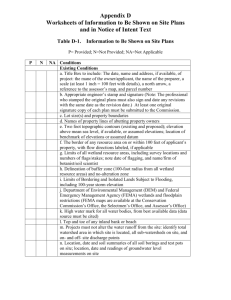
Wetland Drainage Effects on Prairie Water Quality Final Report Submitted by:
advertisement

Wetland Drainage Effects on Prairie Water Quality Final Report Agriculture Development Fund Project # 20070083 Submitted by: Cherie J. Westbrook1, Nathalie Brunet1, Iain Phillips2 and John-Mark Davies2 1 Centre for Hydrology, University of Saskatchewan, 117 Science Place, Saskatoon, SK S7N 5C8 phone: (306) 966-1818 email: cherie.westbrook@usask.ca 2 Saskatchewan Watershed Authority, #330-350 3rd Ave. N., Saskatoon, SK S7K 2H6 Centre for Hydrology Report No. 9 January, 2011 Table of Contents 1 2 3 4 Summary ................................................................................................................................. 6 Introduction............................................................................................................................. 8 Study Site .............................................................................................................................. 12 Methods................................................................................................................................. 13 4.1 Wetland water quality characterization ........................................................................ 13 4.2 Design of wetland drainage experiment ....................................................................... 14 4.3 Natural spills and artificial ditches sampling strategy .................................................. 15 4.4 Stream water quality sampling...................................................................................... 16 4.5 Analysis of water quality .............................................................................................. 17 4.6 Ecosystem health sampling sites and frequency........................................................... 18 4.7 Macroinvertebrate assemblage sampling methods ....................................................... 19 4.8 Habitat Assessment....................................................................................................... 20 4.9 Data analyses ................................................................................................................ 20 4.9.1 Characterization of wetland water quality ............................................................ 20 4.9.2 Wetland drainage experiment ............................................................................... 21 4.9.3 Comparison of ditches and spills .......................................................................... 21 4.9.4 Stream water quality ............................................................................................. 21 4.9.5 Macroinvertebrate analysis ................................................................................... 22 5 Results................................................................................................................................... 23 5.1 Chemical characterization of wetlands ......................................................................... 23 5.2 Wetland drainage experiment ....................................................................................... 25 5.3 Comparison of ditches and spills .................................................................................. 34 5.4 Stream water quality ..................................................................................................... 35 5.5 Description of macroinvertebrate assemblages ............................................................ 40 6 Discussion ............................................................................................................................. 43 6.1 Wetland water quality characterization ........................................................................ 43 6.2 Factors controlling temporal patterns in wetland water quality.................................... 46 6.3 Water quality characteristics of a newly constructed drainage ditch............................ 48 6.4 Comparing artificial ditches and natural spills ............................................................. 50 6.5 Stream water quality ..................................................................................................... 50 6.6 Exceedance of Federal and Provincial water quality guidelines for wetlands, ditches, spills and streams ...................................................................................................................... 51 6.7 Stream ecosystem health............................................................................................... 52 6.7.1 Effects of wetland drainage stress levels on receiving lotic ecosystem health..... 52 6.7.2 Findings on the influence of time on ecosystem health........................................ 53 6.7.3 Unanswered aspects of drainage-biotic relationships and future directions......... 53 7 Conclusions and recommendations....................................................................................... 54 7.1 Conclusions................................................................................................................... 54 7.2 Recommendations......................................................................................................... 55 8 Acknowledgements............................................................................................................... 56 9 References............................................................................................................................. 57 10 Other aspects..................................................................................................................... 65 10.1 Communications ........................................................................................................... 65 10.2 Personnel involved........................................................................................................ 65 11 Appendices........................................................................................................................ 66 2 List of Tables Table 4.1: Summary of number of wetlands sampled in each land cover and permanence class. 14 Table 4.2: Photographs of artificial ditches (DT) and natural connections (SP) studied that drain wetlands at Smith Creek watershed and means of physical properties measured along the connections: discharge (Q), velocity (v), depth (d), and width (w). Sample locations were measured from the wetland edge along the connection. ................................................... 16 Table 4.3: Wetland loss (%) and corresponding sub-basin drainage classification used in this study. The wetland loss data are derived from wetland areal extent changes between 1958 (Ducks Unlimited Canada data) and supervised land use classification data from 2007 and 2008 SPOT5 images (Fang et al., 2010). ................................................................... 17 Table 4.4: Sites, names and coordinates included in the Smith Creek ecosystem health assessment 2008 and 2009. ............................................................................................... 19 Table 5.1: Mean (standard error) of wetlands within each permanence and land cover class, as well as a summary of results for two-way ANOVAs. Semiperm is semipermanent wetlands. Differing letter superscripts indicate significantly different (α = 0.05) Tukey’s pairwise comparisons; * and ** denote a statistically significant difference at α = 0.05 and α = 0.01, respectively. ................................................................................................ 25 Table 5.2: Mean (standard error) stream export coefficients for the 2009 freshet by site. Differing superscripts indicate significant difference at α = 0.05, assessed using a one-way ANOVA and the Holm-Sidak method for multiple pair-wise comparisons..................... 39 Table 5.3: Sites sampled for ecosystem health in 2008. ............................................................... 41 Table 5.4: Metric values and their probability of difference from reference (Metric Significance) ........................................................................................................................................... 42 Table 5.5: Test Site Analysis results for sites from April 13, 2009 in the Smith Creek Watershed. D indicated the magnitude of effect or difference from reference, while ncP is the measure of significance..................................................................................................... 43 Table 5.6: Test Site Analysis results for sites from April 13, 2009 in the Smith Creek Watershed. D indicated the magnitude of effect or difference from reference, while ncP is the measure of significance..................................................................................................... 43 3 List of Figures Figure 2.1: Map of the prairie pothole region of North America (Euliss et al., 1999) ................... 8 Figure 2.2: Potential pathways of surface runoff. Runoff can a) enter streams directly; b) enter and be stored in wetlands; stored runoff can be released from wetlands via drainage ditches and flow either into c) other wetlands or d) streams (modified from McAllister et al., 2000). .......................................................................................................................... 10 Figure 3.1: Historic and current day wetland and stream channel distribution in Smith Creek watershed, produced in conjunction with Ducks Unlimited Canada................................ 13 Figure 4.1: Smith Creek watershed, Saskatchewan, Canada showing the location and ID numbers of wetlands studied in relation to soil map units............................................................... 14 Figure 4.2: Location of the ditches (DT) and spills (SP) sampled, and the extent of open water (includes wetlands and lakes) determined by Ducks Unlimited Canada from 2005 air photos (courtesy of Lyle Boychuk). Inset: Air photo of LR3 wetland, showing the sampling sites in the wetland and along the newly constructed drainage ditch (DR). The location of the water level recorder (PT2X) is indicated by a cross................................. 15 Figure 4.3: Location of stream sampling sites and the Environment Canada hydrometric gauge along Smith Creek. Sub-basins were identified by Fang et al. (2010). ............................ 17 Figure 4.4: Map of Smith Creek watershed with ecosystem health assessment sampling locations from: a) 2008, and b) 2009. Note that Smith Creek at Werle Farm is TV1 and Smith Creek East of Langenburg is SC3..................................................................................... 19 Figure 5.1: Distribution of cation and anion dominance groups as a function of specific conductivity for the 67 wetlands studied grouped by A) land cover and B) permanence classes. Semiperm is semipermanent wetlands................................................................. 24 Figure 5.2: Daily rainfall and volume in wetland LR3 prior to and during drainage. .................. 26 Figure 5.3: Concentration of nitrogen, phosphorus, and bacteria measured in the LR3 wetland prior to the drainage experiment and in the newly constructed ditch. .............................. 27 Figure 5.4: Total mass of nitrogen, phosphorus, and bacteria measured in the LR3 wetland prior to the drainage experiment and the cumulative mass exported via the newly constructed ditch................................................................................................................................... 28 Figure 5.5: Mass of various water quality parameters, normalized to May 1, 2008, measured in wetland LR3...................................................................................................................... 