig. 3d). Magurran & Henderson (2003) consider that there are actually two classes of occurrence – those species which are fairly common and regular members of the community and therefore consistently sampled and those species which move in and out of the region and can be considered as transient. This latter group can appear rare but in fact may be widespread and although locally in small numbers are relatively abundant at a regional scale. The data presented above would appear to endorse this perspective. Figure 3a. Numbers of macrofaunal polychaetes recorded only in one sample (rare species) from studies with different intensities of sampling effort (number of samples). There appears to be no direct relationship between the number of samples analysed and the tail of rare species recorded, suggesting that rare species will always be a feature of deep-sea samples. The data were derived from a number of abyssal studies. Figure 3c. Percentage occurence of rare species does not appear to decrease substantially as samples are pooled. At least 50% of the fauna is represented by rare species.The Domes A samples removed from the analyses to avoid the potential influence of the poor sorting. Figure 3b. The polychaete samples analysed to test for the effect of sampling on the numbers of rare species. As the samples are pooled as expected the numbers of rare species decreases but it does not appear that the decrease drops significantly with larger numbers of samples at the regional scale. Fig 3d. Percentage occurrence of rare species in nematodes from the EqPAC transect shows a marked reduction as more samples are pooled. But the number of rare species is still a high percentage when all the samples are pooled. 15 Table 4. Results of rarity analyses No. of species CCFZ Rare 87 47.8 Locally widespread, rare and restricted 13 7.1 Locally rare but widespread 64 35.2 Locally abundant widespread but rare in other localities 7 3.8 Locally abundant and widespread 7 3.8 Common 4 2.2 182 100 Total number of species No. of polychaete species % of species No. of nematode species % of species 101 59.3 70 34.3 9 5.3 28 13.7 55 32.4 95 46.6 Locally abundant and widespread 3 1.8 7 3.5 Common 2 1.2 4 2.0 170 100 204 100 EqPac sites Rare Locally widespread, rare and restricted Locally rare but widespread Total number of species 16 % of species 3. Study areas: current biogeographic knowledge and gaps MIDAS has focussed on four main environments as part of the assessment of mining impacts: nodule province of the CCFZ, hydrothermal vents, non-active vents and non-vent hard substratum environments, and methane hydrate zones in the Arctic and in the Black Sea. In this section an assessment of the current biogeographic knowledge of each these environments will be presented. A case study is given to highlight some of the issues that we currently face in assessing in biogeographic studies. The main gaps associated with each area are also discussed. 3.1 CCFZ – sediment nodule fields Figure 4. Map of the CCFZ showing contract areas and Areas of Particular Environmental Interest (APEI).(image ISA) This region has been of interest since the 1970s and subject to a number of studies (Mincks and Smith, 2006). Currently, 13 different contractors have exploration licences and are undertaking a range of environmental studies in their respective claim areas (Fig. 4). An extensive bibliography has been accumulated for this region ranging from survey reports (often grey literature) to comprehensive studies. 3.1.2 Physical and biogeochemical gradients in the CCFZ Deep-sea biogeographic patterns are frequently correlated with large-scale biogeographic gradients [Lutz et al. 2010; MacClain et al. 2012; Smith et al. 2008]. Within the CCFZ there are two major biogeochemical gradients – an east to west decrease in the nutrient flux to the ocean floor (Wedding et al. 2013) and by a south to north decrease in nutrient flux (Smith et al. 1996; 1997). C R Smith in a presentation to the ISA proposed that the CCFZ could be divided into nine regions, each with a different export regime. These gradients have an impact on the local abundance and diversity of elements of the fauna, such that for macrofauna there is a significant decline in their abundance from east to west and from north to south (for example, Glover et al. 2002; Paterson et al. 1997; Smith et al. 2008). Large ocean basin cycles also impact on parts of the area, in particular El Nino Southern Oscillation impinges the southern edge of the CCFZ. Productivity may affect recolonisation and recovery of exploited areas. Studies of large-scale disturbances in the deep sea returned conflicting results. The analyses of the 17 DISCOL area, following a simulated mining experiment, indicated that the densities of motile megafauna had returned to predisturbance values after three years. Similarly, Glover et al (2001) found that polychaete assemblages were still impoverished after decades in turbidite-affected sediments off Madeira. Such a large-scale disturbance can be considered as an analogue of a the kind of disturbance which might result following mining. The extent of the area disturbed coupled with overlying production appeared to have an important effect on recovery, but also depending on the fauna investigated (in terms of size class, life history and functions). Although studying disturbance at a much smaller scale, Miljutin et al. (2011) investigated the recovery of nematode assemblages following experimental test mining the CCFZ. The nematode samples from tracks of the mining vehicle were compared with nematode assemblages from unaffected areas away for the test zone. After 26 years, there were no signs of recovery neither in densities, diversity nor composition (Miljutin et al . 2011). 3.1.3 Scales of habitat heterogeneity Local topography is heterogeneous with differences in the slope and nodule coverage at the km to 10s of metres scale (e.g. Veillette et al. 2007). There are also large topographical features such as abyssal hills through to seamounts (Fig. 5). This means that to understand fully the faunal composition of the region, the links and connectivity both within and between areas within the CCFZ have to be established as part of future sampling programmes. Site Position Average annual surface productivity (gC m–2 yr–1) Certain elements of the fauna also provide habitat for other organisms such as large xenophyphores and komokiaceans. These structural taxa respond to various biogeophysical gradients and cycles and so understanding the biogeography of the structural taxa and what drives the observed patterns will also help in understanding the spatial distribution of associated taxa. EQPAC 0° 0° N, 140° W 230 3.1.4 Molecular approaches EQPAC 2° N 2° N, 140° W 200 EQPAC 5° 5° N, 140° W 155 EQPAC 9° N ?9° N, 140° W? 140 DOMES A 8° 27’ N,150° 47’ W 140 PRA 12° 57’ N,128° 19’ W 130 ECHO 1 14° 40’ N, 126° 25’ W 135 Molecular approaches, here defined as the use of DNA sequence data, has great potential in the analysis of biodiversity and connectivity in areas of resource exploitation. Molecular data have been used extensively in recent years to address questions and challenges in both marine and terrestrial conservation science. These questions range from the simple (e.g the identification of species through DNA barcoding) to the complex (e.g the analysis of the ecosystem function and response of organisms through study of their expressed genomes). The technological progress of laboratory Table 5. Surface productivity estimates from a number of sites across the CCFZ (from Glover et al., 2002 based on Smith et al. 1997). South to north gradient West to east gradient 18 sequencing and computer analysis of DNA sequence data has been extremely rapid, driven to a large extent by biomedical, agricultural and forensic science. An example is high-throughput sequencing (HTS), typically using sequencing machines manufactured by the company Illumina and now widely employed across almost all biological sciences (Davey et al. 2011). These machines have the potential to produce large reads of genomic DNA sequences, at relatively low cost, from large numbers of samples. However, a note of caution must be sounded: blindly collecting large amounts of ‘data’ using HTS without careful consideration of the scientific questions are being addressed may not be the best approach to filling the gaps. There are two main issues, firstly that of taxonomy and secondly that of sampling bias. In both cases, molecular data can in fact be the solution to the problem, but only through careful and integrated workflow pipelines that link new data to old. Taxonomy, at its core a naming system that allows biological data about organisms to be linked together, is almost completely lacking in areas such as the CCFZ. This means for example that contractors or researchers at different sites in the CCFZ produce species lists that are not comparable with each other, and that these species lists are never transferred into formats that others can use (ie named or described species linked to voucher material). For example, despite over 130 scientific cruises to the CCFZ since the 1970s (Nimmo et al. 2013), there is not a single record of a polychaete (the dominant macrofaunal taxon) on the Ocean Biogeographic Information System (OBIS) within the 5 million square km area. In comparison, a 300,000 square km box drawn in the North Sea between United Kingdom and Holland produces 95,000 polychaete records on OBIS. The reason is that the North Sea has a well worked taxonomy and useful field guides to the local fauna. Molecular taxonomy can help, and in fact has the potential to rapidly speed up this process. Firstly the description of new taxa can be undertaken using DNA data as the primary descriptive tool, alongside high-resolution imagery of key morphological features (assisting identification at a later stage). This sounds simple but requires a comprehensive and fully-funded sampling-analysis-data-archiving pipeline that starts with careful sampling and imagery, sequencing of useful DNA markers and proper archiving of voucher and type material in acessible museum or research collections linked to online databases. The ‘speeding up’ of the taxonomic process (Butcher et al. 2012) is enabled as the DNA data takes away uncertainties with regard species identity, and is easily archived on well-organised global databases (NCBI GenBank). Molecular taxonomy does not require HTS, but it does require a degree of morphological data and proper archiving of voucher and type material. Molecular taxonomy is not the same as DNA barcoding, which in its typical definition is the identification of organisms by sequencing a short DNA marker, such as cytochrome oxidase I (COI) (Vogler & Monaghan 2007). Molecular taxonomy is the creation of the reference database that allows DNA barcoding to take place. In the example of the CCZ, it is not currently possible to ‘DNA barcode’ organisms from samples there because at present there is no reference database to compare the sequences with. It is possible, but unlikely, that matches will be made from better studied regions (e.g bathyal depths or shallow water) but recent surveys are showing that it is very rare (Helena Wiklund, pers. comm.). A major effort must be made to undertake quality molecular taxonomy from poorly-known regions, the resulting database that will be generated will allow future research and monitoring programs to undertake rapid biodiversity assessment using DNA barcoding. DNA data has the potential for far more than the creation of the basic, but essential, taxonomic database. The genomes of organism contain a wealth of information on the demographic history of the population to which it belongs as well as the functional adaptation of the organism to its environment. One of the more applicable with regard deep-sea mining impacts is the estimation of migration rates (e.g across the CCFZ or between hydrothermal vents) and effective population size, which can be undertaken from a surprisingly small sample size of just a handful of individuals (Beerli & Felsenstein 1999). However, there is debate as to whether this approach would be able to detect intraspecific variation so more work needs to be done to clarify this. Today HTS approaches, with the enormous amounts of genetic data from just single individuals, and the sequencing across whole genomes in non-model organisms (NMOs) allow this to be undertaken using analysis of just one or two individuals. functional response of that organism to environmental stress or variable. Of these approaches, all of them potentially of great benefit, those most likely to be applied to questions of biodiversity and connectivity in deep-sea mining regions in the near future are probably (2) RAD-seq and (4) eDNA. The RAD-seq approach can be applied to the study of population genetics, with questions such as migration rates, gene flow, and effective population size being of direct relevance to management decisions. There are examples of RAD-seq being used in population studies in aquatic organisms (e.g Keller et al. 2012), but rather few to date in the deep-sea, (although see Pante et al. 2014). A recent study of HTS approaches on NMOs can currently be split into five major categories (Willette et al. 2014): (1) Whole Genome Sequencing (WGS) i.e the sequencing of entire genomes and assembly using reference transcriptomes, (2) RAD-seq, i.e the Figure 5. Seafloor image showing seafloor topology in the eastern German Claim area sequencing of a large and of the CCFZ. The brown features representing seamounts of varying elevations and randomized sub-sample of shorter sizes. There are a large number of such features within the area. Knowledge of the fauna associated with some of these features would be useful to place the species in homologous regions of the genome the mining areas into a biogeographic context. (Image from BGR) using restriction enzymes, (3) the Amplicon method, i.e the enrichment (using PCR) of a large number of genetic regions of interest (or markers) across whole genomes typically thousands of single-copy genes/exons for the study of phylogenetics, (4) the metagenomic approach, or more commonly ‘environmental DNA or eDNA’ method in which a large number of sequence reads from environmental samples (e.g mud or seawater) are assembled into operational taxonomic units (OTUs) which are assumed to be species and can be analysed as such across samples or regions of interest and (5) RNA-seq (or gene expression study) which is a fairly rapid way of investigation which genes are being expressed in an organism or group of organisms and as such the 19 foraminifera in the Southern Ocean deep-sea has revealed the potential utility for rapid assessment of potential biodiversity in eDNA samples (Lejzerowicz et al 2014). Sampling bias, an age-old problem in deep-sea studies, does not disappear with the advent of HTS. RAD-seq approaches, even with decreasing costs in the future, are unlikely to applied to anything other than abundant and well-characterised species such as charismatic vent fauna. eDNA has great potential to provide long-lists of OTUs for microbial and protists species, but there is currently no knowledge of the spatial scale of sampling required to take eDNA samples of larger metazoa. Careful, academic research must be undertaken on these biases and the appropriate sampling scales required before these HTS tools become widely used by contractors involved in EIA assessment. 