29 Figure 5.6: Mass ratio of DIN:DIP (NO3+NH4:PO4) measured in the LR3 wetland prior to its drainage. Values below seven indicate N limiting conditions and values above 12 indicate P limiting conditions (Wetzel, 2001). ............................................................................... 30 Figure 5.7: pH and concentration of DOC and major ions measured in the LR3 wetland prior to the drainage experiment and in the newly constructed ditch............................................ 31 Figure 5.8: Total mass of DOC and major ions measured in wetland LR3 prior to the drainage experiment and the cumulative mass exported via the newly constructed ditch. ............. 32 Figure 5.9: Slopes of normalized concentrations measured along the newly constructed ditch draining the LR3 wetland at: a) 1 hr, b) 4 hr, c) 6 hr, and d) 23 hr after the start of the drainage experiment. Concentrations were normalized by dividing the concentration at each sample point along the ditch by the concentration at the first sampling point (DR1) in the ditch. A value of one was then subtracted to set the intercept to zero. Only constituent slopes that were at least marginally different (α = 0.10) from the chloride slope and that had significant (α = 0.05) linear relationships between normalized concentration and distance are shown. P values indicate the level of significance for 4 slopes differing from chloride, suggesting that a portion of the variability is due to biotic processes or sorption......................................................................................................... 33 Figure 5.10: Mean and standard error of ditch and spill physical properties where * and ** denote statistically significant differences at α = 0.05 and 0.01, respectively.................. 34 Figure 5.11: Mean and standard error of A) nutrients and DOC, and B) major ions, SC, and pH in ditches and spills where * and ** denote statistically significant differences at α = 0.05 and 0.01, respectively. ...................................................................................................... 34 Figure 5.12: Mean and standard error of ditch and spill nutrient and DOC loads where *denotes a statistically significant difference at α = 0.05. .................................................................. 35 Figure 5.13: Discharge and water quality concentrations during the 2009 freshet at sites characterized by differing watershed drainage: high (TV1), medium high (SC4), and low (SC5). No data are presented prior to April 8, 2009 due to ice at the gauge station. ....... 36 Figure 5.14: Ratios of DIN (NO3 + NH4) to DIP (PO4) measured near the Smith Creek watershed outlet (SC3) during 2008 and 2009. Values below seven indicate N limiting conditions while those above 12 indicate P limiting conditions (Wetzel, 2001). .............................. 37 Figure 5.15: Piper plots for: basin outlet (SC3) and sub-basins characterized by high (TV1), medium high (SC4), and low (SC5) wetland drainage. Blue and red circles indicate samples obtained during the snowmelt period (March 23 – April 19, 2009) and following snowmelt (April 25 – May 26, 2009), respectively. ......................................................... 38 Figure 5.16: Total exports of: a) water (Q), nitrogen (TDN, NO3, and NH4) and phosphorus (TP and orthoP); b) major ions; and c) bacteria (E. coli and T. coli) during the 2009 freshet (3 April - 26 May) at sub-basins characterized by low, medium high, and high wetland drainage. Exports for the SC3 subbasin (c.f. Figure 4.3) were calculated by subtraction of exports for the three subbasins from that for the whole watershed. Total height of the bars indicates export for the whole Smith Creek watershed. ................................................... 40 5 1 Summary This report describes factors influencing the spatial variation in wetland water quality and how drainage of wetlands affects downstream receiving waters in terms of their water quality and biotic health. The specific objectives of this work were to: 1) characterize the spatial and temporal variation in water quality of prairie potholes after snowmelt; 2) quantify solute export along a newly constructed wetland drainage ditch; 3) characterize solute export from drained pothole wetlands; 4) determine the extent to which stream water quality is influenced by wetland drainage; 5) contribute to the understanding of how wetland drainage affects ecosystem health. The research was conducted at the Smith Creek watershed, southeastern Saskatchewan, where there has been controversy over recent renewed efforts to drain wetlands to increase agricultural production. A total of 67 wetlands were sampled following snowmelt in 2009 to determine whether spatial variations in wetland water quality could be attributed to different land cover and permanence classes. It was found that crop wetlands had greater TP and K than wood and grass wetlands; TP, TDN, and DOC were higher in seasonal than permanent wetlands; and salts were lower in wood compared to crop and grass wetlands. Measures of water quality of one permanent wetland over a 20 week period in 2008 showed that the wetland acted as a trap for nutrients, salts and bacteria. Variations in salts and DOC loads were linked to hydrological processes whereas variations in nitrogen, phosphorus, and bacteria loads were associated with both biotic and hydrological processes. The permanent wetland studied was experimentally drained in November 2008. The experiment demonstrated that wetland water quality was an important control of water quality in drainage water. The wetland ditch acted as a simple conduit, meaning that there was little loss or gain of solutes along the length of the ditch. Results also show that the efficiency with which a wetland is drained and the water quality characteristics of the wetland are the factors critical to determining solute exports via ditches. In spring 2009, water quality along seven ditches and five natural connections that form between wetlands (termed spills) was found to be similar, except for concentrations of TDN, DOC, HCO3, K, and Ca that were higher in ditches than spills. As in the wetland drainage experiment, little change in the water quality along ditches and spills was found, likely due to the low temperatures occurring in spring that can restrict biotic processing. The important difference however was the physical characteristics of ditches and spills. The long ditches connect wetlands to streams whereas short spills connect wetlands to other wetlands. Thus ditches have a greater potential to contribute to downstream loading of nutrients, salts and bacteria. Water quality of streams draining three subbasins of the Smith Creek watershed with varying wetland drainage intensities was compared during the 2009 freshet. Nearly all export coefficients for the water quality parameters studied were higher in streams draining subbasins with greater wetland drainage. Total solute export was greatest in the subbasin with medium high wetland drainage, due in part to its comparatively large size. Although water samples generally did not exceed guidelines for nitrogen and salts, the provincial objective for TP was frequently exceeded. Consequently, wetland drainage is likely to exacerbate downstream eutrophication. The macroinvertebrate-based ecosystem health of Smith Creek watershed in stream reaches downstream of drainage activity was assessed and compared to ecosystem health at similar sites across southern Saskatchewan. Sampling was conducted through 2008 and 2009 at four and eight sites in each of these respective years. It was found that wetland drainage does not 6 significantly impact the ecosystem health of receiving waters, and may in fact benefit the macroinvertebrate assemblage, at least in the short term, by increasing the amount of water and habitat available. However, this project did not address the potential decline in ecosystem health in source-wetlands that are being drained, where the amount of habitat and water is decreasing. Overall, this study represents the first field-based research in the Prairie Pothole Region on the effects of wetland drainage on downstream water quality and stream ecological health. Therefore, results should help support effective management decisions regarding future pothole drainage or restoration, balancing public and private costs and benefits. Several specific recommendations were made regarding the challenges faced by this study. It is recommended that similar studies be conducted in other geographic areas of the prairies where climate, soils, wetland configuration and drainage may produce differing results. 7 2 Introduction Prairie pothole wetlands are small, shallow depressions that typically lack surface water connections and are ubiquitous across the prairie pothole region (PPR) of North America (Figure 2.1). The PPR is estimated to cover approximately 715 000 km2 (Euliss et al., 1999). Pothole wetlands formed during the last glacial retreat that created the hummocky, undulating terrain typical of the prairies (Tiner, 2003). About 40%, or 156 000 km2, of the PPR located in Canada consists of hummocky moraines that have a wetland density of 18 wetlands/km2. The remaining 60% of the Canadian PPR landscape is comprised of mostly lacustrine and fluvial materials that have, on average, 5 wetlands/km2 (National Wetlands Working Group, 1988). Pothole wetlands provide important hydrological and ecological functions. For example, the PPR represents only 10% of the continent’s waterfowl breeding area but it produces half of North America’s waterfowl in an average year (Smith et al., 1964; Batt et al., 1989). Figure 2.1: Map of the prairie pothole region of North America (Euliss et al., 1999) Many of the wetlands located in the hummocky moraine region are isolated prairie potholes. These potholes range in permanence from those that contain water for only a few days following spring snowmelt, to those that are continuously inundated. Most potholes do not normally contribute to streamflow (Stichling and Blackwell, 1957). During very wet conditions temporary surface connections can occur among them (Leibowitz and Vining, 2003; Winter and LaBaugh, 2003), which is referred to as the ‘fill and spill’ mechanism (Spence and Woo, 2003; Spence, 2006). Water quality is defined differently by engineers, ecologists, hydrologists, etc., and can encompass a wide range of water quality descriptors depending on the end user, the context of interest, and natural conditions (Meybeck, 2005). Water quality, as defined in this report, is based on chemical, physical, and biological descriptors that affect the structure and function of ecosystems as well as those that negatively impact human and livestock health if in elevated concentrations. Isolated prairie potholes exhibit high spatial and seasonal variations in water quality (e.g., LaBaugh and Swanson, 2004). Previous studies have documented variations in select water quality variables in relation to individual factors such as wetland permanence, water sources, and land use of the catchment area (Rözkowska and Rözkowski, 1969; Miller et al., 1985; LaBaugh et al., 1987; Swanson et al., 1988; Neely and Baker, 1989; Detenbeck et al., 2002; Waiser, 2006). For example, Driver and Peden (1977) studied prairie pond chemistry in Saskatchewan and Manitoba and found differences among temporary, semi-permanent, and permanent ponds. Crosbie and Chow-Fraser (1999) found higher nutrient concentrations in Ontario marshes that a greater proportion of agricultural land in their watersheds. However, a 8 concurrent assessment of nutrients and salts in potholes is needed given that different mechanisms interact to drive their concentrations (LaBaugh et al., 1987; Wetzel, 2001). As well, factors such as permanence and land cover type do not act in isolation on the landscape and could instead interact with one another to