3.1.5 Patterns of biodiversity in the CCFZ Current Knowledge: Biogeography of major faunal groups Megafauna Tilot (2006) has provided an overview of the current knowledge of megafaunal abundance, diversity and biogeography. However most of the information in her report is restricted to a few sites within the CCFZ, especially the French claim areas. Data are lacking on the distribution of megafauna from other claim areas. A workshop on megafauna was held in 2013 at Wilhelmshaven, Germany which brought taxonomic experts and contractors together. A review of the morphotypes of various key groups, holothurians, sponges, fish, etc. was undertaken. An atlas of these morphotypes is available (http://ccfzatlas.com). In addition a series of recommendations was made to the ISA. The main one being that specimens were desperately needed to provide good taxonomic resolution, images and video were not enough. Current knowledge is restricted to a number of scientific publications, which are often old (>30 years) and in need of updating and revision. Nevertheless, the taxonomy of certain megafaunal taxa is quite well 20 established, i.e. echinoderms, fish, some cnidarians, while others need much more work, i.e. sponges and protistans. Meiofaunal biodiversity Yet, to date, studies in both nematodes and harparcticoids have been conducted at various taxonomic levels (species, generic and family), which greatly hampers intercalibration of data sets (e.g,, Miljutin et al. 2011, Ahnert & Schriever, 2001, Vopel & Thiel 2001). Furthermore many families of, for example, harparcticoid copepods had been revised in recent years making comparison with historical data sets, such as DISCOL (Ahnert & Schriever, 2001) even more challenging (P. Martinez, pers. communication). To date there have not been any molecular study examining the diversity and connectivity of meiofauna across the CCFZ. There are a number of studies dealing with the meiofauna from the Pacific polymetallic nodule area, e.g. Hessler and Jumars 1974; Snider et al. 1984; Mullineaux 1987; Renaud-Mornant and Gourbault 1990; Bussau et al. 1995; Radziejewska and Modlitba 1999; Ahnert and Schriever 2001; Radziewska et al. 2001a,b; Radziejewska 2002; Lambshead et al. 2003; Miljutina et al. 2010 and Miljutin et al. 2011. Radziejewska (2014) presented the most recent Microbial biodiversity update on meiobenthos from the Central Pacific nodule province. So far investigations of meiofaunal biodiversity in Protista. Protistan communities from the CCFZ are mostly the CCFZ have mainly focused on harpacticoid copepods known from Foraminifera. This group is abundant in and nematodes, which typically represent the most samples and is thought to play a substantial role in carbon dominant meiofaunal taxa in the abyss. Though other cycling. Larger protistans, like xenophyophores may also higher taxa, such as ostracods, kinorhynchs, tardigrades, provide habitat for other eukaryotic organisms (Lecroq et al gastrotrichs, and acari are also present (Radziejewska 2009). Abyssal Foraminifera morphotypes have been 2014). Analyses of nematode assemblages revealed studied extensively, for example Gooday et al. (2004); between 10 and 246 nematode genera per study with most Schröder et al. (1988). occurring throughout the CCFZ. However, differences in generic composition between the eastern CCFZ and the Some of the hard-shelled morphospecies recorded in central part of the CCZ have been reported (Radziejewska et Kaplan samples from the Eastern and Central CCFZ by al. 2001, Miljutina et al. 2010); the nematode genera Gooday et al. in Smith et al. (2008) were widely distributed Terschellingia, Desmoscolex and Pareudesmoscolex seem to in the Pacific and other oceans, such as Adercotryma be more prevalent in the eastern CCFZ whereas glomeratum, Spiroplectammina biformis, Lagenammina Thalassmonhystera, Theristus and Acantholaimus are more tubulata, L. difflugiformisand and some of the Reophax dominant in the central part. morphotypes (e.g. R. helenae, ‘R. bilocularis’ and ‘R. scorpiurus’). The widely distributed species, however, Between 34 and 62 genera of harpacticoid copepods are represented a small proportion (approximately 7%) of the known from the CCFZ (Radziejewska, 2014). Harpacticoids species recognised at the Kaplan sites. (species, generic and family level) have been used to monitor faunal responses following experimental and test Many of Gooday et al. (2004)’s monothaumous mining in the CCFZ and DISCOL area (Ahnert & Schriever, morphotypes were also recorded widely in more than one 2001, Radziejewska et al. 2001a, Mahatma 2009; Miljutin et ocean basic, i.e. Atlantic and Pacific, and more than one al. 2011). For example, species diversity and the total locality within the Pacific. Nozawa et al. (2006) studied density of harpacticoids recovered after 26 years following sediment Foraminifera from one site in the eastern CCFZ an experimental test mining study in the CCZ, whereas and reported that some of the morphotypes of species composition had not returned to pre-mining Foraminifera and komokiacean were similar to those with a conditions (Mahatma 2009). wide distribution, recorded from other ocean basins. Nevertheless, several other morphotypes had a more Macrofauna – Case Study 2 It is important when analysing biogeographic patterns to understand whether the fauna changes because of turnover – the replacement of one species group with another – or whether the fauna appears to be a restricted subset of the overall species pool – nestedness. It is important that these two aspects of b-diversity are separated or at least their contributions to the measurement is detailed, as very different conclusions and therefore management decisions can be made. For example, nestedness implies that the fauna is fundamentally one species pool and that activity, which might cause an effect in one area, is unlikely to impact the pool generally. However if faunal turnover is the primary finding activity in one area could have profound impacts on the fauna. In the CCFZ it has already been established that there are both east – west and north – south gradients (Table 5) and so it is hypothesised that such gradients will structure the biogeography of the benthic fauna. In other deep-sea studies the impact of productivity gradients has led to both nestedness in the case of molluscs – particularly gastropods (Brault et al. 2012) but also turnover – asteroids (Brault et al. 2013). So it appears that different taxa will respond in different ways to potential gradients. In this section the response of macrofaunal polychaetes will be tested using archived data from a number of studies in the CCFZ. The reduction in nutrient transport across the CCFZ may well result in a reduction in the similarity of the fauna. It is reasonable to hypothesise that such a gradient may well lead to nestedness. “Put Spionid polychaete from the CCFZ with an apparent widespread distribution (NHM) simply, as POC-flux decreases … species drop out because of insufficient food supply.” (Brault et al. 2013). The first based on a comparison across the CCFZ undertaken by George Wilson (Australian Museum; Paterson et al. 1997). The second was an analysis of polychaetes from the EqPac programme (Glover et al. 2002). A third dataset made during the Kaplan study (Smith et al. 2008) also studied sites from the east to west of the CCFZ. Unfortunately none can be amalgamated because of differences in the taxonomy; each was identified by a different set of taxonomists. In both samples sets there were a high number of new species and so classification was to a numbered OTU. These OTUs have not yet been intercalibrated. A fourth data set looked at meiofaunal nematodes identified from the EqPac transect (Brown, 1998; Brown et al. 2000). While this taxa is not strickly macrofauna they are included to indicate the generality of the patterns obtained. Methodology The analyses of b-diversity follows the approach outlined by Baselga (2010). The methodology proposed provides a more robust separation of the contributions of nestedness and turnover to the dissimilarity between sites than previous methods, i.e. NODF (Almeida-Neto et al., 2008). Analyses was undertaken using the R scripts provided by Baselga (2010) – beta-multi.R and beta-pairwise.R (R Development Core Team, 2006). The analyses are based on presence-absence of species. rather that different species area able to survive at different points along the gradient. Results However these results are based on low numbers of sites and /or sampling within a site. Only DOMES A had sufficient sampling for the species accumulation curve to approach asymptote. This means that effectively each site could be a random subset of the regional species pool, the lack of nestedness notwithstanding. This is because of the large ‘tail’ of rare, apparently site-unique, species encountered at all sites sampled. Sufficient sampling will never be achievable so detecting the real distribution of these species will remain problematical. Nevertheless the implication for assessing environmental impacts in the CCFZ is that consistent sampling across the area, together with unified taxonomy is essential. While it may not be possible to sample adequately to solve the problem of the ‘rare endemics’, a stratified and comprehensive sampling programme will go someway to providing the data that can be used to support management. CCFZ east to west indicates that overall dissimilarity Beta SOR is =51% but that there is virtually no nestedness (B NES = 0.02) while the turnover component is relatively high (B SIM = 0.49). There are several dominant species shared across the CCFZ which underlie the overall similarity and possible contribute to the nestedness value. The EqPac north–south gradient shows a slightly modified pattern with higher overall dissimilarity (B Sor=0.78) but again indicates that it is turnover (B Sim = 0.63) not nestedness (B NES = 0.16) which accounts for the pattern in polychaetes. For the nematodes, a similar high level of dissimilarity is observed and also turn-over which appears to be responsible for the patterns observed (BNES = 0.1.7). Overall the results point to degrees of faunal turnover with distance as postulated by Paterson et al. (1997) and Smith et al. (2008).. The biogeographic analyses points to a relatively large number of species common across the CCFZ, both east to west and north to south but with a varying number of unique species at each site. It does not appear that the nutrient gradients found in the CCFZ drive nestedness of the fauna as might have been predicted, Table 6. b-diversity measures for the CCFZ stations based on polychaetes – PRA, ECHO and DOMES A; Polychaetes and Nematodes–EqPac 0N, 2N, 5N, 9N and HOT station 23N, Kaplan polychaetes. SIM=Simpson’s multiple site dissimilarity; SOR = Sorensen’s pairwise dissimilarity measure of turnover, NES = nestedness measure. $beta.SOR dissimilarity (spatial turnover) $beta.SIM â-diversity $beta.NES (nestedness) Polychaetes multi.ccz 0.511 0.487 0.0244 Polychaetes Kaplan 0.806 0.625 0.181 Polychaetes multi.eqpa 0.783 0.626 0.156 Nematodes EqPac 0.629 0.460 0.168 Image of the head of the nematode Acantholaimus angustus from the CCFZ (Senkenberg) 21 Table 7. Summarising the bacterial and archaea composition found in COMRA area A (Xu et al. 2005). The majority of the clones isolated had a high (95%) similarity to other environmental clones from other regions Bacterial composition 46 clones (RLFP) 32 species 27 genera (16SrDNA) Proteobacteria i. Gammaproteobacteria ii. Alphaproteobacteria iii. Theta/deltaproteobacteria 75% 92.9% 5.3% 1.8% Non-sulphur bacteria 5.3% Cytophaga-Flexbacter-Bacteroides 11.8% Unclassified 5.3% Archaea composition Marine group 1 - Crenarcheota 22 clones restricted distribution within the Pacific e.g. Resigella moniliforme, or within the CCFZ region. Nozawa et al. (2006) stress the need to re-examine the original material. There were at least 10 species of foraminifera that are abundant at one of the three Kaplan sites, but rare or absent at the other. Molecular analyses of Foraminifera has revealed considerable diversity, much of it not yet related to morphotypes and thought to reside in the small size fraction of benthos (Lecroq et al. 2009ab;2011; Pawlowski et al. 2011). New discoveries are likely to increase with greater sampling. These analyses indicate that most of the deep-sea sediments are undersampled with regards to this taxon. Yet despite this high biodiversity, molecular evidence suggests that for at least some species, biogeographic distributions may be quite large and interoceanic (Lecroq et al. 2009). 22 In general though, Foraminifera show similar biogeographic patterns to other elements of the benthic assemblages, some apparent cosmopolitan species, those with a more ocean basin distribution and then those with restricted distributions. The data suggests that significant turnover of major components of the foraminiferan faunal over scales of roughly 1,000 km across the CCZ. More sampling is needed to provide a better understanding of the distribution of this group at a number of different spatial scales. form biofilms and so may be particularly associated with nodules and their formation (see section below). Virtually nothing is known about other protistan taxa such as ciliates, flagellates etc. The importance of this obvious gap in knowledge is also unknown. Xu et al. (2005) reported that the majority of the bacteria isolated were Proteobacteria (Table 7) and that the majority appeared to be similar (>95%) to isolates from other environments. Many proteobacteria are metal-reducers. It is perhaps not surprising that such types of bacteria are commonly distributed and recorded from habitats such as vents. The Cytophaa- Flexbacter- Bacteroides group showed a higher degree of novelty with only 61-93% showing a degree of similarity with other environmental clones, but again many isolates appeared related to those from a very wide range of habitats. The implications from Xu et al. Bacteria. Bacterial communities are thought to be important foundations for the nutrient cycling and related ecosystem services. In nodule areas microbial communities, particularly bacteria, also play a role in metal cycling and may be involved with nodule formation (Xu et al. 2005; Wang et al. 2010). Some of the isolates belonging to the Pseudomonas are known to Studies of such communities are not yet common in the CCFZ (Wang et al. 2010, Wu et al. 2013; Xu et al. 2005; 2007 see Tables 7 and 8) but some patterns can be deduced. Bacterial communities are often diverse and show similar taxon richness and community patterns as other elements of the fauna such as macrofauna, e.g. high diversity with long tails of unique or rare taxa. Table 8. Results from Wang et al. (2010). Four sites across the central Pacific, the first three within the CCFZ, the fourth outside the Area and close to the continental slope of Mexico. Station Depth No of sequences isolated No of OTUs Dominant group West WS0505 Highest nodule concentration 5120 m 68 47 Alphaproteobacteria Deltaproteobacteria East ES0502 5307 m 63 58 Gammaproteobacteria Alphaproteobacteria East ES2005-01 Lowest nodule concentration 5197 m 67 45 Gammaproteobacteria Alphaproteobacteria ES2005-03 Lowest concentrations of MnO and Fe2O3 4299 m 115 71 Alphaproteobacteria Deltaproteobacteria (2005) are that the microflora from the nodule province is composed of representatives closely related to widespread clones. Molecular sampling methodologies have improved and so has the resolution. However, molecular sampling methodologies have improved and so has the resolution. The low numbers both of samples and of isolates cautions against reading too much into these early results. In a comparative study of four sites across the CCFZ, Wang et al. (2010) found a similar bacterial composition to Xu et al. (2005). The Proteobacteria