regulate wetland water quality. Studies examining the potential governing influence of the interaction of permanence and land cover type on pothole water quality are lacking. Approximately 40-70% of the potholes in the western prairies have been drained since 1900 to increase agricultural production (Tiner, 1984; Dahl, 1990; Brinson and Malvarez, 2002). Recently, there have been renewed efforts to drain potholes (Watmough and Schmoll, 2007). Temporary wetlands are the most common permanence class in the PPR, and the most impacted by drainage and farming practices (Euliss et al., 2001). Issues surrounding pothole drainage centre around attempts to balance the social benefits and private costs associated with potholes on agricultural lands (Porter and van Kooten, 1993). The social benefits of potholes are believed to be water storage and flood attenuation, wildlife habitat, and improvements of water quality (Saskatchewan Watershed Authority, 2007a). Costs accrued by private landowners include the nuisance of farming around potholes, delayed seeding dates, and the foregone opportunity to increase agricultural production (Scarth, 1998; Brinson and Malvarez, 2002; Saskatchewan Watershed Authority, 2007a). Drainage impacts remain a critical information gap for the agricultural community, with the potential to have broad-scale economic impacts. Drainage ditches create new surface water connections between wetlands that were previously isolated and other wetlands, roadside ditches, and streams. The new connections transform the hydrologic conditions of the prairies such that previously non-contributing areas now contribute to streamflow. Figure 2.2 illustrates the different ways that surface water can enter a stream where drainage ditches are present (McAllister et al., 2000). Isolated wetlands have the potential to intercept and store surface runoff (Figure 2.2b). Drainage ditches can transport water from one wetland to another (Figure 2.2c), which can cause local flood damage to cropland or communities surrounding the terminal wetland. Drainage ditches can also transport surface water runoff directly from a wetland to a stream (Figure 2.2d). Drainage of many potholes has the potential to significantly increase downstream flood frequencies and magnitudes (Campbell and Johnson, 1975). For example, Yang et al. (2008) used the SWAT model to show that a loss of 70% of 1968 wetlands in the Broughton’s Creek watershed in western Manitoba to drainage and degradation increased the basin’s contributing area by 31% (19 km2), increased peak flows by 18%, and increased stream flow by 30%. Pomeroy et al. (2009) used the newly developed Prairie Hydrological Model to show a 117% increase in streamflow volumes with complete drainage of the wetlands in Smith Creek watershed, Saskatchewan based on the 2007-2008 meteorological conditions. Further, a Saskatchewan Watershed Authority (2008) assessment identified agricultural drainage as an important contributor to high water levels in terminal (Waldsea) and near-terminal (Deadmoose, Houghton, Fishing) lake basins in 2007. Pothole wetlands have been shown to trap nutrients, salts and bacteria from catchment runoff (Neely and Baker, 1989; Crumpton and Goldsborough, 1998). Wetland water quality is thus expected to be an important control of water quality in drainage water. As wetland drainage connects potholes to downstream water bodies, it has the potential to generate excessive nutrients and sediment pollution that may impact the water quality, and ultimately the ecosystem health of receiving streams and rivers (Leibowitz and Vining, 2003; Whigham and Jordan, 2003). To date, however, there have not been field studies to support or refute this conjecture. Despite a lack of 9 data, the popularly held belief that wetland drainage will impair downstream waters has influenced decision making in the courts. For example, a recent decision by the Water Appeal Board (16 August 2007) in the case of Ducks Unlimited Canada vs. Jack Kalmakoff to close a drainage ditch was due, in part, to a perceived risk of impact to downstream aquatic systems resulting from wetland drainage. While no studies of water quality in wetland drainage ditches has been conducted in the PPR, there is a body of literature describing nutrient export along upland drainage ditches located in low order agricultural watersheds, which may prove useful to understanding solute export from potholes drainage. Main findings are reviewed in the following two paragraphs. Figure 2.2: Potential pathways of surface runoff. Runoff can a) enter streams directly; b) enter and be stored in wetlands; stored runoff can be released from wetlands via drainage ditches and flow either into c) other wetlands or d) streams (modified from McAllister et al., 2000). The water regime in upland drains is characterized by the advective flow of water downstream. Turbulent mixing encourages oxygenation of the water. For redox-sensitive water quality parameters, increased oxygenation during transport along ditches would be expected to change their concentrations. Kemp and Dodds (2001) showed stimulation of nitrification and the inhibition of denitrification in a 2nd order prairie stream with higher dissolved oxygen (DO) concentrations, which would likely result in a net reduction in N removal along the stream (Birgand et al., 2007). Stimulated nitrification rates can also lead to increased transport of nitrogen (N) to receiving streams since nitrate (NO3-) is quite mobile compared to ammonium (NH4+), which is easily adsorbed to negatively charged ditch surface particles (Strock et al., 2007). Solute sedimentation rates vary inversely with the velocity of flowing water (Julien, 2002), affecting concentrations of water quality parameters that have an affinity for the particulate phase, for example, phosphorus (P). Recent studies have shown relatively high retention of mostly P (as well as some N) in upland ditches where soils have substantial sorption or retention capacities (Sharpley et al., 2007; Strock et al., 2007). Nguyen and Sukias (2002) showed ditch sediments in New Zealand contained 42-57% of P originating from agricultural catchments loosely bound with Al, Fe, and CO3-, and 6-39% of P stored more permanently in the sediment as refractory P. They also showed the proportion of P transported was governed by the form of P and the retention characteristics of the ditch sediments. Periodic high flow events that occur during snowmelt and significant rainfall events increase velocity, shear force, and scour along the ditch bottom causing the re-suspension of sediments and organic matter and consequently their downstream 10 transport (Sharpley et al., 2007; Birgand et al., 2007). Also, macrophytes and algae living in the drains have been shown to temporarily store nutrients (Needleman et al., 2007). When they die during season changes, water level drops, or other causes, they can contribute to the organic matter content of the water column and also to the accretion of drain sediment. Macrophytes can also play an indirect role in contaminant retention by reducing flow velocities and re-suspension rates, and increasing sedimentation rates (Birgand et al., 2007). There is very little published research on the relationship between land use and wetland drainage on stream water quality in the Canadian Prairies. Carpenter et al. (1998) is the seminal paper outlining the issue of how stream water quality is controlled by land use. Alberta Agriculture and Rural Development prepares annual reports cards for stream water quality, as measured through a water quality index, in Alberta’s watersheds with varying agricultural intensities. Saskatchewan Watershed Authority makes similar reports for watersheds in Saskatchewan using a different water quality index. Stream biotic integrity is commonly assessed through measures of the aquatic macroinvertbrate community. Summaries of the ecosystem structure and function in macroinvertebrate communities are used as proxies of ecosystem health, and are frequently applied to describe impacts of human activities. In particular, specific components of the macroinvertebrate assemblage are sensitive to abiotic changes in the aquatic environment, and the expression of these changes as metrics relative to reference, or unaffected streams, is to evaluate impact. By identifying metrics that respond to specific stressor, management can be applied to the stressors in a watershed that are causing ecosystem degradation. Leibowitz and Vining (2003) suggest that stream water quality is likely to be altered by wetland drainage as this would hydrologically connect previously isolated wetlands to streams. Similarly, how Saskatchewan macroinvertebrate assemblages respond to stressors brought about through wetland drainage (e.g., nutrients, sediment etc.) is poorly understood. Tangen et al. (2003), in their attempt to create a macroinvertebrate index of biotic integrity found only a weak correspondence of macroinvertebrate assemblages and land use in the Prairie Pothole region of North Dakota due to an inadequate number of low impact sites. Wetland drainage was not one of the land use factors considered. Further, as benthic macroinvertebrate assemblages are affected by a number of factors in addition to wetland drainage, changes in assemblage composition should currently be interpreted as a complimentary tool to primary water quality tests. The overall objective of this research project was to determine changes in water quality of streams and impacts to stream ecosystem function associated with wetland drainage. The specific research objectives of this study were to: 1) characterize spatial and temporal variations in water quality of prairie potholes after snowmelt; 2) quantify solute export along a newly constructed wetland drainage ditch; 3) compare solute export from naturally and artificially drained pothole wetlands; 4) determine how stream water quality is influenced by wetland drainage; and 5) contribute to the understanding of how wetland drainage affects ecosystem health by quantifying benthic macroinvertebrate communities. Saskatchewan Watershed Authority was sub-contracted to complete objective 5. 