were the dominant group within which Alphaproteobacteria (23.3%) and the Gammaproteobacteria (21.7%) were the most abundant of the isolated sequences. The dominant non-Proteobacteria group was Acidobacteria – 7% of the total sequences. Betaproteobacteria, Chloroflexi, Firmicutes, Verrucomicrobia, were minor with <3% of the sequences. Wang et al. (2010) showed that in terms of biogeography the bacterioflora had similarities with other faunal groups. While many isolates are restricted, approximately half showed basin-scale distributions, being closely related to isolates from areas of ocean crust, the Pacific nodule province and marine basalts; some CCFZ isolates were very similar to clones from these habitats (=99%). The marine basalts and crusts contain large amounts of Fe and Mn and are highly reactive. Wang et al. (2010) suggest that the similarity is related to bacterial function particularly manganese oxidising abilities. Within the CCFZ, there were also differences in the occurrence of particular microbial groups pointing to potential basin-scale biogeographic patterns possibly related to the presence of nodules and their densities. At the level of the isolate, similar patterns to other fauna were observed with some being restricted to one site while others occurred across the CCFZ, or across more than one site. In this latter finding bacterial biogeographic patterns bares some similarity to other elements of the fauna. A polychaete species belonging to the genus Parexogone. This species reproduces by developing stolons along the body which then grow into miniture versions of the adult and break off to lead separate lives. The stolons can be seen in the picture as outgrowths coming off the body. Image: A.Glover (NHM) and Viacheslav Melnik (Yuhzmorgeologia) 23 3.1.5 CCFZ: Nodule fauna Mining will remove nodules. Yet the nodules are themselves a habitat or series of microhabitats which are utilized by a range of organisms (Mullineaux 1987; Theil et al 1993; Veillette et al 2007). Our knowledge of the nodule fauna is remarkably scarce considering the amount of study that has been directed towards them. Previous studies have been mostly geological and mineralogical with little focus on the organisms using the nodules as a substratum or potentially as a substrate. Nodules are one of the few hard surface habitats in the abyss and therefore potentially have a different species composition to the surrounding sediment. The importance of these organisms, particularly the bacteria, is that they may be agents in the formation nodules and accumulation and cycling of metals within them (Lysyuk, 2011; Reiman, 1983; Wang et al. 2009 and references therein). Megafauna. Many taxa appear to use nodules as a potential hard surface upon which to settle and grow. Images taken from the region indicate large protistans, sponges, cnidarians such as actiniarians and octocorals, crinoids, particularly stalked taxa, and ascidians are observed on nodules. Megafaunal organisms often have a large geographic range and photographs do indicate some widespread morphotypes (http://ccfzatals.com). Images cannot totally resolve the taxonomy of the megafauna. More studies based on collected specimens are needed and so it remains difficult to be sure of the distribution and biogeography of large epifaunal taxa. The lack of specimens means that there is no information as to whether the morphotypes observed are opportunistic or obligate nodule epifauna. Similarly if species are not obligate then might they occur on other hard surfaces such as rocky outcrops on spreading centres or on seamounts? Macrofauna. Mullineaux (1987) compared the macrofauna from nodules in two sites in the Pacific, one within the CCFZ (Domes C), and with samples from metallic crusts on a seamount (Table 9). There was a high degree of ‘endemism’ in that most of the taxa recovered were restricted to one nodule site (71%), with only one species 24 Table 9. Macrofauna recovered from nodules and crusts (from Mullineaux, 1987). Taxon No of species Distribution Functional Group Sponge 5 4 species nodule only 1 species nodules and crusts Suspension feeder Cnidaria: Hydrozoa 4 3 species nodules only 1 species crusts only Suspension feeder Cnidaria: Scyphozoa 1 Nodules and crusts Cnidaria: Anthozoa 2 Nodules only Suspension feeder Annelida: Polychaeta 3 morphotyp es 1 species nodules only 1 species crusts and nodules 1 species crusts only Deposit and suspension feeders Mollusca: Gastropoda 3 Nodules only Deposit feders Mollusca: Bivalvia 2 Nodules only Suspension/deposit feeders Mollusca: Mono and Polyplacophoran 2 Nodules only Deposit feeders Crustacea: harpacticoid 1 Nodules only Deposit feeder Crustacea: Tanaidacea 1 Nodules only Deposit feeder Bryozoa 8 7 species nodules only 1 species crusts only Suspension feeders Brachiopoda 1 Nodule only Suspension feeder Echinodermata: Crinoidea 1 Nodule only Suspension feeder Tunicata: Ascidiacea 2 Nodule only Suspension feeder being found in all sites and a further one being recorded from the crusts and one of the nodule sites. Four species were common to the two nodule sites (11%). It must be stressed that this was a restricted study and that a greater sampling both of nodules and metallic crusts is need. Mullineaux concluded that a major driver for the species richness was nutrient availability particularly nutrient flux from the surface. Thiel et al. (1993) studied the epifauna associated with nodules and detected that there were distinct microhabitats such as crevices between and within nodules which were inhabited by a distinct fauna. Little is known about the spatial distribution of such epifauna assemblages at a basin scale or between ocean basins or whether such fauna also occur in metallic crusts on seamounts. Protistan assemblages. Mullineaux (1987) found 64 species of Foraminifera on nodules, the dominant group. Table 10 shows the numbers of species recorded from sites and between sites. While many species appeared to occur across the sediment sites of the Eastern North Pacific and Central North Pacific, the fauna found on the crusts were not found in the nodule sites. Mullineaux noted that there was little overlap between the nodule fauna and species found in the sediment. She found that the abundance and percentage cover of many Foraminifera suggested that they may also be important agents in nodule formation. Veillette et al. (2007) studied four nodule zones, three within the French eastern and one in the western CCFZ site (Table 11). Each zone had a different nodule facies with different sizes and coverage of nodules. The aim of the study was to assess whether there were distinctive microhabitats within the nodules and whether there was a distinct associated fauna with them. The vast majority of foraminiferal morphotypes (85%) were widespread across the two sites and also across the different facies identified. This contrasted with the macrofauna species which appeared more endemic recorded from only one facies of the eastern region (59%). In terms of local diversity, the eastern sites were more species rich than the west, possibly a measure of the higher nutrient flux in the east. They also noted that exposed nodule surface was a significant factor in determining the number of species found on nodules at a regional scale. They concluded that the greater the range of microhabitats, i.e. nodule surface complexity, the greater the habitat heterogeneity and therefore more species diversity. Veillette et al. (2007) noted an overlap with Mullineaux (1987) species, 21 of the 73 species they recorded similar to those of Mullineaux. Caution needs to be exercised in interpreting these results. The study relies on morphotypes. Molecular analyses on Foraminifera have returned equivocal results with some species appearing to be widespread but many apparently more restricted, possibly endemic. Further work is needed focused on reconciling morphotypes with a molecular barcode (Pawlowski et al. 2013) but also which extends the sampling to provide a more rigorous survey across the CCFZ. As with the epifaunal macrofauna, these studies also need to look at the relationship of the fauna associated with polmetallic cobalt-rich crusts and nodules. Table 10. Foraminifera species recorded by Mullineaux (1987) from a number of sites in the Pacific. Crusts is a site of metallic crusts on the Hawaiian Islands chain, the other areas are nodule zones, ENP is within the CCFZ, CNP is outside the Area. No of species Eastern N Pacific Cen tal N Paci fic Cru sts Crusts + ENP/C NP ENP & CNP ALL 19 2 12 5 16 8 Table 11.Spatial distribution of fauna found on nodules based on Veillette et al. (2007). Two sites were sampled – one in the East with three different nodule facies and one in the West with just one facies. Both East and West All E facies Two E facies One East facie W only Foraminifera morphotypes 61 1 3 5 2 Macrofauna 3 4 10 Microbial Assemblages. Wu et al. (2013) compared the bacterial assemblages of metallic nodules from two sediment sites in the CCFZ and from the flanks of a seamount (Table 12). In addition they compared the diversity and composition of bacteria on nodules and in the 25 sediment. The diversity of bacteria as determined from 16S rRNA sequences was high with 1013 bacterial sequences representing 414 OTUs and with 640 sequences representing 31 Archaea MOTUs. While there was a degree Table 12. Degree of specificity between nodules and the sediment for the bacteria based on Wu et al. (2013) Nodules Only Both Sediment only Bacteria 189 55 170 Archaea 10 12 9 of overlap between the nodule assemblages and the sediment, each habitat had a distinctive assemblage (Table 12). Wu et al. (2013) reported that the bacteria appeared to be more diverse in the sediments at two sites but at the Table 13. Bacteria and Archaea community composition from sediments and nodules 15 phylogenetic bacterial groups unique to nodules Gammaproteobacteria 57 OTUs Alphaproteobacteria 41 OTUs Deltaproteobacteria 37 OTUs C Closely related to vent OTUs implying metal cycling functionality 155 phylogenetic bacterial groups unique to sediment Deltaproteobacteria 26 44 OTUs Many related to bacteria from various marine sediments third there was no difference, indicating a degree of heterogeneity. But they also reported that the rarer bacterial groups were found on the nodules perhaps a suggestion that the nodules have specific assemblages. The Archaea had a higher diversity on the nodules that from the sediment and overall had a lower diversity compared to the bacteria. on seamounts, and nodules. More work across all the size categories is needed to establish the degree of commonality as these areas beyond the easily exploitable seafloor may harbour the same species and thus act as a reservoir either for subsequent recolonisation or for future conservation. 3.1.6 CCFZ: Gaps to our knowledge In terms of assemblages the composition differed between sediment and nodules (see Table 13). The members of each Sampling and sampling methodology assemblage appeared to have close similarities to bacteria There is a lack of standardisation in both the sampling from other marine environments; the nodule bacterial OTUs programmes and the methodology. Despite decades of more closely to those from vents and other metal rich exploration few data have been made available in the environments while the sediment OTUs to other marine scientific literature. These datasets are a mix of academic sediments. research cruises, i.e. JGOFS, and environmental cruises by contractors, i.e. KORDI, IOM (see Mincks and Smith, 2006). Comparison of the fauna between the three study sites The latter data may be available to the ISA but are not showed a similar pattern for both the bacteria and archaea, generally available to the wider scientific community. the nodule assemblages from one of the CCFZ sites (WS0904) and from the flank of the seamount clustered together and all the sediment sites clustered as a group. This suggests that the sediment assemblage was comprised of widely distributed OTUs while the nodule assemblages had more distinct and restricted assemblages (Wu et al. 2013). They concluded that microhabitats were more important than distance between localities and that similarities between the bacterial assemblages had more to do with functional aspects, particularly metal cycling, than with biogeography. As with the other faunal studies caution must be taken in interpreting these results because of the lack of sampling across a range of scales. However, the association of some isolates with nodules and their hypothesised role in nodule and mineral cycling needs further investigation. In particular are these isolates only found on nodules or are they do they have a more diverse metabolic repertoire? The answer could have important implications for exploitation and the role of APEIs. Tantalizing preliminary evidence suggests that for some of the taxa there may be links between metallic crusts, found Such a gap in data availability makes any assessment of the distribution and biogeography of fauna difficult. However even the release of data will only be of limited value because there is little intercalibration of the taxonomy between the contractors and even between the academic studies. This is because many of the species being discovered are new to science and the formal taxonomic classification of these new taxa lags far behind the environmental studies. Large-scale geographic coverage The central and north-eastern areas of CCFZ are without data. Claim areas are undersampled. There are no data available for the APEIs. Nor is there any incentive or regulatory imperative for any of the contractors to undertake surveys in such areas. So there is no way of knowing how representative these areas might be. Sampling standardisation and effectiveness for biogeographic and connectivity studies Different sampling gears collect different elements of the fauna. Quantitative devices such as box cores generally provide good samples in nodule areas but they suffer from the drawback that the macrofaunal infauna is sparse so considerable sampling is required to adequately sample the environment. This results in considerable effort to process the resultant cores to isolate specimens. Quantitative meiofaunal samplers such as multicorers are faster and provide larger numbers of individuals but identification of the meiofauna can be problematic – even of a molecular approach is employed. Next Generation Sequencing and omic (genomic, transcriptomic and proteomic) approaches may provide a solution but the costs in terms of processing samples and the informatics involved mean that this approach is, at the moment, still beyond standard monitoring methodologies. Qualitative samplers such as epibenthic sledges are able to provide enough specimens of macrofauna and smaller megafauna. The fact that they are not quantitative is not a drawback for biogeographic studies. Again combining this gear with molecular and morphological study can provide data useful to connectivity (i.e. population genetic studies) and biogeography. Some of the contractors are now using this approach. The disadvantage to this is that epibenthic sledges, like other sampling devices, may be selective in the organisms and size classes they capture. Different contractors undertake megafaunal surveys in different ways, over different amounts of surface area, using a variety of gears from video sledges to ROVs and submersibles. The results of a workshop in 2013 on megafauna from the CCFZ indicated that imaging was insufficient for the kind of taxonomic resolution required to assess biogeographic distribution of key megafaunal species. Existing Data and Access Currently we lack data on faunal distributions across a range of scales from local to regional. Given the heterogeneity present across the region again at a range of scales this makes it particularly difficult to assess connectivity and patterns of biogeography of the fauna. Similarly, different faunal elements, from micro to megafauna, are likely to demonstrate different patterns of distribution across the abyssal regions. Contractors follow the regulations and guidelines as laid down by the ISA and are collecting data, very little of it is available to either other contractors or to the scientific community. Taxonomic impediment Much has been made of the taxonomic impediment in deep-sea biodiversity research. There are a number of reasons for the lack of formal taxonomic study of the samples already available but the consequence is a lack of data just at a time when the need is crucial. The ISA has recognised this and is convening a series of workshops to try to standardise taxonomy between contractors. The main issue in developing a biogeographical overview of data being collected by the contractors for the ISA is that there is no taxonomic consistency between the contractors. The nature of the contracts for exploration does not encourage or require any collaboration between contractors. All contractors are required to undertake is environmental assessments of their own areas and submit these data to the ISA. Therefore, when assessment of potential impact are being addressed it is actually impossible for any contractor to be able to say with any degree of certainty that exploitation of the mineral resources will not have irreversible consequences, such as the extinction of species, because there are not the data to be able to compare species between areas. The ISA is the obvious organization to enable this comparison to be made by making data available from the contractors. But consistent taxonomy of all groups is lacking to enable these fundamental analyses to be undertaken. This obvious failing has been recognized the Legal and Technical Commission of the ISA. A number of taxonomic standardization workshops is being organized to address the lack of data. Taxonomic standardisation is compounded by a lack of expertise–some contractors do not have access to trained staff and there is generally a lack of taxonomic experts to aid with training or undertake the identification and standardisation of the fauna. This lack is often not just the lack of a suitable taxonomist but the lack of time available for existing experts to be able to work on samples from the CCFZ. Some areas such as the Protista also have shortages in expertise. The microbial area is also neglected and in this case it is a result of the regulations not requiring microbial assessments of the benthic environment to be undertaken. For the megafauna few samples of individual specimens are available and as a result there is little genetic information. Fauna on nodules Is it distinct? Significantly different from surrounding sediment? If so how? Are their specific obligate relationships or is it facultative and opportunistic? For example, sponges are a dominant element of the megafauna. How many images of sponges show them associated with nodules? We do not yet know if the same sponge species are to be found in areason seamounts as well as the nodule areas. In nematodes there seems to be a difference between the species found in nodules crevices and those found in the sediments surrounding nodules (Thiel et al. 1993). There will need to be further research to assess whether these differences rise from different abundance levels of faunal assemblages, or if nodules and sediment clearly host different sets of species and genera. Microbial communities form biofilms on nodules and are thought to be involved in nodule formation and manganese oxidation but it is not clear whether this relationship is obligate and preliminary evidence suggests that many bacteria and archaea may be shared across metal-rich habitats. Nodule fauna relationship to fauna on hard substratum habitats on seamounts within claim areas and across the region generally needs to be investigated. 27 Molecular sampling Megafauna Taxonomy Advances in molecular techniques may provide a faster and more pragmatic approach. Currently, a major disadvantage of most methods is that they do not provide estimates of abundance of the organisms being isolated. And even here new approaches in transcriptomics will enable future studies to undertake a full molecular assessment. The pace of technological advance in this field of analyses and interpretation suggests that some form of molecular methodology will become routine in general ecological research. However, for the current contracts such methods have not been widely used and are unlikely to become routine in the short term. KNOWLEDGE GRID for CCFZ Sampling The lack of molecular data, given modern taxonomic practice, is particularly limiting in addressing the issues of species distribution and biogeography. Increasingly survey teams are turning to molecular methods to provide quick taxonomic results (A.Glover, pers. comm.) But there is still a need to provide such data to enable comparisons between contract areas and thus provide the biogeographic patterns needed to assess potential impacts of exploitation. Meiofauna Microbial: Protista Microbial Bacteria Microbial Archaea Taxonomic resolution Good general knowledge of species But wide use of morphotypes reduces resolution Limited to a few taxa. Mostly (M)OTU Limited to a few taxa Mostly (M)OTU. Some taxa only to family or genus level Foraminifera and Xenophypohores. Many (M)OTU Limited MOTU Limited MOTU Keys, etc Available for many groups but may be in need of revision Available for some groups None available None N/a N/a Taxonomic information availability Some exchange of information between scientists No intercalibration No intercalibration Mostly academic exchange Data available via external databases such as GenBank Data available via external databases such as GenBank Sampling Mostly video and still imaging Quantitative and qualitative samples Quantitative samples Quantitative Quantitative Quantitative Molecular sampling Little Limited Limited Some All molecular - eDNA All molecular eDNA For video and stills but not for specimens. Collecting gear used is based on ISA recommendatio ns but new gear being introduced – epibenthic sledges ISA have recommendations which apply Limited but follows accepted scientific standards Limited but follows accepted scientific standards Limited but follows accepted scientific standards Some but scattered in scientific literature. Spatial coverage insufficient Some but scattered in scientific literature. Spatial coverage insufficient Assessment for some groups – Harpacticoids Spatial coverage insufficient Some preliminary assessments but coverage limited Some but limited to a few areas Some but limited to a few areas Consistent sampling between different teams Biogeogrphic data 28 Macrofauna 3.2 SMS – MIDAS areas from the Mid Atlantic Ridge The Mid-Atlantic Ridge (MAR) in the North Atlantic is a semi-continuous, complex series of rocky ridges, fracture zones, seamounts and areas of volcanism. The ridge is an active spreading centre with a central rift valley. Features such as sediment fields found in areas between rock ridges and outcrops represent 95% of the MAR. The combination of habitats contributes to environmental heterogeneity on a range of scales (Priede et al, 2013). The MAR separates the east and west Atlantic basins potentially acting as a barrier to distributions but has also been hypothesized as a pathway and stepping stones for the distribution of fauna. The circulation of water masses within the Atlantic is affected by the MAR and deep interchange between the two main basins is complex often being focused on particular features such as fracture zones (Lackschewitz et al. 1996). a b Differences in overlying production have been noted along the ridge, for example, the northern areas overlying the Reykjanes Ridge to the north of the Sub-Polar Front have been shown to have a higher productivity than areas to the south (Gebruk et al. 2010). However, the presence of the ridge itself is not thought to enhance overlying productivity although shallower Figure 6 a) ISA map showing the two claim sites on the MAR. b) MAR around the Azores EEZ, TAG and Snakepit occur within the French claim features associated with the ridge, area while Logachev occurs in the Russian claim area (IMAR). e.g. seamounts, may produce localized areas of increased production and are often associated MAR (Bergstad et al. 2008; Bergstad & Gebruk, 2008) and (Braga Henriques et al, 2012, Tempera et al. 2010) and with deep ocean fisheries (Preide et al. 2013b; Braga UK-led MAR-ECO (Priede et al. 2013a) provided the most sponges aggregations (Tempera et al. 2010). Henriques et al, 2013) recent studies on the MAR outside the Azores EEZ (Fig. 6b). All of these studies report that the high heterogeneity and The MAR is a slow spreading ridge system. The active In general studies on the MAR have been few or particularly undersampling greatly underestimate the biodiversity of volcanism associated with the spreading centres along the localized as in the case of hydrothermal sites with sustained the MAR. The Azores Triple Junction area is also MAR has resulted in a series of hydrothermal vent sites. studies based around the Azores EEZ. Expeditions led by characerised by a high density of non-hydrothermal These are generally widely separated, 100–350km apart Russian oceanographers (Mironov & Gebruk, 2006) habitats often dominated by deep-water coral communities (Murton et al. 1994; German et al. 1996). Van Dover (1995) together with large European multidisciplinary studies ECO 29 suggested that the larger distances between active areas was relatively large and thus might act as a biogeographic filter. At the species level, the some of the larger megafauna Pacific Rise vents (Cuvelier et al. 2014, Gollner et al. 2010b). species have been studied to assess population 12 dirivultid copepods are currently described from MAR connectivity, e.g. the vent mussel Bathymodiolus (Comtet vents and 4 species even occur in other oceans (i.e. Pacific) et al. 2000; O’Mullan et al. 2001; van der Heijden et al. (Gollner et al. 2010a). A first molecular study on dirivultids Despite some biogeographic studies on vent megafauna 2012) and its symbionts (Won et al. 2003; Duperron et al, showed that this taxon might have high dispersal potential using genus data at the global scale, little has been 2006), the blind shrimp Rimicaris (Teixeira et al. 2011; 2012) (Gollner et al. 2011). conducted on different vents from the Mid-Atlantic Ridge. and associated bacteria for some vent fields (Petersen et al. Microbial biodiversity The lists of species lists were published in the 90’s and early 2010), Riftia pachypila at the population level (Shank and 2000’s (Van Dover et al, 1996; Desbruyères et al, 2001; Halinych 2007). Such studies have also compared Protistan. There are few studies of protists around vents. Cuvelier et al. 2011), have just been recently updated, but populations from the two biogeographic provinces Folliculinid foaminifera were found in large numbers on no clear quantitative data, and often with no supporting proposed by Bachraty et al. (2009) – 8ATJ and North settlement panels sited in Clam Acres on the East Pacific genetics analyses. Atlantic. Rise site (Van Dover et al. 1985) which implied that Macrofaunal biodiversity foraminiferans were likely to be found and that some Vent fauna is evolutionarily adapted to the hydrothermal species may be common. No studies have been undertaken chemosynthetic ecosystem. It is highly specialised and often Macrofaunal studies are not numerous. The mussel bed on the MAR. endemic to a particular vent system (Van Dover, 2012). As a fauna of Logatchev, Snake Pit and Lucky Strike were result the biota is particularly at risk should active vent sites Bacterial compared by Van Dover and Doerries (2005). They found be mined. that there was evidence of a north-south split in the faunas, Bacterial biota associated with vents encompass both partly explained by depth, Lucky Strike is shallower than the The vents TAG, Snakepit and Logatchev are currently being free-living and symbiotic species. There is evidence that two southern sites, but also by geological setting at the evaluated by contractors from France and Russia as some symbiotic species also occur as free-living. There are sites. All three sites had many of the same species but with potential areas for the mining of massive sulphide deposits just a few studies on the biodiversity or functional different abundance and could be differentiated as a result. (MSD) (see Fig. 6b). Some of these sites have been studied biodiversity of microbes (Creapeau et al. 2011). since 1988. Meiofaunal biodiversity 3.2.1 Patterns of biogeography Megafaunal biodiversity The large charismatic megafauna are one of the visual characteristics of vent sites. Different vents often having different suites of these organisms. For example, many sites in the Atlantic are characterized by immense swarms of shrimps such as Ramicaris, while those of the Galapagos vents have mixtures of large clams and tube worms. Biogeographic studies depend on the number of vent fields explored, the methods and the underlying hypotheses used to delineate them. This has resulted in the North Atlantic being considered either as one biogeographic province (Bachraty et al. 2009; Moalic et al. 2012) or as two (Van Dover et al. 2002). 30 3.2.2 MAR-Vent systems: Gaps to our knowledge Meiofaunal diversity and biogeography is rather poorly studied at the MAR. Diversity is likely relatively low, with i.e. Sampling and sampling methodology 15 meiofauna species found associated with mussel beds at German et al. (2011), in summarising the next phase of Snake Pit (Zekely et al. 2006a), and ~20 species found research after the Census of Marine Life, estimated that associated with artificial substrates at Lucky Strike (Cuvelier only 20% of global mid-ocean ridges have been investigated et al. 2014). Most dominant taxa are dirivultid copepods, a and they calculated that as many as 1000 vents were yet to family which is endemic to the vent environment (Gollner et be discovered. It can be assumed that some of these will be al. 2010a), harpacticoid copepods and nematodes which on the MAR. In the recommendations for future work, both are represented by genera known from a variety of biogeography stands as a recurrent area of research for all other environments. Whilst only a single nematode species the hydrothermal regions. They also highlight the MAR is from MAR is taxonomically described (Zekely et al. 2006b), one of the regions in need of more research. The critical the knowledge on harpacticoid copepod species is a bit questions which remain to be answered focus on the better: Smacigastes micheli (Ivanenko et al. 2004, 2012, discovery and assessment of the distribution of the biota Cuvelier et al. 2014) was reported several times from Lucky associated with vents systems. It is clear that many gaps Strike, Bathylaophonte azorica was reported from Snake Pit remain even in this relatively well-studied ecosystem (for a and Lucky Strike (Zekely et al. 2006 a, Cuvelier et al. 2014), deep-sea habitat). and Xylora bathyalis was found at MAR vents and on East Large-scale geographic coverage As suggested above considerably more vents are thought to exist. Within the MAR system more surveys will provide sites between the known ones. This should provide more data on the distribution of key species. Sampling standardisation and effectiveness for biogeographic and connectivity studies connectivity. As commercial exploration activities increase the ISA (2004) have put forward standardised protocols which contractors are expected to follow. There is, however, no such agreement between research teams. Existing Data and Access Existing data are mostly within scientific literature, lab research reports, students’ theses. Generally available. Data are not widely available from later commercial exploration of the two claim areas Different teams tend to study vents for a range of reasons and even multidisciplinary studies tend to have a range of objectives. As a result standardised approaches to surveying, sampling and subsequent analyses are difficult. This generates a lag between the initial results and data which can be standardised for biogeographic and Taxonomic impediment The charismatic nature of the large megafaunal organisms are usually well covered, although the lag between discovery and formal classification can still be quite long. As for other habitats, classification of the smaller biota often lags behind the sampling. As pointed out by Van Dover (2012) the rate of discovery of macrofaunal vent species shows no sign of levelling off even in well-studied sites. Taxonomic resources are not adequate to provide the necessary data quickly, and possibly not within the timeframe between exploration and exploitation unless better taxonomic pipelines are established. Molecular sampling Sampling for genetic analyses is now becoming a standardized approach. Some taxa lack any genetic data but these are becoming the exception. There are good protocols and standards now in place. 