11 3 Study Site The research was conducted at Smith Creek watershed (SCW; 50°50'4"N 101°34'48) in southeastern Saskatchewan. The watershed is ~400 km2 (Pomeroy et al., 2009) with an effective contributing area of ~58 km2 (Environment Canada, 2006). The contributing area is continuously increasing as producers drain more potholes to increase arable area. Smith Creek drains into the Assiniboine River and then Lake Winnipeg where excessive N and P loadings are causing eutrophication (Armstrong, 2002). Snowmelt is the dominant, and often only, hydrological event. The terrain of SCW is level to undulating and rolling. SCW is located in the Aspen Parkland Continental Prairie Wetland sub-region (National Wetlands Working Group, 1988). Soils in the watershed are a mixture of Black (Oxbow) and Thick Black (Yorkton) chernozems formed in loamy glacial till (Agriculture and Agri-Food Canada, 2009). The regional climate is semi-arid. Average seasonal temperatures are -17.9oC and 17.8oC for the winter and summer, respectively (as measured at the Yorkton airport, ~50 km west). The mean (1942-2009) annual precipitation is 438 mm with 28% falling as snow. Precipitation amounts prior to the 2008 study period were in the 53rd, 34th, and 26th percentiles for winter 2007, May – October 2007, and winter 2008, respectively. Summer 2008 and winter 2009 precipitation amounts were in the 60th and 66th percentiles, respectively. Total precipitation for the months of May 2008 (18 mm) and July 2008 (208 mm) were respectively in the 13th and 95th percentile of values measured at the Yorkton weather station. Land uses were determined by Pomeroy et al. (2009) using unsupervised classification of SPOT 5 images from October 1, 2008. The dominant land use is agriculture, which occupies 54% of the watershed. Common crops include wheat, canola, and flax. Eight percent of the watershed is grassland and pasture. Wooded areas and wetlands/open water account for 23% and 11% of the watershed, respectively. Wooded stands are characterized by trembling aspen (Populus tremuloides) with pockets of balsam poplar (Populus balsamifera), together with an understory of mixed herbs and tall shrubs. Wetland vegetation is predominately willow (Salix spp.), cattails (Typha latifolia L.), sedges (Carex spp. and Scirpus spp.), duckweed (Lemna spp.), pondweed (Potamogeton spp.), and water smartweed (Polygonum amphibium L.). Grasslands are comprised largely of western porcupine grass (Stipa curtiseta), plains rough fescue (Festuca hallii), pasture sage (Artemisia frigida), and Lewis wild flax (Linum lewisii). The majority of the wetlands in SCW belong to the marsh and shallow open water classes (National Wetlands Working Group, 1988). The average wetland density in the basin is ~20 wetlands/km2. Many of the wetlands in the basin are typical, isolated prairie potholes that formed in glacial depressions, which at average surface water level have no surface inflows or outflows. The wetlands range from temporary through permanent. Recently, a number of drainage ditches have been constructed to enhance agricultural production. These drainage ditches either completely eliminate the wetland or drastically lower the water levels and create downstream connections with other wetlands, roadside ditches, or streams. Between 1958 and 2001 the portion of land cover occupied by wetlands in the Smith Creek watershed decreased ~60%%. Drainage has been most prolific in the eastern and southwestern areas of the watershed (Figure 3.1). Many water control structures such as road culvert gates exist in the basin and are operated by local farmers to regulate the runoff in their cropland areas; the gates are closed during extremely high runoff periods, i.e. during fast snowmelts or intense rain storms but remain open otherwise. 12 2009 1958 Figure 3.1: Historic and current day wetland and stream channel distribution in Smith Creek watershed, produced in conjunction with Ducks Unlimited Canada. 4 Methods 4.1 Wetland water quality characterization Water quality in 67 wetlands (Figure 4.1) was assessed following snowmelt in 2009. Wetland selections were made based on obtaining relatively equal numbers of wetlands in the different land cover and permanence classes (Table 4.1), as well as on accessibility. Land cover classes were crop, wood, and grassland while permanence classes were seasonal, semipermanent, and permanent. Wetland permanence classes were determined using a combination of the vegetation structure of the pothole, as per Millar (1976) and Stewart and Kantrud (1971), assessed in May and mid-August 2009, in combination with observations of the presence or absence of surface water recorded on air photos (26 October 1959 and ~31 May 2001), and SPOT 5 imagery (5 July 2007). Water samples (1 L) were collected once during 19-21 May 2009 from roughly the deepest point in the wetland, which is typically the centre of the ponds, at the midpoint in the water column. 13 Figure 4.1: Smith Creek watershed, Saskatchewan, Canada showing the location and ID numbers of wetlands studied in relation to soil map units. Table 4.1: Summary of number of wetlands sampled in each land cover and permanence class. Land Cover Type Crop Grass Wood Total Seasonal 7 9 6 22 Permanence Class Semipermanent 6 7 7 20 Permanent 8 8 9 25 Total 21 24 22 67 4.2 Design of wetland drainage experiment A drainage experiment was carried out on a permanent wetland, LR3. It was selected for study because hydrological data for it prior to drainage were available from a concurrent study (c.f. Minke et al., 2010). Additionally, the landowner was keen to drain it and the drain was expected to connect to Smith Creek. Water samples were collected weekly at the center of the wetland LR3 from 18 April to 22 October 2008. Water level was measured hourly using a PT2X pressure transducer (Northwest Instrumentation Inc.) which was located near the edge of the wetland. Rainfall was measured nearby using a tipping bucket (Texas Electronics Inc., TR-525M) and a standard volumetric rain gauge. A crawler excavator and professional operator were employed to construct the artificial drainage ditch (DT6) starting 19 November 2008. Water sample collection 14 began 20 November 2008 (1 hr) once the ditch was completed and connected to a downstream wetland. Three additional sets of samples were collected 4 hrs, 6 hrs, and 23 hrs after the start of drainage. Water samples were collected from within the wetland and at points 45 m, 70 m, 110 m, and 140 m along the ditch, measured from the wetland edge (Figure 4.2 inset) at the midpoint in the water column. Figure 4.2: Location of the ditches (DT) and spills (SP) sampled, and the extent of open water (includes wetlands and lakes) determined by Ducks Unlimited Canada from 2005 air photos (courtesy of Lyle Boychuk). Inset: Air photo of LR3 wetland, showing the sampling sites in the wetland and along the newly constructed drainage ditch (DR). The location of the water level recorder (PT2X) is indicated by a cross. 4.3 Natural spills and artificial ditches sampling strategy Seven artificial drainage ditches (hereafter, ditches, DT) and five natural connections (hereafter, spills, SP) (Table 4.2) were selected for study. The ditches and spills drained wetlands that did not have any surface water inflows, with the exception of DT3, which had receives inflow from DT1. However, at the time of sampling, DT1 was not flowing because it was frozen and snow covered. Due to the variation in ditch and spill length, locations for water sampling along the ditch varied; they are reported in Table 3.2. Water samples were collected 10-18 April 2009 from the thalweg of the connection, starting at the most downstream sample location. Samples were obtained from the midpoint of the water column. Manual flow gauging was carried out at the time of water quality sampling using a Marsh-McBirney Flo-mate 2000 velocity meter and wading rod. Velocity at 60% depth was measured at 20 – 40 cm intervals, so that no more than 20% of the total stream discharge was measured at each point. Ditch or spill discharge was then calculated as the sum of the product of velocity and average stream depth at each sample interval. 15 Table 4.2: Photographs of artificial ditches (DT) and natural connections (SP) studied that drain wetlands at Smith Creek watershed and means of physical properties measured along the connections: discharge (Q), velocity (v), depth (d), and width (w). Sample locations were measured from the wetland edge along the connection. Sample locations (m): Sample locations (m): 0, 30, 60, 90, 120, 150 0, 25, 50, 75, 100 Q: 0.06 cms Q: 0.002 cms v: 0.04 m/s v: 0.009 m/s d: 0.37 m d: 0.07 m w: 3.05 m w: 2.99 m Sample locations (m): Sample locations (m): 0, 25, 50, 75, 100 0, 25, 50, 75, 100, 125 Q: 0.02 cms Q: 0.16 cms v: 0.02 m/s v: 0.11 m/s d: 0.33 cm d: 0.27 m w: 4.45 m w: 5.59 m Sample locations (m): Sample locations (m): 0, 25, 50, 75, 125, 175 0, 25, 50, 75, 100, 125 Q: 0.02 cms Q: 0.06 cms v: 0.01m/s v: 0.06 m/s d: 0.22 m d: 0.32 m w: 11.0 m w: 4.01 m Sample locations (m): Sample locations (m): 0, 50, 100, 150, 200, 250 0, 5, 10 Q: 0.02 cms Q: 0.07 cms v: 0.05 m/s v: 0.01 m/s d: 0.16 m d: 0.28 m W: 3.05 m w: 22.3 m Sample locations (m): Sample locations (m): 0, 15, 30, 45 0, 8, 15 Q: 0.02 cms Q: 0.006 cms v: 0.005 m/s v: 0.02 m/s d: 0.16 m d: 0.07 m W: 27.1 m w: 6.40 m Sample locations (m): Sample locations (m): 0, 9, 18, 36 0, 10, 20 Q: 0.002 cms Q: 0.001 cms v: 0.001 m/s v: 0.001 m/s d: 0.18 m d: 0.10 m W: 8.49 m w: 4.57 m 4.4 Stream water quality sampling Stream water samples were collected at three locations along Smith Creek in 2008, and four locations in 2009. One sample site was the watershed outlet (SC3), and the other sample sites were three sub-basins of Smith Creek (SC4, SC5, TV1; Figure 4.3) that represent areas of the watershed with differing degrees of wetland drainage. Wetland distribution across the watershed was historically (1958) even (Fang et al., 2010). Although sampling sites were chosen to reflect differing proportions of wetland loss between 1958 and 2007-2008, quantification of wetland loss was completed after site selection through the land cover classification of Fang et al. (2010). Wetland losses in the three subbasins sampled and resulting drainage classes used in this study are reported in Table 4.3. Stream water quality measurements were taken daily during peak flow (spring runoff), weekly during low flows, and following heavy summer rains (2008 16 only). Stream discharge at the subbasin sampling sites was estimated at the time of water quality sampling (2009 only) using a Marsh McBirney flow meter. Streamflow at the watershed outlet (Figure 4.3) was obtained from Water Survey of Canada gauge 05ME007. The water quality sampling station representing the watershed outlet was located just upstream of the town of Langenburg’s drainage (SC3; Figure 4.3) as preliminary chemical analyses showed that it contributes high fecal bacteria and nutrients to Smith Creek. Figure 4.3: Location of stream sampling sites and the Environment Canada hydrometric gauge along Smith Creek. Sub-basins were identified by Fang et al. (2010). Table 4.3: Wetland loss (%) and corresponding sub-basin drainage classification used in this study. The wetland loss data are derived from wetland areal extent changes between 1958 (Ducks Unlimited Canada data) and supervised land use classification data from 2007 and 2008 SPOT5 images (Fang et al., 2010). Sample Site ID SC5 SC4 TV1 Wetland Loss 1958-2007/8 in Sub-Basin (%) 23 57 64 Drainage Class Low Medium High High 4.5 Analysis of water quality Water samples for bacteria, i.e. Escherichia coli (E. coli) and total coliforms (T. coli), were only collected for the wetland drainage experiment and from the stream due to challenges associated with transport times to the laboratory. They were collected in 100 mL sterile bottles, preserved with Na2S2O3, and submitted to Saskatchewan Research Council Analytical Laboratories, Saskatoon, SK (SRC) within 24 hours. They were analyzed using the chromogenic substrate method (Standard Methods part 9223). Water samples for ion and nutrient analyses were collected in pre-rinsed 1 L polyethylene or two 350 mL glass bottles, kept on ice during the day, and then split into aliquots. 17 Temperature-compensated specific conductance (SC) and pH were measured in the laboratory on the day of sampling using Hach sension156 and Orion 3-Star hand-held meters, respectively. Two sample aliquots were preserved by the addition of HNO3 and H2SO4, respectively, followed by refrigeration at 4oC. A third aliquot was filtered through a 40 μm Whatman GF/C glass microfiber filter then frozen. The HNO3 preserved sub-sample was analyzed for total phosphorus (TP) concentration at SRC within three days of sampling by inductively coupled plasma atomic emission spectroscopy (Standard Methods part 3120). Samples preserved with H2SO4 were only collected for the 2008 wetland drainage experiment and analyzed for total Kjeldahl nitrogen (TKN) at SRC within three days of sampling by digestion and subsequent ammonia analysis (EPA 351). Filtered samples were analyzed for a variety of chemical parameters. Total dissolved nitrogen (TDN) and dissolved organic carbon (DOC) for samples collected in 2009 were analyzed at the University of Saskatchewan on a Shimadzu TNM-1. DOC samples from 2008 were analyzed at SRC by UV persulfate digestion and non-dispersive IR detection, (Standard Methods part 5310C). Ortho-phosphate (orthoP) was also analyzed at SRC colorimetrically (Standard Methods part 4500). A Westco SmartChem Discrete Analyzer (SmartChem 200, Method 375-100E-1) was used for analysis of ammonium (NH4) and nitrate plus nitrite (reported as NO3). Major ions (Cl, HCO3, SO4, Na, K, Mg, and Ca) were analyzed by ion chromatography with a Dionex Model ICS-2000 using potassium hydroxide and methanesulfonic acid EluGen for anion and cation analysis, respectively, at the University of Saskatchewan. Carbonate concentrations were assumed to be negligible due to high sample pH and its absence in pilot tests. One duplicate and one blank sample are analyzed for every fifteen samples collected for QA/QC. Note that charges have been left off ions throughout this report to enhance readability. 