3.3 Non-active vents and non-vent hard substratum sites 3.3.1 Current Knowledge: Biogeography of major faunal groups Megafaunal biodiversity. Outside the direct influence of the vent fluids (inactive areas and the beyond vent fields), there are significant gaps in our knowledge, not only due to the lack of data, but also due to the patchiness of the habitats. Despite the North Atlantic being the best studied oceanic basin concerning deep-water benthic organisms (Cairns, 2007; Mironov & Gebruk 2006; Watling & Auster 2005), even in this region, data on the distribution of many megafaunal organisms is still scarce. This lack of information may result from: 1) the absence of sampling in most deep-sea areas because of high costs and technological challenges; 2) difficulties with identification of the species sampled or in collections; or 3) they may simply be naturally rare across their distribution range. The mega and larger macrofauna of the MAR have been assessed in a number of recent studies. Sampling in MAR is challenging and undersampling remains an issue. MAR ECO studied the region to the north of the Azores. Table 14 summarises the distribution patterns of the taxonomic groups reported. Most of the taxa have wide geographic ranges with the morphotypes being found in other areas of the Atlantic or extending into other oceans. The results from the MAR-ECO project indicate that despite decades of sampling new megafaunal species are still being discovered. From a relative low number of trawls one new genus of echinoid, four new species of holothurian, four of ophiuroids and one new asteroid species were discovered. Macrofauna Little is known, of fauna of the sponge aggregations and coral gardens and coral framework. There are some recent data from the Condor seamount in the Azores (Bongiorni et al. 2013, Zeppilli et al. 2013) but with no supporting molecular data information. For example, within the group of epizoan zoanthids associated with octocorals, most information available in the Atlantic Ocean comes from shallow water, but recent studies have revealed the existence of an hidden diversity in the deep-sea with the identification of several new genera and species (Carreiro-Silva et al. 2011; Carreiro-Silva et al in prep). The macrofauna associated with the sediment found between ridges along the MAR were studied by Shields and Blanco-Perez (2013). They reported that the MAR did not appear to present a physical barrier to the distribution of macrofauna and that the diversity was similar to the continental margins are comparable depths. However, with a few notable exceptions, most of the species were identified to morphological OTUs and so links of this MAR fauna to the continental margins or abyssal plains remains unknown. Meiofauna Very few papers have been produced comparing hard surfaces with other environments or between seamounts and surrounding sediments. In a recent paper Zepilli et al. (2013) reported urgent research was needed on this topic. 31 KNOWLEDGE GRID for MID ATLANTIC RIDGE - Vent systems Meiofauna Microbial: Protista Microbial Bacteria Microbial Archaea Good general knowledge of species Generally good resolution Limited to a few taxa (M)OUT common Limited MOTU – both free-living and symbiotic MOTU Keys, etc Available for many groups. Monographic treatment/fieldguide Available for some groups None None N/a N/a Taxonomic information availability Scientific literature, on-line databases Scientific literature, on-line databases Limited information ? Data available via databases such as GenBank Data available via databases such as GenBank Sampling Quantitative ad qualitative; video surveys Quantitative and qualitative samples Quantitative samples Quantiitative Quantitative Quantiative Molecular sampling Almost standard Becoming common Becoming common ? All molecular eDNA All molecular - eDNA Consistent sampling between different teams Widespread acceptance of sampling methodology Widespread acceptance of sampling methodology Widespread acceptance of sampling methodology ? Limited but follows accepted scientific standards Limited but follows accepted scientific standards Sythensis available but more spatial coverage would be useful. Some but scattered in scientific literature. Lacking sufficient spatial coverage Assessment for some groups – Harpacticoids but others lacking sufficient spatial coverage? Some. Coverage? Biogeographic data 32 Macrofauna Taxonomic resolution Sampling Taxonomy Megafauna Some . Coverage? 3.3.2 MAR – non-vent and off vent environments: Gaps to knowledge Sampling and sampling methodology The heterogeneity of the environment make standardised approaches difficult – different types of trawl and semi-quantitative gears are used. Similarly differences in the sampling programme and strategies using ROV and AUVs between different research and contracting teams make existing data held by institutes available. To some extent this is being done but specimen records in Museums and other institutes are often not available. Releasing this data or digitising such records would provide another source of information about species level biogeography. The heterogeneity of the MAR non-vent sites is a major challenge to collecting. Traditional methods using quantitative samplers such as corers Table 14. The number of species collected during the MAR-ECO project and an assessment of their are very difficult to deploy successfully. biogeographical distribution. The numbers in the body of the table represent the number of species of each Semi-quantitative methods such as trawls taxonomic group. have been used in projects such as Also in MAR-ECO and ECO-MAR but such gears only NW MAR only N. Atlantic NE Atlantic other Atlantic collect megafauna and often the condition oceans of the samples are poor. Hexactinellida1 ROVs are frequently deployed in rock areas and can provide valuable specimens and quantitative samples but the numbers of ROVs are limited, the areas surveyed and sampled can be quite small compared with the size of the environment and navigating among the complex topography encountered can be challenging. Nevertheless they represent to optimum way to sample the smaller elements of the biota. Carefully planned sampling strategies together with extensive collecting are probably the optimum way to provide the data on species distributions along the MAR. Large-scale geographic coverage Anthozoa2 Hexacorallia Antipatharia Scleractinia Octocorallia Echinodermata Asteroidea3 Ophiuroidea4 Holothuroidea5 Echinoidea6 7 1 3 2 1 1 3 4 1 1 2 4 3 16 14 5 4 1 2 1 1 1 4 6 7 6 Taxonomic coverage is problematic as mentioned above. Molecular sampling Considerably more molecular samples need to be collected as part of sampling programmes. Larger specimens need to have molecular subsamples taken. The specimens need to be imaged both in situ (if taken by ROV) and ‘on deck’. Sediment samples need to be processed on ship and the infauna imaged, subsampled for molecular analyses or if large enough fragments taken and frozen or preserved in absolute ethanol for future analyses. 1 Microbiology Tabachnick & Collins (2008); 2 Molodtsova et al. (2008); 3 Dilman (2008); 4 Martynov & Litvinova (2008); 5 Gebruk (2008); 6 Mironov (2008) 1 There are gaps in the coverage along the MAR and particularly off-vent areas. Historical records tend to be concentrated around the fishing areas such as seamounts around the Azores (see Moldosova, 2007; Braga Henriques et al. 2013 for a review). Sampling standardisation and effectiveness for biogeographic and connectivity studies 3 Taxonomic impediment make standardisation difficult. There are various examples of best practice which could be adopted if agreement on over all data needs was made. Protista. Very little is known about protistan diversity from hard surfaces as they are difficult to sample. No published data could be retrieved. Bacteria and Archaea. No literature sources were found for this biota. Existing Data and Access Historical data is being collated for some regions such as the Azores but more ‘data archaeology’ could be carried out to 33 Cnidarian megafauna–Case study 3 The current knowledge of the taxonomic diversity and distribution patterns of octocorals is scarce and fragmented, compounded by the lack of taxonomic expertise (Riedel et al, 2013]. The North Atlantic Ocean is one of the most comprehensively studied regions (Watling & Auster, 2005; Cairns, 2007), For example, knowledge on octocoral taxonomy is not uniform on both sides of the Atlantic. Most recent octocoral taxonomic studies have focused on the northwestern Atlantic (Gulf of Mexico, Florida Strait), resulting in the description of several new species [e.g. Quattrini et al, 2014]. By contrast, the octocoral fauna of the northeastern Atlantic (Mid Atlantic Ridge, Porcupine Seabight, Rockall Trough), and Mediterranean (e.g. Alboran Sea, Silicily Channel) have received less attention, with most taxonomic inventories dating back to the early 1990’s (Grasshoff, 1986; Vafidis et al. 1994). Moldosova (2008) suggested that the Mar-Eco data 40.6% of the fauna was widely distributed beyond the Atlantic (see Table 14). In the Central Atlantic (Azores), a recent study has identified 164 species in seamounts and island slopes, but there were a large number of octocorals with uncertain taxonomic affinity (Braga Henriques et al, 2013). Underlying factors influencing the regional distribution are complex and have not been clearly identified for the MAR. There are both latitudinal and bathymetric patterns in the corals around the Azores (Braga Henriques, 2013). The majority of records were concentrated in 100–600 m. Factors such as hard substratum availability, hydrography and productivity were hypothesised to play a part. The highest diversity found coincided with important fishing areas suggesting that both habitat availability and productivity are central to local to regional distributions. Modelling of coral distributions using similar approaches to Titttensor et al. (2009) around the Azores and along the MAR may be useful in developing predictions as to the impact of mining MSD off-vent areas. Most of the studies that have been done on the reef-building coral Lophelia pertusa (Le Goff-Vitry et al, 2004) have focused on the northern seas around Norway and Scotland. Some genetic population studies have been done comparing the northern fjords with the east Atlantic offshore areas. It was shown that there was a high genetic differentiation between fjord subpopulations and offshore ones. For this species associated microorganism have also been subject of recent studies showing that coral skeleton surface, coral mucus, ambient seawater and reef sediments present clear habitat-specific differences in community structure and species richness based on operational taxonomic unit (OTU) numbers on the Norwegian communities (Schöttner et al, 2009). To the best of our knowledge the few studies of beta diversity for this species have been done on populations from western Scotland (Henry et al. 2009). There is little information on the distribution at the global scale of the octocoral genera Corallium (Sampaio et al, 2009). Corallium is best known as species exploited to provide jewelry but only certain species have a commercial value. Most Corallium species live in restricted geographical areas. The Western and Central Pacific (from Japan to northern Philippines and Hawaii) and the Northeast Atlantic are hotspots for these gorgonians (Sampaio et al, 2009). Corallium rubrum Lamarck, 1816 in the Mediterranean and other seven species from the Western Pacific and Hawaii form large aggregations in the upper bathyal (to 500 m) and support local fisheries. amphiatlantic C. niobe Bayer, 1964 and C. medea Bayer, 1964, which is restricted to the American slopes. In this context, the Macaronesian deep-sea rocky bottoms emerge as a probable radiation centre for these species. As for other cold water corals, Corallium spp. are highly sensitive to anthropogenic disturbances: they have slow growth rates, relatively late maturity, long life spans, and limited dispersal potential). A better understanding of the factors which underlie the distribution of species of Corallium is needed together with surveys to determine how common it is in different areas associated with off-vent mineral sites. Outside this subtropical belt, there are records from off Ireland and the Bay of Biscay ( Grasshoff 1982b, 1986). Apparently, these species do not aggregate, are uncommon throughout the area, and their biology is virtually unknown. In the western Atlantic the genus is less diversified, and according to Watling & Auster (2005) only two species are known from that region: the Octocorals and scleratinians from the MAR.Left: Colony of Corallium tricolor from Menez Gwen marine protected area. Middle: Dense colonies of Madrepora oculata and Lophelia pertusa with a rockfish. Right: Trachyscorpia cristata echinata. (Images courtesy of IMAR/DOP) 34 KNOWLEDGE GRID for MID-ATLANTIC RIDGE - Non-vent environments Sampling Taxonomy Megafauna Macrofauna Meiofauna Microbial Protista Microbial Bacteria Microbial Archaea Taxonomic resolution Good general knowledge of species Generally good resolution but OTUs from sediment regions Limited to a few taxa (M)OTUs common Limited MOTU – MOTU Keys, etc Available for many groups. Monographic treatments for some taxa Available for some groups None None N/a N/a Taxonomic information availability Scientific literature, on-line databases for some taxa Scientific literature, on-line databases Limited information ? Limited information Limited information Sampling Quantitative, video surveys Quantitative and qualitative samples Quantitative samples Limited samples available Limited Limited Molecular sampling Limited Becoming common Becoming common ? All molecular - eDNA All molecular - eDNA Consistent sampling between different teams Sampling variable between studies Widespread acceptance of sampling methodology Widespread acceptance of sampling methodology Limited but follows accepted scientific standards Limited but follows accepted scientific standards Some synthesis for some taxa but spatial coverage variable - most information from northern MAR Some but scattered in scientific literature. Lacking spatial coverage Insufficient coverage Insufficient coverage Insufficient coverage Biogeographic data Insufficient coverage? 