4.6 Ecosystem health sampling sites and frequency Benthic macroinvertebrate-based biomonitoring approaches are best applied in two time windows through the course of a year – early spring shortly after ice-off, and fall shortly before ice-on (Hilsenhoff 1988). Because many Smith Creek streams cease to flow by midsummer, benthic macroinvertebrates were sampled in late April and early May in 2008 and 2009. Our 2008 study of the Smith Creek provided sampling of four sites on May 8, 2008 (Figure 4.4a; Table 4.4). However, in 2009 we sampled the stream sites on two occasions early in the spring; once on April 16, 2009 (8 sites) as the freshet commenced, and later on May 13, 2009 as water levels declined (Figure 4.4b; Table 4.4). Our first sampling event in 2009 involved collecting macroinvertebrates from eight sites comprised of four Ditch sites (DT 1, DT 2, DT 3, and DT 4) and four stream sites (SC 3, SC 4, SC 5, and TV 1). Ditch site DT 5 included in the water quality and hydrology components of this project was not included here as it was too ephemeral and observations showed that it lacked macroinvertebrates altogether. In the second sampling for 2009, however, we only repeated sampling at the four stream sites. 18 Table 4.4: Sites, names and coordinates included in the Smith Creek ecosystem health assessment 2008 and 2009. Stream name Location SWA_Code Smith Creek Smith Creek Langenburg Creek Smith Creek Smith Creek Tributary Smith Creek Tributary Smith Creek Tributary Smith Creek Tributary Smith Creek Smith Creek Thingvala Creek Smith Creek Smith Creek North of Marchwell Smith Creek East of Langenburg Langenburg Creek East of Langenburg Smith Creek at Werle Farm Ditch at DT3 Ditch at DT1 Ditch at DT4 Ditch at DT2 Smith Creek at SC4 Smith Creek at SC5 Thingvala Creek at TV1 Smith Creek at SC3 SWA_2008_16 SWA_2008_17 SWA_2008_18 SWA_2008_19 SWA_2009_9 SWA_2009_10 SWA_2009_11 SWA_2009_8 SWA_2009_42 SWA_2009_43 SWA_2009_6 SWA_2009_7 UTM13East UTM13North 740829.6 736372.3 734626.6 732536.2 723906.0 723876.0 723870.0 725264.0 726911.7 727319.9 731532.3 736414.4 5636736.0 5638305.1 5637867.9 5640900.0 5645464.0 5645266.0 5645266.0 5652321.0 5648988.6 5648240.1 5641002.9 5638022.5 Figure 4.4: Map of Smith Creek watershed with ecosystem health assessment sampling locations from: a) 2008, and b) 2009. Note that Smith Creek at Werle Farm is TV1 and Smith Creek East of Langenburg is SC3. 4.7 Macroinvertebrate assemblage sampling methods At each site, four qualitative 500 μm mesh D-frame net samples were collected along the length of the reach (defined as 6 times bankful width). Each sample consisted of five, 10 second, ~ 0.3 m2, composite sweeps across the width of the stream; at the left bank, ¼ distance across, the middle, ¾ distance across, and the right bank. Samples were concentrated on a 500 μm mesh sieve and preserved in 80 % ethanol in the field. Organisms were returned to the laboratory where they were sorted from the organic material under 7 X magnification and stored in 80 % ethanol until they were identified to lowest possible taxon designation. Samples were subsampled where necessary. Final macroinvertebrate abundance was calculated from the 19 subsample fraction and extrapolated to that of the original sample. Samples were required to produce a minimum of 100 specimens to be included in the data analysis. Benthic macroinvertebrates were identified to lowest possible designation using keys for North America (Merritt and Cummins, 1996) and Western Canada (Brooks and Kelton, 1967; Dosdall and Lemkuhl, 1979; Clifford, 1991; Larson et al., 2000; and Webb, 2002). Voucher series were deposited in both the Saskatchewan Watershed Authority Invertebrate Voucher Collection (Saskatoon, Saskatchewan), and the Royal Saskatchewan Museum (Regina, Saskatchewan). 4.8 Habitat Assessment Physical characteristics of habitat structure were collected at each site in addition to benthic macroinvertebrate sampling. Recorded were in-stream substrate composition, vegetation, algae, and detritus compositions along with notes on surrounding land use and field water quality measures at the time of sampling. These data and other more detailed GIS information are used in the test site analysis and reference condition assessment in the future. The characteristics described in the habitat assessments are summaries and coarse descriptors of habitat condition at sites, and follow the rapid assessment protocols used by other jurisdictions in the United States (Barbour et al., 1999). Three main areas were covered in the site evaluation were substrate (% composition of material, dominant sub-dominant classification), in-stream habitat (macrophyte, algae, woody debris, and detritus use and prominence as well as characteristics of embeddedness and riparian cover), and human land use (riparian land use classification). This information will be used in discerning the condition of the sample sites when evaluating them in test site analysis in the future. 4.9 Data analyses 4.9.1 Characterization of wetland water quality A multivariate analysis of water quality was carried out to evaluate the influence of wetland permanence and surrounding land cover type on resulting wetland water quality. Data were log transformed to correct for nonnormality and heteroscedasticity (Legendre and Legendre, 1998). Following preliminary data analysis, the water quality data set was divided into nutrient (TP, orthoP, TDN, NO3, NH4, DOC, and K) and salinity (SC, pH, Cl, HCO3, SO4, Na, Mg, K, and Ca) variable sets because the mechanisms controlling nutrient concentrations in the wetlands have been shown to be different from mechanisms controlling ionic concentrations (Labaugh, 1987). Mass per volume concentrations (i.e., mg/L) were converted to equivalence concentrations (i.e., meq/L) for direct comparison of major ions. A two-way multivariate analysis of variance (MANOVA) was used to test the relationships among land cover and permanence for the water quality parameters measured. Post hoc comparison tests (two-way ANOVAs) were used to determine which variables contributed to the occurrences of significant differences between factors. Significant differences among land cover and permanence classes were assessed using Tukey's Honest Significant Difference (HSD) pairwise comparisons test, which is applicable to mildly unbalanced designs (Everitt and Hothorn, 2006). Statistical analyses were conducted using the R statistical language and environment (R Core Development Team, 2005). A type I error rate of 0.05 was used in significance tests unless otherwise stated. 20 4.9.2 Wetland drainage experiment Wetland volumes were estimated using the full volume-area-depth method (Hayashi and van der Kamp, 2000) in combination with PT2X water level data. Coefficients required for the estimates were obtained by Minke et al. (2010) from a digital elevation model derived from total station survey data. The volume of ice water in the wetland at the time of drainage was estimated using the density of ice (920 kg/m3) and the average ice thickness (Andres and Van Der Vinne, 2001). Constituent mass in the wetland was estimated using the concentration and the wetland volume estimate at the time of sampling. Changes in solute concentrations relative to chloride, which is impacted by evapotranspiration and dilution in the wetland but is biotically conservative, were used to indicate biotic or geochemical processing of constituents in the wetland (Duff et al., 2009). The average concentration along the ditch multiplied by the change in estimated wetland volume at each sampling point was used to calculate total loads exported from the wetland via the newly constructed drainage ditch. Loads were normalized along the length of the new ditch by dividing the load at each sample point along the ditch by the load at the first sampling point (DR1) in the ditch. A value of one was then subtracted to set the slope intercept to zero. The statistical significance of the linear relationship with normalized load and distance along ditch was tested using the linear model (l m) function of the R statistical language and environment (R Core Development Team, 2005). The slopes of the normalized concentrations were statistically compared as per Zar (2008) to the slope of biotically conservative Cl, at each sampling instance to assess whether constituents were added or removed along the length of the ditch. Significantly different slopes indicate nutrients or salts are either abiotically or biotically removed (or added) as water travels along the ditch length. 4.9.3 Comparison of ditches and spills Statistically significant differences in water quality constituents (concentrations and loads) and physical properties between ditches and spills were assessed with t-tests computed using SPSS (version 14.0). Data used were log transformed, means measured along each ditch or spill. Differences between loads at the connection inlet and outlet were also compared using ttests to assess whether transformations of water quality constituents occurred along the length of the connection. A type I error rate of 0.05 was used unless otherwise stated. 4.9.4 Stream water quality To account for differences in sub-basin size and water flows, export coefficients (Eij) for nutrients and salts were calculated by Eij = Cij Qij (1) Ai where C and Q are concentration and discharge measured on day j, and A is the area of subbasin i. This permitted direct statistical comparison of findings among subbasins characterized with differing wetland drainage. Eij values were compared during the 2009 snowmelt period among subbasins using a one-way ANOVA and Sidak post-hoc comparisons in SPSS v.14.0. Additionally, total water and solute exports for spring melt in 2009 were computed by summing daily values. Solute data from 2008 are not reported because of lack of discharge measurements 21 at all but site SC3 due to equipment failure. However, trends in water quality concentrations in 2008 were similar to those reported for 2009. 