35 3.4 Methane hydrate – Svalbard margin and Black Sea 3.4.1 Svalbard margin. These seep sites have only recently been discovered so there is relatively little biological information available (e.g. dos Santos et al. 2009), most of the reports focused on the geological setting (e.g. Bunz et al. 2012). While initial surveys indicated that sites along the Svalbard margin are likely to be similar to those on the Norwegian margin and the well-studied Håkon Mosby mud volcano site, seep sites do have faunistic differences (Vanreusel et al. 2009). Such differences relate to the oceanographic, geochemical settings and the depth of the seep. As a result there is a limited amount of detailed biogeographical assessment that can be made at this point. The priority for future research should be to undertake general survey which will place this region into context with the other areas within the Arctic. 36 Figure 7 below images of gas hydrate fields from the west of Salbard (78°35’N, 9°40‘E) and from the Håkon Mosby Mud Volcano Left. At a water depth of 380m off western Svalbard – the upper limit of the gas hydrate stability zone in the area – methane is released and seeps from the seafloor. The methane itself or sulfide generated upon the anaerobic oxidation of methane (AOM) in the sediments fuels biomass production by chemosynthetic microorganism. Gutless siboglinid tubeworms hosting chemoautotrophic symbionts appear in dense colonies and are partly covered by filamentous sulfide oxidizing bacteria. Other members of the benthic community depicted in the image include sea spiders and bivalves. Expedition HE387 of the German RV HEINCKE, August/September 2012. Image courtesy of MARUM – Center for Marine Environmental Sciences, University of Bremen, Germany. Midle. At a water depth of 380m off western Svalbard – the upper limit of the gas hydrate stability zone in the area – methane is released and seeps from the seafloor. The methane itself or sulfide generated upon the anaerobic oxidation of methane (AOM) in the sediments fuels biomass production by chemosynthetic microorganism. Gutless siboglinid tubeworms hosting chemoautotrophic symbionts colonizes soft sediments between boulders and cobble stones next to whitish mats of sulfide oxidizing bacteria. Anemones and gadid fish are also visible in the image. Expedition HE387 of the German RV HEINCKE, August/September 2012. Image courtesy of MARUM – Center for Marine Environmental Sciences, University of Bremen, Germany. Top right. A sea spider living on methane-rich muds emitted from the subsurface at the Håkon Mosby Mud Volcano (HMMV) at 1250m water depth. Expedition ARKXXIV/2 of the German RV POLARSTERN, July 2009. Image courtesy of MARUM – Center for Marine Environmental Sciences, University of Bremen, Germany. Bottom right. Muds and fluids with high concentrations of methane from deep subsurface reservoirs are emitted at the Håkon Mosby Mud Volcano (HMMV) at 1250m water depth. Whitish mats of thiotrophic bacteria that oxidize sulfide generated by anaerobic oxidation of methane in the sediments appear at the surface. It appears that also non-seep organisms, e.g., zoarcid fish and starfish benefit from this chemosynthetic primary productivity. Expedition ARKXXIV/2 of the German KNOWLEDGE GRID for SVALBARD MARGIN Megafauna Taxonomy Meiofauna Microbial : Protista Microbial Bacteria Microbial Archaea Mostly dominant species , MOTU and OTU MOTUs MOTUs N/a N/a Resolution to species possible Resolution to species possible Resolution at family and genus level is good but species is poor Keys, etc Some available Russian keys to many groups. Other primary works and keys to particular taxa available Family and genus level. Few species level Taxonomic information availability Good background knowledge of Arctic Megafauna species available Good background knowledge of Arctic macrofauna species. But new species possible Mostly primary sources Mostly primary sources Gene sequences on external databases Gene sequences on external databases Sampling Specimens needed as well as video/still imaging Quantitative and qualitative at different spatial scales needed Quantitative at different spatial scales Quantitative at different spatial scales Quantitative at different spatial scales Quantitative at different spatial scales Molecular sampling Essential Essential Essential Essential Essential Essential Good background available to place sites into context of Arctic biogeograph Increasing information to produce contextual information Synthesis is needed based on more taxonomic work More needed More needed More needed Taxonomic resolution Sampling Macrofauna Biogeographic data Little available 37 3.4.2 Black Sea The deep zones of the Black Sea are unique in the World Ocean in that they are totally anoxic below 100 to 200 m. The Deep Water Mass is the domain of anaerobic processes dominated by bacteria and archaea (Kuypers et al. 2003; Durisch-Kaiser et al. 2005). Surveys of the microbial systems were undertaken during the EU HERMES project (Foucher et al. 2009). A large range of microbial communities was analysed. The discovery of carbonate reefs indicates some of the unique aspects of this deep-sea system (Arnds et al. 2007). Microbial diversity was heterogeneous in the reefs with different parts having distinct diversities. The Archaea involved in this process were anaerobic methane oxidising groups but sulphur-reducing bacteria were also present. There also did not seem to be any consistency in community composition between the reefs; each being a mixture of the main species. A great deal more study of this remarkable ecosystem is needed. Microbial biogeography as it relates to ecosystem functioning is a priority. What are the connections between the sulphur and methane reducers in this system with those found in vents and Figure 9 (left and right) Carbonates formed upon anaerobic oxidation of methane (AOM) by consortia of sulfate reducing bacteria and archaea build large reef structures at 220m depth at the northwestern Crimean shelf. While the microbial communities fueled by gas seepage from the seafloor form extremely high biomasses within the up to four meter high chimney structures, any metazoan life is missing due to the absence of oxygen in Black Sea waters at this depth. Expedition M72/2 of the German RV METEOR, March 2007. Images courtesy of MARUM – Center for Marine Environmental Sciences, University of Bremen, Germany. 41 seeps? Generally, microbial biogeography depends on the sampling. Various assessments indicate that several isolates have wide distributions. How much this is a result of limited taxon sampling remains to be determined. All ecosystems studied indicate that isolates with wide distributions are the result of functional congruence rather than biogeography. 4. Modelling The logistical difficulties, expense and vast areas associated with deep-sea sampling leads to gap in the knowledge of faunal distribution. One approach to fill these gaps on the spatial distribution of species is the species distribution model (SDM). This approach correlates species occurrence (presence-only or presence–absence) records with environmental data in geographic space to explain and predict a species’ distribution. A wide variety of SDM methods have now been developed, including correlative and mechanistic approaches. Each approach has advantages and disadvantages, but the vast majority of studies to date have been correlative. Such models were first applied in the terrestrial domain where their use has increased rapidly over the past 20 years and is now applied in conservation biogeography. By comparison, application of SDMs to marine species is rare, although interest in their application is increasing. Their use has been recently advocated in the support of marine spatial planning . In the deep sea, SDMs are increasingly used but mainly to predict the distribution of Cold Water Corals (CWC) . The emphasis given to CWC can be explained by three main reasons: i) they are recognized as vulnerable, threatened or ecologically significant by a number of regional or international agreements ; ii) taxonomy is consistent and presence data are widely available in open-access databases and iii) environmental data that best explain their occurrences such as temperature, aragonite saturation state, primary productivity or bathymetry and its derivatives are available at global scale . The resolution of the environmental dataset however is a major restriction to the reliability and applicability of cold water coral SDMs. The first global scale models to be developed had a resolution of about 100 km. The best predictors for CWC occurrences were temperatures, oxygen, aragonite saturation state and enhanced primary productivity thus providing a global envelope for potential environmental niches. Regional to local scale models now have a resolution on the order of 10 m or even less . In these models, bathymetric terrain attributes have shown good potential as environmental predictors as they act as proxies indicating areas of enhanced currents and food supply for suspension-feeding corals , narrowing down the footprint of the potential niche. In the framework of the environmental management of resource exploitation in the deep sea, SDMs such as those developed for CWC may prove useful to fill gaps in the distribution of megafaunal assemblages associated with inactive vents or cobalt crusts as these environments are potentially suitable for sessile suspension feeders including CWC. The reliability of such models would be constrained by the availability of data on the occurrence of species in the area to be modeled as well as the resolution of the bathymetry. Indeed, SDMs have proven powerful when they are used for interpolation (i.e. to fill in the geographical gaps in species distribution within an area where occurrences are known) but the most commonly used correlative SDMs are limited in their ability to extrapolate species distribution across space or time . For example, modeling the distribution of species along the mostly unknown southern Mid-Atlantic Ridge should not rely on correlative models developed from the better known northern Mid-Atlantic Ridge. Modeling the distribution of species in nodule provinces would further face three major issues: i) taxonomically robust datasets are limited for most taxonomic groups and nodule provinces; ii) SDMs would only apply to the most common and widespread species but not the bulk of rare species for which no correlation with environmental factors could be drawn; and iii) the environmental drivers for species distribution are not well constrained nor are the influence of functional traits such as dispersal ability or the phylogeography of species. These later biotic drivers of species distribution are not considered in correlative models but the development of processus-based or mechanistic approaches coupled with correlative models are promising tools. Species distribution models are thus valuable tools to fill gaps in the known distribution of species, which are increasingly used in conservation biogeography. As any model they may provide insights into the most critical gaps in knowledge that need to be filled through targeted studies and as any model their reliability is highly constrained by the quality and resolution of input data, both biological and environmental. 5. General discussion The known The information gained by over a century of research and discovery has produced data on the identification and distribution of many large megafaunal organisms. The revelation of high species diversity in the deep-sea sediments has been verified across the World Ocean. While the discovery of chemosynthetic systems with their associated large, abundant megafauna has resulted in focused study and exploration of such systems. In chemosynthetic systems taxonomic knowledge has almost been able to keep up with the discovery of new vents and seeps. The factors driving the local and to some extent the regional distribution patterns have been established. But for the other high diversity environments and for the smaller sized biota taxonomic and biogeographic knowledge have not been able to keep pace with the rate of discovery or sampling. The drivers of local and regional diversity 42 patterns have been hypothesised but a robust ecological framework is still being developed. The unknown contractors and research groups. However, time is short and the resources being deployed are not always commensurate with the effort needed. Providing sufficient resources is important to concentrate effort on the key gaps needed to be filled before irretrievable change occurs. Despite considerable sampling in the deep sea over the past century, our knowledge of the distribution of organisms at To fully understand the faunal composition of the region, different scales is rudimentary. Certainly, very few areas the links and connectivity both within and between areas have been studied sufficiently to be able to assert with any within potential mining areas have to be established as part degree of confidence what the impact of mining will be on of future sampling programmes. the survival of species. The scale of the problem is mirrored by the vast size of the environment. Yet there is an The unknowable opportunity to address many of the key questions by focussed biological sampling programmes, use of new Deep-sea sediment samples are characterised by a long tail genetic technologies and greater collaboration between of species with only one or possibly two individuals which have not been recorded from other sites and often found in a single sample. Quantitative samples are particularly conspicuous in this regard. The presence of rare species appears to be an emergent property of deep-sea systems and is not just a function of sampling efficiency. It is therefore unlikely that we will ever be able to know the distribution of these species and whether they are in danger of extinction due to mining and mineral extraction. Modelling, improved genetic sampling and co-ordinated, nested sampling programmes may go some way to reducing the risk but such approaches will not totally solve this problem. Ultimately it will be to policy makers and regulators to make the decision as to allow exploitation. 6. Conclusions We have been studying the deep-sea ecosystem for over a century and in that time have made significant advances in our understanding of the fauna and ecology of this part of the planet. Yet the knowledge we have is not enough to be able to predict, let alone guess, what the long-term consequences of extracting mineral resources will be. There are several reasons which explain our apparent ignorance despite all the efforts that have been made to explore and explain this ecosystem. The most obvious one is the sheer scale of the ecosystem itself. Efforts to date have only literally scratched the surface of a very small area. It is also a difficult environment to sample, requiring good technologically-advanced, ocean-going vessels to deploy scientists and collecting gear. Thus sampling programmes tend to be focussed and of limited duration. The kinds of surveys which were common in the early days of deep-sea 43 research and championed by the Soviet programmes of the 1950s and 60s are no longer fundable. Collections of biota are therefore usually from surveys of particular areas to address particular scientific questions. Sample coverage is as a result very patchy and subsequently it is difficult to make extrapolations from such small sample sets. This change in focus is a reflection of the way science is funded and carried out. The consequence for biogeographic studies has been that taxonomy, the description and classification of species, has become more difficult to fund. There have been exceptions in recent years associated with the novelty of seep and vent species but the numbers involved are actually relatively low in comparison with the high species richness to be found in sediment systems. So it is relatively easy for the taxonomic effort to keep pace with the rate of discovery (although there is a backlog even among seep and vent fauna). The size of the task facing taxonomists in nodule fields and off vent areas is a magnitude larger but the resources to support this work available are actually less. The future will depend on maximising the resources we have, increasing the co-ordination between industry and academia so that the most is made of the efforts currently underway and being planned. 12 Dec 2014 6. References Ahnert, A., Schriever, G., 2001. Response of abyssal Copepoda Harpacticoida (Crustacea) and other meiobenthos to an artificial disturbance and its bearing on future mining for polymetallic nodules. Deep-Sea Research II, 48, 3779–3794. Almeida-Neto, M., Guimaraes, P., Guimaraes, P.R., Jr., Loyola, R.D., Ulrich, W., 2008. A consistent metric for nestedness analysis in ecological systems: reconciling concept and measurement. Oikos, 117 (8), 1227-1239. Arnds, J., Knittel, K., Boetius, A., Amann, R., 2007. Black Sea shelf microbial reefs. Geochimica Et Cosmochimica Acta 71 (15), A36-A36. Bachraty C., Legendre P., Desbruyères D. 2009. Biogeographic relationships among deep-sea hydrothermal vent faunas at global scale. Deep Sea Research I, 56, 1371–1378. Baselga, A. 2010. Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography, 19, 134–143 Bayer, F.M., 1964. The genus Corallium (Gorgonacea : Scleraxonia) in the western North Atlantic Ocean. Bulletin of Marine Science of the Gulf and Caribbean, 14 (3), 465-478. Beerli, P., Felsenstein, J, 1999. Maximum-likelihood estimation of migration rates and effective population numbers in two populations using a coalescent approach. Genetics, 152, 763-773. Bergstad, O.A., Falkenhaug, T., Astthorsson, O.S., Byrkjedal, I., Gebruk, A.V., Piatkowski, U., Priede, I.G., Santos, R.S., Vecchione, M., Lorance, P., Gordon, J.D.M., 2008. Towards improved understanding of the diversity and abundance patterns of the mid-ocean ridge macro- and megafauna. Deep-Sea Research Part II, 55 (1-2), 1-5. 41 Bergstad, O.A., Gebruk, A.V., 2008. Approach and methods for sampling of benthic fauna on the 2004 MAR-ECO expedition to the Mid-Atlantic Ridge. Marine Biology Research, 4 (1-2), 160-163. Cairns S.D. (2007) Deep-water corals: an overview with special reference to diversity and distribution of deepwater Scleractinian corals. Bulletin of Marine Science, 81, 311–322. Bongiorni, L,. Ravara, A., Parretti, P., Amaro, T., Cunha, M.R. 2013. Organic matter composition and macrofaunal diversity in sediments of the Condor Seamount (Azores, NE Atlantic). Deep-Sea Research II, 98, 75-86. Carreiro-Silva M., Braga-Henriques, A., Sampaio, I., Matos, V., Porteiro, F., Ocaña, O. 2011. Isozoanthus primnoidus, a new zoanthid species (Anthozoa: Hexacorallia) associated with the gorgonian Callogorgia verticillata (Anthozoa: Octocorallia) in the Azores. ICES Journal of Marine Science, 68: 408-415. Braga-Henriques, A., Porteiro, F. M., Ribeiro, P. A., de Matos V., ´ Sampaio, I., Ocaña O., and Santos, R. S. 2013 Diversity, distribution and spatial structure of the cold-water coral fauna of the Azores (NE Atlantic). Biogeosciences, 10, 4009–4036. Brault, S., Stuart, C.T., Wagstaff, M.C., McClain, C.R., Allen, J.A., Rex, M.A., 2013a. Contrasting patterns of alpha- and beta-diversity in deep-sea bivalves of the eastern and western North Atlantic. Deep-Sea Research Part II, 92, 157-164. Brault, S., Stuart, C.T., Wagstaff, M.C., Rex, M.A., 2013b. Geographic evidence for source-sink dynamics in deep-sea neogastropods of the eastern North Atlantic: an approach using nested analysis. Global Ecology and Biogeography 22 (4), 433-439. Brown, C.J. 1998. Effects of phytodetriatal input on nematode communities of the abyssal equatorial Pacific. PhD Thesis, University of Southampton, UK. 302pp. Bunz, S., Polyanov, S., Vadakkepuliyambatta, S., Consolaro, C., Mienert, J., 2012. Active gas venting through hydrate-bearing sediments on the Vestnesa Ridge, offshore W-Svalbard. Marine Geology, 332, 189-197. Bussau, C., Schriever, G., Thiel R., 1995) Evaluation of abyssal metazoan meiofauna from a manganese nodule area of the eastern south Pacific. Vie Milieu, 45(1), 39-48. Carreiro-Silva M., Ocaña O, Sampaio I, Matos V, Porteiro F Braga-Henriques A, Steffani (in prep) Zoanthid diversity associated with cold-water corals in the Azores Region: evidence for substrate specificity. Marine Biodiversity CBD, 2009. Azores Scientific criteria and guidance for identifying ecologically or biologically significant marine areas and designing representative networks of marine protected areas in open ocean waters and deep sea habitats. Convention on Biological Diversity, p. 12. Collins, P.C., Kennedy, R., Van Dover, C.L., 2012. A biological survey method applied to seafloor massive sulphides (SMS) with contagiously distributed hydrothermal-vent fauna. Marine Ecology Progress Series, 452, 89-107. Comtet, T., Jollivet, D., Dixon, D.R. 2000. Molecular and morphological identification of settlement-stage vent mussel larvae, Bathymodiolus azoricus (Bivalvia: Mytilidae), preserved in situ at active vent fields on the Mid-Atlantic Ridge. Limnology and Oceanography, 45 (7). 1655-1661. Crepeau, V.; Bonavita, M-A. C., Lesongeur, F.; et al. Diversity and function in microbial mats from the Lucky Strike hydrothermal vent field. FEMS Microbiology Ecology, 76 (3) 524-540 Cuvelier D., Beesau, J., Ivanenko, V.N., Zeppilli D., Sarradin, P.M., Sarrazin, J. (2014) First insights into macro-and meiofaunal colonisation patterns on paired wood/slate substrata at Atlantic deep-sea hydrothermal vents. Deep Sea Research Part I, 87:70-81. Davey, J.W., Hohenlohe, P.A., Etter, P.D., Boone, J.Q., Catchen J.M., Blaxter, M.L., 2011. Genome-wide genetic marker discovery and genotyping using next-generation sequencing. Nature Reviews Genetics, 12, 499-510. Desbruyères, D., Olu, K., Colaco, A., Comtet, T., Sarradin, P.-M., Biscoito, M., Fouquet, Y., 2001. Composition and distribution of communities associated with oceanic hydrothermalism at the Azores Triple Junction in the Mid Atlantic. Boletim do Museu Municipal do Funchal, Suplemento, 6, 75-110. Dilman, A.B., 2008. Asteroid fauna of the northern Mid-Atlantic Ridge with description of a new species Hymenasterides mironovi sp nov. Marine Biology Research, 4 (1-2), 131-151. Durisch-Kaiser, E. Klauser, L., Wehrli, B. & Schubert, C. 2005. Evidence of intense archaeal and bacterial methanotrophic activity in the black sea water column. Applied and Environmental Microbiology, 71(12), 8099-8106. Foucher, J.-P., Westbrook, G.K., Boetius, A., Ceramicola, S., Dupre, S., Mascle, J., Mienert, J., Pfannkuche, O., Pierre, C., Praeg, D., 2009. Structure and Drivers of Cold Seep Ecosystems. Oceanography, 22 (1), 92-109. Gaston, K. J. 1994. Rarity. Chapman & Hall, London. 205pp. Gebruk, A.V., 2008a. Benthic fauna of the northern Mid-Atlantic Ridge: results of the MAR-ECO expedition. Marine Biology Research, 4 (1-2), 1-2. Gebruk, A.V., 2008b. Holothurians (Holothuroidea, Echinodermata) of the northern Mid-Atlantic Ridge collected by the GO Sars MAR-ECO expedition with descriptions of four new species. Marine Biology Research, 4 (1-2), 48-60. Gebruk, A.V., Budaeva, N.E., King, N.J., 2010. Bathyal benthic fauna of the Mid-Atlantic Ridge between the Azores and the Reykjanes Ridge. Journal of the Marine Biological Association of the United Kingdom, 90 (1), 1-14. German, C.R., Parson,L.M., Bougault,F., Coller,D., Critchley,M., Dapoigny,A., Day, C., Eardley,D., Fearn,A., Flewellen,C., Kirk,R., Klinkhammer,G., Landure,J.Y., Ludford, E.,Miranda, M.,Needham,H.D., Patching,J., Pearce,R., Pelle,H., Radford-knoery, J., Rouse,I., Scott,J., Stoffregen,P., Taylor,P., Teare,D., Wynar,J., 1996. Hydrothermal exploration near the Azores triple junction: tectonic control of venting at slow-spreading ridges? Earth and Planetary Science Letters, 138, 93–104. German, C.R., Ramirez-Llodra, E., Baker, M.C., Tyler, P.A. and the ChEss Scientific Steering Committee, 2011. Deep-Water Chemosynthetic Ecosystem Research during the Census of Marine Life Decade and Beyond: A Proposed Deep-Ocean Road Map. PLoS ONE, 6(8): e23259. doi:10.1371/journal.pone.0023259. Glover, A., Paterson, G., Bett, B., Gage, J., Sibuet, M., Sheader, M., Hawkins, L., 2001. Patterns in polychaete abundance and diversity from the Madeira Abyssal Plain, northeast Atlantic. Deep-Sea Research Part I, 48 (1), 217-236. Glover, A., Smith, C., Paterson, G., Wilson, G., Hawkins, L., Sheader, M. 2002. Polychaete species diversity in the central Pacific abyss: local and regional patterns, and relationships with productivity. Marine Ecology-Progress Series, 240, 157-169. Gollner, S., Ivanenko, V.N., Martínez Arbizu P., Bright M. 2010a. Advances in taxonomy, ecology, and biogeography of Dirivultidae (Copepoda) associated with chemosynthetic environments in the deep sea. PLoS ONE, 5(8): e9801. doi:10.1371/journal.pone.0009801 Gollner, S., Riemer, B., Martínez Arbizu, P., Le Bris, N., Bright, M. 2010b. Diversity of Meiofauna from the 9°50’N East Pacific Rise across a gradient of hydrothermal fluid emissions. PLoS ONE, 5(8): e12321. doi:10.1371/journal.pone.0012321 Gollner, S., Fontaneto, D., Martínez Arbizu, P. 2011) Molecular taxonomy confirms morphological classification of deep-sea hydrothermal vent copepods (Dirivultidae) and suggests broad physiological tolerance of species and frequent dispersal along ridges. Marine Biology, 158:221-231. doi: 10.1007/s00227-010-1553-y Gooday, A. J., Hori, S., Todo Y., Okamoto, T., Kitazato H. & Sabbatini A. 2004. Soft-walled, monothalamous benthic foraminiferans in the Pacific, Indian and Atlantic Oceans: aspects of biodiversity and biogeography. Deep Sea Research Part I, 51, 33-53, Grasshoff, M., 1982. Die Gorgonaria, Pennatularia und Antipatharia des Tiefwassers der Biskaya (Cnidaria, Anthozoa). Ergebnisse der franzosischen Expeditionen Biogas, Polygas, Geomanche, Incal, Noratlante und Fahrten der ‘Thalassa’. 2. Taxonomischer Teil. Bulletin du Museum National d’Histoire Naturelle Section A Zoologie Biologie et Ecologie Animales, 3 (4), 941-978. Grasshoff, M. 1986. Die Gorgonaria de Expeditionen von “Travailleur” 1880_1882 und “Talisman” 1883 (Cnidaria, Anthozoa). Bulletin Muséum national d’Histoire Naturelle, Paris, 4e série, 8, 9-38. Grigg, R.W., 1994. History of the precious coral fishery in Hawaii. Precious Corals & Octocoral Research, 3, 1-18. Henry L.-A., Davies A.J., & Roberts J.M. 2010. Beta diversity of cold-water coral reef communities off western Scotland. Coral Reefs, 29,427-436 Hessler, R. R. and P. A. Jumars, 1974. Abyssal community analysis from replicate box cores in the central North Pacific, Deep-sea Research 21,185–209. 42 Ivanenko, V.N., Corgosinho, P.H.C., Ferrari, F.D., Sarradin, P.M., Sarrazin, J. 2012. Microhabitat distribution of Smacigastes micheli (Copepoda: Harpacticoida: Tegastidae) from deep-sea hydrothermal vents at the Mid-Atlantic Ridge, 37°N (Lucky Strike), with a morphological description of the nauplius. Marine Ecology Evolutionary Perspectiv,e 33(2), 245-256. Ivanenko, V.N., Defaye, D. 2004. A new and primitive genus and species of deep-sea Tegastidae (Crustacea, Copepoda, Harpacticoida) from the Mid-Atlantic Ridge, 37°N (Azores Triple Junction, Lucky Strike). Cahiers De Biologie Marine, 45, 255-268. Jain, M., Flynn, D.F.B., Prager, C.M., Hart, G.M., DeVan, C.M., Ahrestani, F.S., Palmer, M.I., Bunker, D.E., Knops, J.M.H., Jouseau, C.F., Naeem, S., 2014. The importance of rare species: a trait-based assessment of rare species contributions to functional diversity and possible ecosystem function in tall-grass prairies. Ecology and Evolution, 4 (1), 104-112. Keller, I., Wagner, C.E., Greuter, L., Mwaiko, S., Selz, O.M., Sivasundar, A., Wittwer, S., Seehausen, O. 2013. Population genomic signatures of divergent adaptation, gene flow and hybrid speciation in the rapid radiation of Lake Victoria cichlid fishes. Molecular Ecology, 22, 2848-2863. Kuypers, M.M. M, Slieker, A.O., Lavik, G, Schmid, M., Jorgensem, B.B., Kuenen, J.G., Sinninghe Damste, J.S., Strous, M., Jetten, Mike S.M. 2003. Anaerobic ammonium oxidation by anammox bacteria in the Black Sea. Nature, 422, 608-611. Lackschewitz, K.S., Endler, R., Gehrke, B., WallrabeAdams, H.J., Thiede, J., 1996. Evidence for topography- and current-controlled deposition on the Reykjanes Ridge between 59 degrees N and 60 degrees N. Deep-Sea Research Part I, 43 (11-12), 1683-1711. Lambshead, P.J.D., Brown, C.J., Ferrero, T.J., Hawkins, L.E., Smith, C.R., Mitchell, N.J., 2003. Biodiversity of nematode 43 assemblages from the region of the Clarion-Clipperton Fracture Zone, an area of commercial mining interest. BMC Ecology 3 (1), Unpaginated. Le Goff-Vitry, M.C., Pybus, O.G. & Rogers, A.D., 2004. Genetic structure of the deep-sea coral Lophelia pertusa in the northeast Atlantic revealed by microsatellites and internal transcribed spacer sequences. Molecular Ecology, 13, 537-549. Lecroq, B., Gooday, A.J., Cedhagen, T., Sabbatini, A. , Pawlowski, J., 2009a. Molecular analyses reveal high levels of eukaryotic richness associated with enigmatic deep-sea protists (Komokiacea). Marine Biodiversity, 39 (1), 45-55. Lecroq, B., Gooday, A.J. & Pawlowski, J., 2009b. Global genetic homogeneity in the deep-sea foraminiferan Epistominella exigua (Rotaliida: Pseudoparrellidae). Zootaxa, 2096, 23-32. Lecroq, B., Lejzerowicz, F., Bachar, D., Christen, R., Esling, P., Baerlocher, L., Osteras, M., Farinelli, L., Pawlowski, J., 2011. Ultra-deep sequencing of foraminiferal microbarcodes unveils hidden richness of early monothalamous lineages in deep-sea sediments. Proceedings of the National Academy of Sciences of the United States of America, 108 (32), 13177-13182. abiogenous origin. Instruments, Methods, and Missions for Astrobiology, Xiv, 8152. Magurran, A.E., Henderson, P.A. 2003. Explaining the excess of rare species in natural species abundance distributions. Nature, 422: 714-716. Mahatma, R., 2009. Meiofauna Communities of the Pacific Nodule Province: Abundance, Diversity and Community Structure. Ph.D. Thesis. Carl von Ossietzky Universit¨ at, Oldenburg, 142 pp. Martynov, A.V., Litvinova, N.M., 2008. Deep-water Ophiuroidea of the northern Atlantic with descriptions of three new species and taxonomic remarks on certain genera and species. Marine Biology Research, 4 (1-2), 76-111. Maybury, C. 1996. Crevice foraminifera from abyssal south east Pacific manganese nodules. Microfossils and oceanic environments, 281-295. Miljutin, D.M., Miljutina, M.A., Arbizu, P.M., Galeron, J., 2011. Deep-sea nematode assemblage has not recovered 26 years after experimental mining of polymetallic nodules (Clarion-Clipperton Fracture Zone, Tropical Eastern Pacific). Deep-Sea Research Part I, 58 (8), 885-897. Lejzerowicz, F., Voltsky, I. and Pawlowski, J, 2013. Identifying active foraminifera in the Sea of Japan using metatranscriptomic approach. Deep-Sea Research Part II, 86-87, 214-220. Miljutina, M.A., Miljutin, D.M., 2012. Seven new and four known species of the genus