4.9.5 Macroinvertebrate analysis For a complete description of the construction of SWA’s RCA and Test Site Analysis (TSA) based aquatic ecosystem health tool please refer to the Aquatic Health Feasibility study (Phillips and McMaster, 2010). A summary of tool construction is provided below. Reference sites included in this study are a subset of 100 sites collected in 2006 and 2007 as part of the construction of SWA’s aquatic health feasibility study (Phillips and McMaster, 2010). Reference sites were separated from a larger set of sites based on the amount of human land-use in a buffer area 10 km long and 1 km wide on either side of the stream, upstream of the site. Because of the predominance of agricultural development in southern Saskatchewan watersheds, reference condition is not identified as pristine land-cover with the absence of human activity, but more practically at that condition available that has the ‘least’ amount of human activity (Stoddard et al., 2006). Specifically, reference site candidacy was restricted to the following criteria: < 50% cropland, < 5% urban land-use, < 50% pasture, < 80% total land under human influence (combination of cropland, urban, and pasture), ≤ 2 landfills, oil wells, bridges or road crossings upstream, and no reservoirs within 10 km. Groups of reference sites with similar community compositions were identified using cluster analysis, and used to define what underlying physiochemical conditions structure macroinvertebrate communities in southern Saskatchewan. In these analyses, it was found that surficial geology composition in the 10 km buffer upstream of the site, and the site ecoregion membership best explained the composition of the macroinvertebrate community. From this step, test sites (potentially disturbed sites) were matched to the appropriate reference group based on similarities in surficial geology and ecoregion. The test site was then compared with the reference grouping for which it had the highest probability of membership, determined using discriminate functions analysis. All Smith Creek Test Sites included in this study were identified as belonging to the Biological Grouping 2, and thus further analyses comparing the test sites to reference sites were conducted against the pool of Biological Grouping 2 reference sites (c.f. Phillips and McMaster, 2010). Next, attributes of the benthic macroinvertebrate assemblage were selected for comparison between test and reference sites using those adopted as part of the aquatic health feasibility study (Phillips and McMaster, 2010) and 2010 SWA State of the Watershed Reporting (SWA 2010). Specifically, the biological endpoints, or metrics, of abundance, taxa richness, Shannon’s Diversity, and community structure were summarized using Correspondence Analysis (CA) ordination axes. The measure of taxa richness used was produced as Margalef's species richness for each sample, and was a measure of the number of species making some allowance for the number of individuals. Both taxa richness and Shannon’s Diversity were calculated using PRIMER version 6 (Plymouth Marine Labs, Plymouth, UK; Clarke and Warwick, 2001). We conducted our CAs using the Biplot add-in for Excel 2009. Separate CA analyses were conducted for each Smith Creek Test Site with the combination of Reference Sites. The resulting axis 1 score was used in further test site analysis (Michelle Bowman, personal communication). Finally, TSA was used to formally evaluate the magnitude of difference between each test site and the appropriate reference sites (e.g., Bowman and Somers, 2006) based on the metrics mentioned above. TSA based on individual metrics is equivalent to evaluating whether a metric value is outside the normal range of variation in the reference sites. A non-central alpha value (ncP) > 0.95 indicates a site is significantly within reference condition and healthy, a value 22 between 0.95 and 0.05 indicates the site is outside the normal range and is stressed, while a ncP value ≤ 0.05 indicates that the site is significantly outside the normal range of reference variation and is thus impaired. 5 Results 5.1 Chemical characterization of wetlands Nutrient concentrations in the wetlands studied ranged widely (Appendix A; Table 5.1). TP ranged from 0.02 to 2.8 mg/L. Most wetlands studied were classified as eutrophic, with the exception of six which were hypereutrophic (i.e. TP > 0.6 mg/L), based on trophic classification presented in Wetzel (2001). The majority of P was typically in the organic form, with the exception of six cropped wetlands and one wood wetland that were characterized by greater proportions of orthoP. TDN in the wetlands ranged from 0.8 to 2.8 mg/L. N was predominantly present in the organic form with DON making up 96 % on average of TDN. Comparing DIN:DIP to the Redfield Ratio allows for determination of the limiting nutrient on algae growth (Rhee and Gotham, 1980; Wetzel, 2001). Eight wetlands were P limited (DIN:DIP>12:1), 46 were N limited (DIN:DIP<7), and 13 were limited by neither N nor P (i.e., DIN:DIN 7-12). DOC ranged from 19 to 55 mg/L. Salt concentrations in the wetlands studied were also greatly varied (Appendix B; Table 5.1). The wetlands were neutral to slightly basic (pH of 6.6-8.6) and ranged from fresh to brackish (SC varied from 57 to 1780 μS/cm). Maximum concentrations of major ions were HCO3 =3.8 meq/L, SO4=15.5 meq/L, Mg =13.7 meq/L, and Ca =6.8 meq/L. Distinct patterns of ion dominance groups were apparent (Figure 5.1). With the exception of one wetland (W3), all 29 wetlands with SC > 413 μS/cm were characterized by SO4>HCO3>Cl and Mg>Ca>K>Na. All but two of these wetlands were classed as crop or grassland and 41% and 34% were classed as permanent and semi-permanent, respectively. The other anion dominance pattern observed was HCO3> SO4>Cl, and 50% of these wetlands were also characterized by Mg>Ca>K>Na. The second most common (21%) cation dominance pattern for these wetlands was Ca>Mg>K>Na. The two-way MANOVA for the nutrient variable set indicated significant differences existed among land cover types (p = 0.009) and permanence classes (p = 7x10-5). However, there was no significant interaction between land cover type and wetland permanence class (p = 0.23). For the salinity variable set, there was a significant difference among land cover types (p = 5x1010 ). However, differences among permanence classes (p = 0.11) and the interaction between land cover type and wetland permanence class (p = 0.46) were not significant. 23 Figure 5.1: Distribution of cation and anion dominance groups as a function of specific conductivity for the 67 wetlands studied grouped by A) land cover and B) permanence classes. Semiperm is semipermanent wetlands. Subsequent pairwise comparison tests elucidated differences among land cover types and permanence classes for the nutrient variable set and land cover types for the salinity variable set (Table 5.1). There were significant differences among land cover types for TP and K, i.e., crop wetlands had greater TP and K than wood or grass wetlands. Significant differences among permanence classes for TP, TDN, and DOC were also found. Permanent wetlands had lower TP than seasonal and semi-permanent wetlands. In addition, TDN and DOC were higher in seasonal wetlands compared to semi-permanent and permanent wetlands. SC, Cl, HCO3, SO4, Na, Mg, and Ca concentrations were significantly lower in the wood wetlands compared to the crop and grass wetlands. 24 Table 5.1: Mean (standard error) of wetlands within each permanence and land cover class, as well as a summary of results for two-way ANOVAs. Semiperm is semipermanent wetlands. Differing letter superscripts indicate significantly different (α = 0.05) Tukey’s pairwise comparisons; * and ** denote a statistically significant difference at α = 0.05 and α = 0.01, respectively. Variable Unit pH SC uS/cm Cl meq/L HCO3 meq/L SO4 meq/L Na meq/L Ca meq/L Mg meq/L K meq/L TP mg/L orthoP mg/L TDN mg/L NO3 mg/L NH4 mg/L DOC mg/L DIN:DIP -- Permanence Permanence Class Seasonal Semiperm Permanent 7.18a 7.21a,b 7.52b (0.09) (0.09) (0.10) 446 417 601 (81) (56) (102) 7.4 5.0 4.5 (2.0) (1.1) (0.7) 101.3 96.0 104.6 (9.0) (7.4) (9.5) 99.1 93.6 156.2 (37.9) (20.8) (37.5) 14.3 11.5 21.4 (5.8) (3.6) (6.1) 28.5 23.4 25.9 (6.5) (3.1) (3.7) 28.3 26.7 47.3 (8.1) (4.9) (9.8) 21.7 19.9 16.7 (2.1) (2.7) (1.1) 0.36a 0.34a 0.11b (0.08) (0.14) (0.02) 0.06 0.09 0.04 (0.02) (0.03) (0.01) 1.58a 1.21b 1.12b (0.10) (0.07) (0.05) 0.02 0.04 0.02 (0.00) (0.01) (0.01) 0.026 0.024 0.030 (0.003) (0.003) (0.003) 38.0a 31.5 b 28.8 b (2.0) (1.2) (1.4) 4.3 6.8 5.6 (1.0) (2.4) (1.3) Two-way ANOVA F p 4.26 0.0188* 1.68 0.1952 0.67 0.5158 0.17 0.8423 1.56 0.2187 2.15 0.1260 0.08 0.9206 2.54 0.0880 1.44 0.2464 8.73 0.0005** 0.20 0.8214 11.43 0.0001** 0.16 0.8513 1.02 0.3661 8.96 0.0004** 0.30 0.7422 Land Cover Land Cover Class Two-way ANOVA Crop Grass Wood F p 7.33c 7.42 c 7.18d 1.96 0.1507 (0.08) (0.09) (0.12) 633 c 649 c 194 d 22.86 0.0000** (104) (75) (23) 6.7 c 8.1 c 1.8 d 19.77 0.0000** (0.8) (1.9) (0.2) 115.4 c 114.5 c 72.3 d 10.22 0.0002** (9.0) (7.7) (6.5) 155.5 c 180.9 c 15.9 d 22.07 0.0000** (39.0) (36.0) (6.3) 13.2 c 32.0 c 1.6 d 31.60 0.0000** (3.8) (7.0) (0.5) 33.2 c 31.9 c 12.7 d 13.15 0.0000** (4.7) (5.4) (1.2) 44.1 c 50.2 c 9.4 d 20.72 0.0000** (10.1) (8.1) (2.0) 24.8 c 16.7 d 16.9 d 4.17 0.0203* (2.7) (1.2) (1.4) 0.46 c 0.18 d 0.15 d 4.68 0.0131* (0.14) (0.04) (0.03) 0.10 0.02 0.07 1.98 0.1479 (0.03) (0.01) (0.03) 1.40 1.23 1.27 2.62 0.0817 (0.08) (0.10) (0.06) 0.03 0.02 0.02 0.54 0.5853 (0.01) (0.01) (0.01) 0.026 0.027 0.026 0.56 0.5720 (0.004) (0.003) (0.026) 34.0 32.3 31.7 0.39 0.6822 (2.1) (1.6) (1.6) 4.3 6.7 5.3 1.75 0.1833 (1.2) (2.0) (1.4) 5.2 Wetland drainage experiment The water stored in wetland LR3 increased following snowmelt from 776 m3 on October 23, 2007 to 2703 m3 on April 18, 2008. Water volume tended to decrease during rain free periods and increase following rain events throughout the 2008 open water season (Figure 5.2). Following the snowmelt period, water volume decreased ~60% throughout spring and early summer, reaching a minimum on July 7. Frequent and large rain events occurred between July 7 and August 14, which caused water volume in LR3 to increase above that in spring and remain high until the drainage experiment in November. Based on daily wetland water level fluctuations (data not shown) and precipitation data, ~6 mm/day of water was estimated to be lost from the wetland between May 1 and October 22, 2008. 25 Figure 5.2: Daily rainfall and volume in wetland LR3 prior to and during drainage. Concentration (Figure 5.3) and mass (Figure 5.4) of forms of nitrogen and phosphorus were highly seasonal variable. TP concentrations in LR3 ranged from 0.22 mg/L to below analytical detection limits (i.e., 0.01 mg/L). TP mass and concentration were both elevated April 18 and decreased sharply to lows on May 8. During the relatively rain free spring and early summer, TP concentration increased, peaking July 10. TP mass also increased during the relatively rain free period, however, it continued to increase during the July and August rain events whereas TP concentration declined in late July and increased in early August. TP concentrations and mass stabilized throughout much of August and September and began to decline in late September. TP mass and concentration spiked October 15 following a 15.8 mm rain event on October 13-14. OrthoP concentrations in the wetland ranged from 0.15 mg/L to below analytical detection limits (i.e., 0.01 mg/L). Seasonal variations in orthoP concentration and mass were similar to those for TP. TP (p = 0.42) and orthoP (p = 0.29) were not significantly correlated with Cl. The normalized mass data (Figure 5.5) show that TP and orthoP were both added to the wetland relative to Cl during the growing season. 26 Figure 5.3: Concentration of nitrogen, phosphorus, and bacteria measured in the LR3 wetland prior to the drainage experiment and in the newly constructed ditch. 27 Figure 5.4: Total mass of nitrogen, phosphorus, and bacteria measured in the LR3 wetland prior to the drainage experiment and the cumulative mass exported via the newly constructed ditch. 28 Normalized Mass 30 20 TP 10 OrthoP TKN 5 NO3 NH4 Cl 0 2.5 Normalized Mass Mg 2.0 Ca Na 1.5 K SO4 1.0 HCO3 0.5 DOC Cl 0.0 May Jun Jul Aug Sep 2008 Oct Nov Dec Figure 5.5: Mass of various water quality parameters, normalized to May 1, 2008, measured in wetland LR3. TKN concentrations in the wetland ranged from 3.7 mg/L to 1.1 mg/L. NO3 and NH4 concentrations ranged from below analytical detection limits (i.e., 0.01 mg/L) to 3.6 mg/L and 0.9 mg/L, respectively. Average concentrations of TKN, NO3, and NH4 were respectively 1.9 mg/L, 0.23 mg/L, and 0.06 mg/L. TKN, NO3, and NH4 concentrations were elevated following snowmelt and masses decreased to lows May 22, May 15 and May 1, respectively. During the relatively rain free period of April 18 to June 6, TKN concentration increased, peaking July 3. NH4 and NO3 concentrations were quite variable throughout the study period. At the beginning of the rain events (July 10) TKN and NO3 decreased substantially while NH4 increased slightly. Peaks in concentrations were reached July 17 (TKN and NH4) and July 23 (NO3). Minimum NH4 and NO3 concentrations occurred October 8 and 22, respectively. Mass of TKN and NH4 increased steadily from July 10 until August 21 and August 28, and then tended to decrease until the start of the drainage experiment. NO3 mass was highly variable during the same time period, but remained constant and low after September 24. Comparing mass ratios of dissolved inorganic nitrogen (NO3 + NH4) to dissolved inorganic phosphorus (PO4) (i.e. DIN:DIP) to the Redfield Ratio indicates that the limiting nutrient in the wetland was P in spring (Figure 5.6; Wetzel, 2001). After June 5, the limiting nutrient in the wetland was predominantly N for the remainder of the study. TKN was positively correlated with Cl (r = 0.56, p = 1x10-3), however, NH4 (p = 0.72) and NO3 (p = 0.64) were not correlated with Cl. The normalized mass data (Figure 5.5) show that over the course of the study period, NH4 was added to the wetland, NO3 was removed, and TKN was neither greatly added nor removed relative to Cl. 29 600 DIN:DIP Ratio 500 50 40 30 20 12:1 7:1 10 0 May Jun Jul Aug Sep Oct Nov 2008 Figure 5.6: Mass ratio of DIN:DIP (NO3+NH4:PO4) measured in the LR3 wetland prior to its drainage. Values below seven indicate N limiting conditions and values above 12 indicate P limiting conditions (Wetzel, 2001). Variations in density (Figure 5.3) and loads (Figure 5.4) of both T. coli and E. coli were very similar throughout 2008, and as such only trends in loads are summarized. Following the snowmelt period, loads of T. coli and E. coli in LR3 decreased until May 28, then increased until June 26 and June 19, respectively. Minimums were reached July 10. Loads of T. coli and E. coli then increased during the mid summer rain events. T. coli and E. coli loads then generally decreased until the start of the drainage experiment. A secondary peak in E. coli and T. coli occurred October 8. Neither T. coli (p = 0.12) nor E. coli (p = 0.50) were significantly correlated with Cl. Seasonal variations in major ion and DOC concentrations were similar throughout summer 2008, with the exception of HCO3 (Figure 5.7). DOC (r = 0.88, p = 3x10-11) and all major ions (r = 0.93-0.98, p = 3x10-25-7x10-14), with the exception of HCO3 (p = 0.32), were significantly correlated with Cl. Following snowmelt and during the relatively rain free spring and early summer, ion and DOC concentrations generally increased, peaking around July 10. Ion concentrations then reached minimums on July 30 during rain events and then increased until the start of the drainage experiment. In contrast, HCO3 concentration did not noticeably increase between the snowmelt and the end of July. Steady increases in HCO3 concentrations were observed between August and November. Trends in major ion and DOC mass (Figure 5.8) differed from trends in concentration. DOC and major ion mass decreased in the wetland during the relatively rain free spring and early summer until July 10 then increased on average by a factor of 2.4 and peaked ~August 28. Major ion and DOC mass mostly decreased in September and increased in October. The ion dominance pattern in the LR3 wetland was SO4>HCO3>Cl and Mg>Ca>Na>K, which remained constant throughout the study period in 2008. Seasonal variation in pH (Figure 5.7) was similar to that observed for concentrations of major ions. However, pH peaked at 9.5 on July 3 and reached a minimum value of 7.4 August 14. The normalized mass data (Figure 5.5) show that DOC, HCO3, and Ca were added to the wetland relative to Cl. At the time of the drainage experiment, the LR3 wetland was covered by ~8 cm of ice. As a result, only 81% of the water was in liquid form. The wetland volume decreased rapidly when the drainage ditch was completed (Figure 5.2). After 4 hrs of drainage, 30% of the water (total) had exited the wetland via the ditch. Concentrations of N, P, bacteria, and major ions were greater in the drainage ditch than those measured in the wetland at the start of drainage (Figures 5.3 and 5.4). DOC concentrations in the ditch were less than that measured in the wetland at the start of drainage, not including the final sampling instance, 23 hr after the start of drainage. The 30 pH was consistently lower in the newly constructed drainage ditch than in the wetland at the start of drainage (Figure 5.7). Figure 5.7: pH and concentration of DOC and major ions measured in the LR3 wetland prior to the drainage experiment and in the newly constructed ditch. 31 Figure 5.8: Total mass of DOC and major ions measured in wetland LR3 prior to the drainage experiment and the cumulative mass exported via the newly constructed ditch. Over the course of the drainage experiment, concentrations of N, P, and DOC generally increased in the drainage ditch. However, concentrations of TP, NO3, and NH4 were greater 1 hr after the start of drainage compared to concentrations measured 4 and 6 hr after the start of drainage. Total mass exported from LR3 for each constituent during the experiment is summarized in Figures 5.4 and 5.8. The total masses of TP, orthoP, NO3, and NH4, and the total number of E. coli and T. coli colonies exported via the drainage ditch exceeded those estimated in the wetland at the start of the drainage experiment by a factor of 1.2, 2.1, 19.0, 4.3, 19.4, and 18.9, respectively. Total masses of TKN, DOC, and major ions exported via the drainage ditch were less than those estimated in the wetland at the start of the drainage experiment by factors ranging from 0.4 to 0.6. Results for solutes that were significantly correlated (α = 0.05) with distance along the drainage ditch and that also had at least marginally significantly different (α = 0.10) slopes than 32 Cl are shown in Figure 5.9. Slopes that differ from Cl suggest that the nutrient or ion experienced biotic processing, sorption or release along the ditch length. Slopes of DOC (1 hr), orthoP (1 hr, 4 hr, and 23 hr), T. coli (4 hr), HCO3 (6 hr), and NH4 (6 hr and 23 hr) were less steep than the Cl slope. Slopes of HCO3 (4 hr), T. coli (23 hr), and NO3 (23 hr) were steeper than the Cl slope. Normalized Cl concentrations were not significantly (α = 0.05) correlated with distance 4 hr and 6 hr after the start of drainage. Figure 5.9: Slopes of normalized concentrations measured along the newly constructed ditch draining the LR3 wetland at: a) 1 hr, b) 4 hr, c) 6 hr, and d) 23 hr after the start of the drainage experiment. Concentrations were normalized by dividing the concentration at each sample point along the ditch by the concentration at the first sampling point (DR1) in the ditch. A value of one was then subtracted to set the intercept to zero. Only constituent slopes that were at least marginally different (α = 0.10) from the chloride slope and that had significant (α = 0.05) linear relationships between normalized concentration and distance are shown. P values indicate the level of significance for slopes differing from chloride, suggesting that a portion of the variability is due to biotic processes or sorption. 33 5.3 Comparison of ditches and spills Ditches were on average 71 % longer (p = 1x10-4), 12 % narrower (p = 0.035), and tended to have 33% higher flow velocities (p = 0.016) than spills (Figure 5.10). Constituent concentrations also differed between ditches and spills (Figure 5.11). Specifically, TDN (p = 0.003), DOC (p = 0.007), HCO3 (p = 0.023), K (p = 0.001), and Ca (p = 0.010) concentrations were greater in ditches than spills. NO3 (p = 0.058) and NH4 (p = 0.055) concentrations tended to be higher in ditches than spills. Loads of TDN (p = 0.038), NO3 (p = 0.034), and K (p = 0.048) were significantly greater in ditches than spills (Figure 5.12). In contrast, loads and concentrations of TP and orthoP were not significantly different between ditches and spills. Ttest comparisons of inlet and outlet loads showed that solute loads did not change along the length of ditches or spills (0.29 < p < 0.97), with the exception of orthoP which tended to be greater (p = 0.058) at spill outlets than inlets. Figure 5.10: Mean and standard error of ditch and spill physical properties where * and ** denote statistically significant differences at α = 0.05 and 0.01, respectively. Figure 5.11: Mean and standard error of A) nutrients and DOC, and B) major ions, SC, and pH in ditches and spills where * and ** denote statistically significant differences at α = 0.05 and 0.01, respectively. 34 Figure 5.12: Mean and standard error of ditch and spill nutrient and DOC loads where *denotes a statistically significant difference at α = 0.05. 5.4 Stream water quality Typical of the northern prairies, the 2009 snowmelt period was short, lasting 28 days. Thaw (and subsequent flow) at the low drainage subbasin was delayed by about a week compared to that at the Smith Creek outlet and its other subbasins. Maximum stream discharge and concentrations of the water quality parameters measured, with the exception of SC and E. coli, occurred during onset of the spring freshet (Figure 5.13). Ratios of DIN (NO3 + NH4) to DIP (PO4) measured near the Smith Creek outlet indicate N limiting conditions on nearly all sampling dates in 2009 (Figure 5.14). During the recession limb of the hydrograph, the dominant anion switched from HCO3 to SO4 (Figure 5.15), coincident with an increase in SC to >520 mS/cm (Figure 5.13). This switch was observed in each of the studied subbasins. There was a trend toward higher discharge, and nutrient and salt concentrations at the subbasins with medium high and high wetland drainage compared to the subbasin with low wetland drainage (Figure 5.13). After normalizing concentration data for variations in streamflows and basin areas, differences in export coefficients among subbasins with variable wetland drainage were found for the 2009 freshet (Table 5.2). Subbasins characterized by medium high and high wetland drainage had significantly higher mean export coefficients of TP, orthoP, HCO3, SO4, Mg, and Ca than the subbasin with low wetland drainage. Because of its large size, the subbasin characterized by medium high wetland drainage had the highest total export of nutrient, salts and bacteria (Figure 5.16). However, total export of these water quality constituents at Smith Creek outlet, which is located just upstream of the town drainage (Langenburg Creek), was higher than the sum of the three studied subbasins. 35 1.0 0.5 45 Low Drainage 1.4 1.2 1.0 0.8 0.6 0.4 0.2 0.0 10.00 8.00 6.00 4.00 2.00 0.50 15 10 5 2.00 1400 1200 1000 800 600 400 200 0 SC (μS/cm) NH4 (mg/L) 90 80 20 0 1.00 0.50 0.10 0.00 Jun 30 0.00 0.05 May 35 20 1.50 Apr 40 25 0.25 0.0 OrthoP (mg/L) 50 Smith Creek Outlet High Drainage Mod. High Drainage E. coli (MPN/100 mL) 1.5 NO3 (mg/L) TP (mg/L) 2.0 14 12 10 8 6 4 2 0 DOC (mg/L) TDN (mg/L) Discharge (cms) 7 6 5 4 3 2 1 0 Apr May Jun Apr May Jun 2009 2009 2009 Figure 5.13: Discharge and water quality concentrations during the 2009 freshet at sites characterized by differing watershed drainage: high (TV1), medium high (SC4), and low (SC5). No data are presented prior to April 8, 2009 due to ice at the gauge station. 36 18 16 DIN:DIP 14 12:1 12 10 7:1 8 6 4 2 0 Apr May Jun 2009 Figure 5.14: Ratios of DIN (NO3 + NH4) to DIP (PO4) measured near the Smith Creek watershed outlet (SC3) during 2008 and 2009. Values below seven indicate N limiting conditions while those above 12 indicate P limiting conditions (Wetzel, 2001). 37 Basin Outlet High Wetland Drainage Medium High Wetland Drainage Low Wetland Drainage Figure 5.15: Piper plots for: basin outlet (SC3) and sub-basins characterized by high (TV1), medium high (SC4), and low (SC5) wetland drainage. Blue and red circles indicate samples obtained during the snowmelt period (March 23 – April 19, 2009) and following snowmelt (April 25 – May 26, 2009), respectively. 