Acantholaimus (Nematoda: Chromadoridae) from the abyssal manganese nodule field (Clarion-Clipperton Fracture Zone, North-Eastern Tropical Pacific). Helgoland Marine Research, 66 (3), 413-462. Lichtschlag, A., Felden, J., Wenzhofer, F., Schubotz, F., Ertefai, T.F., Boetius, A., de Beer, D., 2010. Methane and sulfide fluxes in permanent anoxia: In situ studies at the Dvurechenskii mud volcano (Sorokin Trough, Black Sea). Geochimica Et Cosmochimica Acta, 74 (17), 5002-5018. Miljutina, M.A., Miljutin, D.M., Mahatma,R., Galéron, J. 2010. Deep-sea nematode assemblages of th Clarion-Clipperton Nodule Province (Tropical North-Eastern Pacific). Marine Biodiversity, 40, 1–15. Lysyuk, G. N. 2011. Biomineral microstructures in ferromanganese nodules: evidence of the biological and Mincks, S. L., Smith, C. R. 2006. Critical Review of Benthic Biological Data from the Clarion-Clipperton Zone (CCZ) and Adjoining Areas. Report to the ISA. Mironov, A.N., 2008. Pourtalesiid sea urchins (Echinodermata : Echinoidea) of the northern Mid-Atlantic Ridge. Marine Biology Research, 4 (1-2), 3-24. Mironov, A.N., Gebruk, A.V. 2006. Biogeography of the Reykjanes ridge, the northern Atlantic. In Mironova, A.N., Gebruk, A.V., Southward, A. J. eds. Biogeography of the North Atlantic Seamounts. KMK Scientific Press, Moscow.pp6-21. Moalic et al, 2012. Biogeography revisited with network theory: retracing the history of hydrothermal vent communities. Systematic Biology, 61(1),127–137, Molodtsova, T.N., Sanamyan, N.P., Keller, N.B., 2008. Anthozoa from the northern Mid-Atlantic Ridge and Charlie-Gibbs fracture zone. Marine Biology Research, 4 (1-2), 112-130. Morato, T., Pitcher, T.J., Clark, M.R., Menezes, G., Tempera, F., Porteiro, F., Giacomello, E., Santos, R.S., 2010. Can We Protect Seamounts for Research? A call for conservation. Oceanography, 23 (1), 190-199. Mouillot, D., Bellwood, D.R., Baraloto, C., Chave, J., Galzin, R., Harmelin-Vivien, M., Kulbicki, M., Lavergne, S., Lavorel, S., Mouquet, N., Paine, C.E.T., Renaud, J., Thuiller, W., 2013a. Rare Species Support Vulnerable Functions in High-Diversity Ecosystems. PLoS Biology 11 (5), 1-11. Mullineaux L.S. (1987) Organisms living on manganese nodules and crusts: distribution and abundance at three North Pacific sites. Deep Sea Research, 34, 165-184. Murton,B.J.,Klinkhammer,G.,Becker,K.,Briais,A.,Edge,D.,Hay ward,N.,Millard,N., Mitchell,I., Rouse,I., Rudnick,M., Sayanagi,K., Sloan,H., Parson,L.,1994. Direct evidence for the distribution and occurrence of hydrothermal activity between 27°-30°N on the Mid-Atlantic Ridge. Earth Planetary Science Letters,125,119–128. Naheem, S. 1998. Species redundancy and ecosystem reliability. Conservation Biology, 12, 39-45. Nimmo M, Morgan C and Banning D, 2013. Clarion-Clipperton Zone Project, Pacific Ocean: UPDATED NI 43-101 Technical Report. 127631013-003-R-Rev1. Nozawa, F., Kitazato, H., Tsuchiya, M., Gooday, A. J. 2006. ‘Live’ benthic foraminifera at an abyssal site in the equatorial Pacific nodule province: Abundance, diversity and taxonomic composition. Deep-Sea Research Part I, 53, 1406-1422. Priede, I.G., Bergstad, O.A., Miller, P.I., Vecchione, M., Gebruk, A., et al. 2013. Does Presence of a Mid-Ocean Ridge Enhance Biomass and Biodiversity? PLoS ONE, 8(5): e61550. doi:10.1371/journal.pone.0061550 Quattrini A.M., Etnoyer P.J., Doughty C., English L., Falco R., Remon N., Rittinghouse M., Cordes E.E. 2014. A phylogenetic approach to octocoral community structure in the deep Gulf of Mexico. Deep Sea Research Part II, 99, 92-102. R Development Core Team, 2006. O’mullan et al, 2001. A hybrid zone between hydrothermal vent mussels (Bivalvia: Mytilidae) from the Mid-Atlantic Ridge. Molecular Ecology, 10, 2819-2831 Radziejewska, T., 2002. Responses of deep-sea meiobenthic communities to sediment disturbance simulating effects of polymetallic nodule mining. International Review of Hydrobiology, 87 (4), 457-477. Pante, E,, Abdelkrim, J., Viricel, A., Gey, D., France, S.C., Boisselier, M-C., Samadi, S, online proof. Use of RAD sequencing for delimiting species. https://hal.archives-ouvertes.fr/hal-01064545/. Radziejewska, T. 2014. Meiobenthos in the sub-equatorial Pacific abyss. Springer, Hedelberg. 105pp. Paterson, G., Wilson, G., Cosson, N., Lamont, P. 1998. Hessler and Jumars (1974) revisited: abyssal polychaete assemblages from the Atlantic and Pacific. Deep-Sea Research Part II, 45 (1-3), 225-251. Pawlowski, J., Fontaine, D., da Silva, A.A., Guiard, J. 2011. Novel lineages of Southern Ocean deep-sea foraminifera revealed by environmental DNA sequencing. Deep-Sea Research Part II, 58 (19-20), 1996-2003. Pawlowski, J., Holzmann, M. 2008. Diversity and geographic distribution of benthic foraminifera: a molecular perspective. Biodiversity and Conservation, 17 (2), 317-328. Petersen, J.M., Ramette, A., Lott, C., Cambon-Bonavita, M.A., Zbinden, M., Dubilier, N. 2010. Dual symbiosis of the vent shrimp Rimicaris exoculata with filamentous Gammaand Epsilonproteobacteria at four Mid-Atlantic Ridge hydrothermal vent fields. Environmental Microbiology, 12, 2204–2218. Radziejewska, T., I. Drzycimski, V. V. Galtsova, L. V. Kulangieva and V. Stoyanova, 2001a. Changes in genus-level diversity of meiobenthic free living nematodes (Nematoda) and harpacticoids (Copepoda: Harpacticoida) at an abyssal site following experimental sediment disturbance, Proceeding of the Fourth Ocean Mining Symposium, Szczccin, Poland, September 23–27, pp. 38–43 Radziejewska, T. and I. Modlitba, 1999, Vertical distribution of meiobenthos in relation to geotechnical properties of deep-sea sediment in IOM pioneer area (Clarion-Clipperton Fracture Zone, NE Pacific), Proceeding of the Third Ocean Mining Symposium, pp. 126–130. Radziejewska, T., Stoyanova, V., 2000. Abyssal epibenthic megafauna of the Clarion-Clipperton area (NE Pacific): changes in time and space versus anthropogenic environmental disturbance. Oceanological Studies, 29 (2), 83-101. 44 Renaud-Mornant, J., Gourbault, N., 1990. Evaluation of abyssal meiobenthos in the eastern central Pacific (Clarion-Clipperton Fracture-Zone). Progress in Oceanograph,y 24 (1-4), 317-329. Riedel A., Sagata K., Suhardjono Y.R., Tänzler, R., Balke, M. 2013. Integrative taxonomy on the fast track – towards more sustainability in biodiversity research. Frontiers in Zoology, 10,15. Riemann, F. 1983. Biological Aspects Of Deep-Sea Manganese-Nodule Formation. Oceanologica Acta, 6, 303-311. Sampaio, I., Oca˜na, O., Tempera, F., Braga-Henriques, A., Matos, V., & Porteiro, F. New occurrences of Corallium spp. (Octocorallia, Coralliidae) in the Central Northeast Atlantic, Archipelago, Life Marine Science, 26, 73–78, 2009. dos Santos, E., Nolso, A., Schander, C., Tendal, O.S., Ragnarsson, S.A., Svavarrsson, J., 2009. Deep-water communities in the West-Nordic area. TemaNord, 529, 1-44. Schöttner, S. et al. Inter- and intra-habitat bacterial diversity associated with cold-water corals. ISME J. 3, 756–759 (2009). Schröder, C. J., Scott, D. B., Medioli, F. S., Bernstein, B. B. , Hessler, R. R. 1988. Larger agglutinated foraminifera comparison of assemblages from central north pacific and western north-atlantic (nares abyssal-plain). Journal of Foraminiferal Research, 18, 25-41. Shank, T. M., Halanych, K. M. 2007. Toward a mechanistic under-standing of larval dispersal: insights from genomic fingerprinting of deep-sea hydrothermal vent populations. Marine Ecology, 28, 25 – 35. Shields, M.A., Blanco-Perez, R., 2013. Polychaete abundance, biomass and diversity patterns at the 45 Mid-Atlantic Ridge, North Atlantic Ocean. Deep-Sea Research Part II, 98, 315-325. Smith, C. R., Berelson, W., Demaster, D. J., Dobbs, F. C., Hammond, D., Hoover, D. J., Pope, R. H. & Stephens, M. 1997. Latitudinal variations in benthic processes in the abyssal equatorial Pacific: control by biogenic particle flux. Deep-Sea Research Part II, 44, 2295-. Smith, C. R., Hoover, D. J., Doan, S. E., Pope, R. H., Demaster, D. J., Dobbs, F. C. & Altabet, M. A. 1996. Phytodetritus at the abyssal seafloor across 10 degrees of latitude in the central equatorial Pacific. Deep-Sea Research Part II, 43, 1309-1338. Smith CR, et al. (2008) Biodiversity, species ranges, and gene flow in the abyssal Pacific nodule province: predicting and managing the impacts of deep seabed mining.Technical Study No. 3. Kingston, Jamaica: International Seabed Authority.38pp Snider, L.J., Burnett, B.R., Hessler, R.R., 1984. The composition and distribution of meiofauna and nanobiota in a central north Pacific deep-sea area. Deep-Sea Research Part A, 31 (10), 1225-1249. Tabachnick, K.R. & Collins, A.G., 2008. Glass sponges (Porifera, hexactinellida) of the northern mid-Atlantic Ridge. Marine Biology Research, 4 (1-2), 25-47. Teixeira S., Cambon-Bonavita, M.-A. et al. 2011. Recent population expansion and connectivity in the hydrothermal shrimp Rimicaris exoculata along the Mid Atlantic Ridge. Journal of Biogeography, 38, 564-574. Teixeira S., Serra˜o E.A. & Arnaud-Haond, S. 2012. Panmixia in a fragmented and unstable environment: the hydrothermal shrimp Rimicaris exoculata disperses extensively along the Mid-Atlantic Ridge. PLoS ONE, 7(6): e38521. doi:10.1371/journal.pone.0038521 Thiel H., Schriever G., Bussau C., Borowski C. 1993. Manganese nodule crevice fauna. Deep Sea Research I, 40: 419-423. Tilot, V. 2006. Biodiversity and distribution of the megafauna. Vol 1. The polymetallic nodule ecosystem of the echinoderms of the Clarion-Clipperton fracture zone. eastern Equatorial Pacific Ocean. Vol 2. Annotated photographic atlas1995. An ecologically important type of biological activity in a deep-sea ferro-manganese nodule environment of the tropical north east Pacific. UNESCO/IOC (IOC Technical series 69). Tittensor, D.P., Baco, A.R., Brewin, P.E., Clark, M.R., Consalvey, M., Hall-Spencer, J., Rowden, A.A., Schlacher, T., Stocks, K.I., Rogers, A.D., 2009. Predicting global habitat suitability for stony corals on seamounts. Journal of Biogeography, 36 (6), 1111-1128. Tully, B. J. & Heidelberg, J. F. 2013. Microbial communities associated with ferromanganese nodules and the surrounding sediments. Frontiers in Microbiology, 4. Vafidis, D,, Koukouras, A., Voultsiadou-Koukoura, E. 1994. Octocoral fauna of the Aegean Sea with a check list of the Mediterranean species: new information, faunal comparisons. Annales de l’Institut oceanographique, Nouvelle serie, 70 (2), 217-229. van der Heijden K., Petersen, J.M., Dubilier, N., Borowski, C. 2012. Genetic Connectivity between North and South Mid-Atlantic Ridge Chemosynthetic bivalves and their symbionts. PLoS ONE, 7(7): e39994. Van Dover, C.L. 1988. Recruitment of marine invertebrates to hard substates at deep-sea hydrothermal vents onthe East Pacific Rise ad Galapagos spreading centre. Deep Sea Research, Part A. 35, 1833-1849. Van Dover, C.L. 2012. Hydrothermal vent ecosystems and consevation. Oceanography, 25, 313-316. VanDover, C.L., Desbruyeres, D., Segonzac, M., Comtet, T., Saldanha, L., FialaMedioni, A., Langmuir, C., 1996. Biology of the Lucky Strike hydrothermal field. Deep-Sea Research Part I, 43 (9), 1509-&. Van Dover, C.L., Doerries, M.B. 2005. Community structure in mussel beds at Logatchev hydrothermal vents and a comparison of macrofauna species richness on slow and fast-spreading mid-ocean ridges. Marine Ecology, 26, 110-120. Van Dover, C.L., German, C., Speer, K.G., Parson, L.M. & Vrijenhoek R.C. 2002. Evolution and biogeography of deepsea vent and seep invertebrates. Science, 295, 1253–1257. Vanreusel, A., Andersen, A.C., Boetius, A., Connelly, D., Cunha, M.R., Decker, C., Hilario, A., Kormas, K.A., Maignien, L., Olu, K., Pachiadaki, M., Ritt, B., Rodrigues, C., Sarrazin, J., Tyler, P., Van Gaever, S., Vanneste, H., 2009. Biodiversity of cold seep ecosystems along the European Margins. Oceanography, 22 (1), 110-127. Veillette, J., Juniper, S. K., Gooday, A. J., Sarrazin, J. 2007a. Influence of surface texture and microhabitat heterogeneity in structuring nodule faunal communities. Deep-Sea Research Part I, 54, 1936-1943. Veillette, J., Sarrazin, J., Gooday, A. J., Galeron, J., Caprais, J. C., Vangriesheim, A., Etoubleau, J., Christian, J. R., Juniper, S. K. 2007b. Ferromanganese nodule fauna in the Tropical North Pacific Ocean: Species richness, faunal cover and spatial distribution. Deep-Sea Research Part I, 54, 1912-1935. Verlaan, P. A. 1992. Benthic recruitment and manganese crust formation on seamounts. Marine Biology, 113, 171-174. Vierod, A.D.T., Guinotte, J.M., Davies, A.J., 2014. Predicting the distribution of vulnerable marine ecosystems in the deep sea using presence-background models. Deep Sea Research Part II, 99, 6-18. Vogler, A.P., Monaghan, M.T., 2007. Recent advances in DNA taxonomy. Journal of Zoological Systematics and Evolutionary Research, 45, 1-10. Vopel, K., Thiel, H., 2001. Abyssal nematode assemblages in physically disturbed and adjacent sites of the eastern equatorial Pacific. Deep-Sea Research Part II, 48, 3795–3808. Wang, c. S., Liao, L., Xu, H. X., Xu, X. W., Xu, M., Zhu, l. Z. 2010. Bacterial Diversity in the Sediment from Polymetallic Nodule Fields of the Clarion-Clipperton Fracture Zone. Journal of Microbiology, 48, 573-585. Wang, X., Schroeder, H. C., Schlossmacher, U., Mueller, W. E. G. 2009b. Organized bacterial assemblies in manganese nodules: evidence for a role of S-layers in metal deposition. Geo-Marine Letters, 29, 85-91. Watling, L., Auster, P.J. (2005) Distribution of deep-water Alcyonacea off the northeast coast of the United States. In Cold-Water Corals and Ecosystems (A. Freiwald and J. M. Roberts, eds), pp. 279_296. Springer-Verlag, Berlin. Wedding, L.M., Friedlander, A.M., Kittinger, J.N., Watling, L., Gaines, S.D., Bennett, M., Hardy, S.M., Smith, C.R.,2013. From principles to practice: a spatial approach to systematic conservation planning in the deep sea. Proceedings of the Royal Society B, Biological Sciences, 280 1471-2954. doi:10.1098/rspb.2013.1684 Willette, D.A., Allendorf FW, Barber PH, Barshis DJ, Carpenter KE, Crandall ED, Cresko WA, Fernandez-Silva I, Matz MV, Meyer E, Santos MD, Seeb LW and Seeb JE, 2014. So, you want to use next-generation sequencing in marine systems? Insight from the Pan-Pacific Advanced Studies Institute. Bulletin of Marine Science, 90, 79-122. Won, Y.-J., Hallam, S. J., O’Mullan, G.D., Pan, I. L., Buck, K. R., Vrijenhoek, R. C. 2003. Environmental Acquisition of Thiotrophic Endosymbionts by Deep-Sea Mussels of the Genus Bathymodiolus Applied Environmental Microbiology, 2003, 69(11),6785. Wu, Y.-H., Liao, L., Wang, C.-S., Ma, W.-L., Meng, F.-X., Wu, M., Xu, X.-W. 2013. A comparison of microbial communities in deep-sea polymetallic nodules and the surrounding sediments in the Pacific Ocean. Deep-Sea Research Part I, 79, 40-49. Xu, M. X., Wang, P., Wang, F. P., Xiao, X. 2005. Microbial diversity at a deep-sea station of the Pacific nodule province. Biodiversity and Conservation, 14, 3363-3380. Xu, M., Wang, F., Meng, J., Xiao, X. 2007. Construction and preliminary analysis of a metagenomic library from a deep-sea sediment of east Pacific Nodule Province. Fems Microbiology Ecology, 62, 233-241. Zekely, J., Van Dover, C.L., Nemeschkal, H.L., Bright, M. 2006a. Hydrothermal vent meiobenthos associated with mytilid mussel aggregations from the mid-atlantic ridge and the east pacific rise. Deep-Sea Research Part I, 53, 1363-1378. Zekely, J., Sørensen, M.V. & Bright, M. 2006b. Three new nematode species (Monhysteridae) from deep-sea hydrothermal vents. Meiofauna Marina, 15, 25-42. Zeppilli, D., Bongiorni, L., Cattaneo, A., Danovaro, R., Santos, R. 2013. Meiofauna assemblages of the Condor Seamount (North-East Atlantic Ocean) and adjacent deep-sea sediments. Deep-sea Research Part II, 98, 87-100. 46 12 December 2014