38 Table 5.2: Mean (standard error) stream export coefficients for the 2009 freshet by site. Differing superscripts indicate significant difference at α = 0.05, assessed using a one-way ANOVA and the Holm-Sidak method for multiple pair-wise comparisons. Variable Units Wetland Drainage Intensity ANOVA results Low Med. High High F P Q m3/day/km2 392a 1212b 1293b 5.24 0.011 (151) (154) (208) TP kg/day/km2 0.06a 0.60b 0.55b 5.81 0.008 (0.02) (0.07) (0.13) OrthoP kg/day/km2 0.01a 0.45b 0.34b 6.94 0.003 (0.01) (0.06) (0.09) DOC kg/day/km2 14.1a 41.2a,b 49.6b 4.59 0.019 (5.53) (4.83) (9.08) TDN kg/day/km2 0.93a 2.31a,b 6.02b 5.49 0.010 (0.42) (0.27) (1.45) NO3 kg/day/km2 0.29a 0.63a 2.34a 3.37 0.048 (0.19) (0.1) (0.79) NH4 kg/day/km2 0.1a 0.1a 1.1b 6.38 0.005 (0.06) (0.01) (0.31) Cl kg/day/km2 1.7a 8.5a 8.0a 3.54 0.042 (0.4) (0.7) (2.2) HCO3 kg/day/km2 34.0a 123.7b 120.9b 4.28 0.024 (11.2) (17.3) (24.2) SO4 kg/day/km2 26.8a 95.0b 82.1b 3.92 0.031 (7.1) (11.2) (18.6) Na kg/day/km2 3.0a 10.4b 6.3a,b 6.56 0.004 (1.1) (1.3) (1.3) K kg/day/km2 6.2a 24.7a,b 29.3b 4.98 0.014 (2) (3.5) (5.7) Mg kg/day/km2 5.4a 23.1b 21.7b 6.43 0.005 (1.8) (2.7) (3.8) Ca kg/day/km2 9.5a 36.0b 40.4b 5.36 0.010 (3.5) (3.4) (7.8) 39 18 10000 300 a) 1000 b) 16 250 800 12 200 10 150 8 6 100 Ions (kg x1000) 14 DOC (kg x1000) 4000 Nutrients (kg x1000) 6000 3 Water (m x1000) 8000 4 2000 600 400 200 50 2 0 0 0 Q TP orthoP TDN NO3 NH4 0 DOC Cl Na K Mg Ca 2.0e+14 2.5e+11 c) 2.0e+11 High Medium High Low Unassessed (SC3 subbasin) 1.5e+14 1.5e+11 1.0e+14 1.0e+11 T. coli (MPN) E. coli (MPN) HCO3 SO4 5.0e+13 5.0e+10 0.0 0.0 E.coli T.coli Figure 5.16: Total exports of: a) water (Q), nitrogen (TDN, NO3, and NH4) and phosphorus (TP and orthoP); b) major ions; and c) bacteria (E. coli and T. coli) during the 2009 freshet (3 April - 26 May) at sub-basins characterized by low, medium high, and high wetland drainage. Exports for the SC3 subbasin (c.f. Figure 4.3) were calculated by subtraction of exports for the three subbasins from that for the whole watershed. Total height of the bars indicates export for the whole Smith Creek watershed. 5.5 Description of macroinvertebrate assemblages Acquired were 17 samples representing eight sites in 2008, and 15 samples representing eight sites across two time periods in 2009. Because we were unable to find suitable sampling sites through much of the upstream Smith Creek watershed, we focused sampling efforts in 2008 on replication at the site level. Specifically, six samples were produced from the Smith Creek at Marchwell (SWA_2008_16) site, eight samples were produced from the Smith Creek East of Langenburg site (SWA_2008_17), three from the Langenburg Creek East of Langenburg Site (SWA_2008_18), and a single sample from the Smith Creek at Werle Farm Site (SWA_2008_19). Sample number 3 from the Langenburg Creek East of Langenburg and samples 2, 3, and 4 from the Smith Creek at Werle Farm all contained too few macroinvertebrates for further analysis. Based on the consistency of the results at the Marchwell and Langenburg sites (SWA_2008_16 and SWA_2008_17 respectively, Table 5.3) in 2008, and constraints of budgeted sorting and identification costs, only a single sample from each of the sites monitored was analyzed in 2009. However, the remaining three samples from each of the sites have been retained for future analysis. Overall, 27,985 benthic macroinvertebrates were collected, representing 106 unique taxa (Appendix C). This study was dominated by the fairy shrimp (Anostraca), the dipteran midge larvae (Chironomidae), freshwater worms (Oligochaeta), nematodes (Nematoda), one genera of 40 mayfly (Caenis) and blackfly larvae (Simuliidae). There were no surprises or occurrences of unexpected or rare taxa based on previous understanding of prairie streams in the area (see Phillips et al., 2008). The 2008 results indicate that the Smith Creek at Marchwell is a healthy site, and not significantly different from reference conditions (Table 5.3). However, the Smith Creek east of Langenburg produced three of seven sites which were stressed, the Langenburg Creek was overall severely impaired, and the Smith Creek at Werle Farm was stressed and close to the impairment threshold (Table 5.3). A closer look at the nature of the ecosystem health conclusions based on the individual metrics that make up this TSA identifies that abundance is consistently within reference condition (Table 5.4). Table 5.3: Sites sampled for ecosystem health in 2008. Smith Creek Smith Creek Site Code Watershed Site SWA_2008_16_1 Smith Creek North of Marchwell SWA_2008_16_2 Smith Creek North of Marchwell SWA_2008_16_3 Smith Creek North of Marchwell SWA_2008_16_4 Smith Creek North of Marchwell SWA_2008_16_5 Smith Creek North of Marchwell SWA_2008_16_6 Smith Creek North of Marchwell SWA_2008_17_1 Smith Creek East of Langenburg SWA_2008_17_2 Smith Creek East of Langenburg SWA_2008_17_3 Smith Creek East of Langenburg SWA_2008_17_4 Smith Creek East of Langenburg SWA_2008_17_6 Smith Creek East of Langenburg SWA_2008_17_7 Smith Creek East of Langenburg SWA_2008_17_8 Smith Creek East of Langenburg SWA_2008_18_1 Langenburg Creek East of Langenburg SWA_2008_18_2 Langenburg Creek East of Langenburg SWA_2008_18_4 Langenburg Creek East of Langenburg SWA_2008_19_1 Smith Creek at Werle Farm 41 TSA Results D ncP 1.86 1.00 1.50 1.00 0.89 1.00 1.28 1.00 0.87 1.00 1.19 1.00 2.39 0.99 1.09 1.00 3.18 0.62 2.21 1.00 4.03 0.12 2.76 0.89 0.93 1.00 7.75 0.00 33.02 0.00 4.09 0.11 4.17 0.09 Overall Health Healthy Healthy Healthy Healthy Healthy Healthy Healthy Healthy Stressed Healthy Stressed Stressed Healthy Impaired Impaired Stressed Stressed Table 5.4: Metric values and their probability of difference from reference (Metric Significance) Metric Values SWA Taxa Shannon's CA Site Abundance Richness Diversity Axis 1 SWA_2008_16_1 453 4.42 2.56 0.87 SWA_2008_16_2 235 4.21 2.46 1.02 SWA_2008_16_3 120 3.55 2.24 1.11 SWA_2008_16_4 396 4.01 2.36 0.98 SWA_2008_16_7 58 3.20 2.37 1.15 SWA_2008_16_8 365 3.73 2.08 0.85 SWA_2008_17_1 1295 4.88 2.15 0.80 SWA_2008_17_2 142 3.63 2.01 1.02 SWA_2008_17_3 844 4.75 2.28 0.38 SWA_2008_17_4 483 4.69 2.25 0.79 SWA_2008_17_6 635 4.49 2.28 0.15 SWA_2008_17_7 311 3.14 2.32 0.49 SWA_2008_17_8 496 2.74 1.78 0.75 SWA_2008_18_1 634 2.33 1.54 2.57 SWA_2008_18_2 732 1.21 0.28 -6.63 SWA_2008_18_4 1634 1.89 1.48 1.64 SWA_2008_19_1 28 0.00 0.00 0.87 SWA_2009_10_1 196 1.14 0.84 3.65 SWA_2009_11_1 224 1.29 0.54 0.88 SWA_2009_4_1_May 1102 1.86 1.12 -0.43 SWA_2009_4_3_April 1068 4.45 1.75 0.85 SWA_2009_5_1_April 192 3.23 2.22 1.00 SWA_2009_5_1_May 337 6.36 2.76 0.82 SWA_2009_6_1_May 523 2.56 1.21 0.93 SWA_2009_6_2_April 740 2.88 1.39 0.90 SWA_2009_7_1_May 424 3.31 1.66 0.91 SWA_2009_7_2_April 546 3.12 1.54 0.89 SWA_2009_8_1 70 0.71 0.39 0.86 SWA_2009_9_1 576 3.46 2.58 0.82 42 Abundance 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 Metric Significance Taxa Shannon's Richness Diversity 0.85 1.00 0.97 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 0.30 1.00 1.00 1.00 0.45 1.00 0.52 1.00 0.77 1.00 1.00 1.00 1.00 1.00 1.00 1.00 0.82 0.01 1.00 1.00 0.01 0.00 0.74 0.43 0.89 0.05 1.00 0.95 0.82 1.00 1.00 1.00 0.00 0.92 1.00 0.99 1.00 1.00 1.00 1.00 1.00 1.00 0.24 0.02 1.00 1.00 CA Axis 1 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 0.89 1.00 1.00 0.00 0.00 0.86 1.00 1.00 1.00 0.02 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 Early 2009 spring sampling results indicate that all stream sites in the Smith Creek were healthy regardless of drainage stress (Table 5.5). However, May samples show that SC 4 had become impaired and SC 5 had become stressed (Table 5.6). Ditches in the 2009 study ranged from healthy at DT 3, stressed in DT 4, DT 2 and DT 1 (Table 5.6). Table 5.5: Test Site Analysis results for sites from April 13, 2009 in the Smith Creek Watershed. D indicated the magnitude of effect or difference from reference, while ncP is the measure of significance. Smith Creek Drainage TSA Results Overall Watershed Site Stress D ncP Health Healthy Smith Creek at SC4 Medium High Drainage 2.46 0.98 Smith Creek at SC5 Low drainage 0.66 1.00 Healthy Healthy Thingvala Creek at TV1 High Drainage 1.51 1.00 Smith Creek at SC3 Very High Drainage 1.23 1.00 Healthy Healthy Ditch at DT3 Ditch 1.76 1.00 Stressed Ditch at DT1 Ditch 3.00 0.75 Stressed Ditch at DT4 Ditch 2.99 0.75 Stressed Ditch at DT2 Ditch 3.30 0.52 Table 5.6: Test Site Analysis results for sites from April 13, 2009 in the Smith Creek Watershed. D indicated the magnitude of effect or difference from reference, while ncP is the measure of significance. Smith Creek Watershed Site Smith Creek at SC4 Smith Creek at SC5 Thingvala Creek at TV1 Smith Creek at SC3 Drainage Stress Medium High Drainage Low drainage High Drainage Very High Drainage TSA Results D ncP 5.54 0.00 3.88 0.18 1.82 1.00 1.28 1.00 Overall Health Impaired Stressed Healthy Healthy 6 Discussion 6.1 Wetland water quality characterization Researchers have previously used ion dominance patterns and SC (as a proxy for net groundwater seepage rates) as indicators of wetland permanence (Sloan, 1972; Millar, 1976; Driver and Peden, 1979). Although exchanges between the wetland and deep groundwater have only a minimal influence on the water balance, their effect on salinity can be important as the direction of flow determines whether salts accumulate in the wetland due to upward flow or are leached out of the wetland by outward flows (Hayashi et al., 1998b; van der Kamp and Hayashi, 2009). SC has thus been used as an indicator of relative position of wetlands along regional groundwater flow paths (LaBaugh and Swanson, 2004). However, results presented herein show that neither SC nor ion dominance can be used to distinguish among wetland permanence classes at Smith Creek watershed. Although five of the six highest SC measurements were obtained from permanent wetlands, there was no significant difference among permanence classes for SC or solute concentrations. The high within permanence class variation likely occurred because localized shallow groundwater flow systems dominate the groundwater hydrology of prairie 43 potholes (Hayashi et al. 1998a; van der Kamp et al., 2008). Thus, linkages between SC and permanence may only be appropriate for comparisons made at the regional scale, and the focus of this study was at a smaller spatial scale. Furthermore, solute concentration and SC were likely diluted during the sampling period by snowmelt runoff, which would mask differences between permanence classes. Differences among permanence classes may however become more pronounced later in the season due to evapoconcentration (Rózkowska and Rózkowski, 1969; LaBaugh et al., 1987). Other studies have mostly conducted their sampling campaigns midsummer (e.g. Arts et al., 2000); however, wetlands included in this study were not sampled later in the season because seasonal wetlands typically become dry. The lack of difference among permanence classes may also be attributed to the fact that classes are not necessarily strictly distinct and static. Instead, potholes within each permanence class can show sharp differences in time of inundation and in water depth between periods of drought and deluge (Johnson et al., 2004), conditions common in the prairies. The wetlands sampled grouped distinctly into low SC and HCO3 dominated groups, largely typical of the wood wetlands, and relatively high SC and SO4 dominated groups, largely typical of grass and crop wetlands. An association between SC and anion dominance has been previously observed in prairie lakes and wetlands (Rawson, 1944; Rózkowska and Rózkowski, 1969; Barica, 1975; LaBaugh et al., 1987, Swanson et al., 1988). As solutes become increasingly concentrated by evaporation in closed basins, saturation levels for calcium and magnesium carbonates are reached first, and then of calcium sulfate, causing the minerals to precipitate in that order (Holland, 1978). The significantly lower SC and solute concentrations measured in the wooded areas may be a result of a high order topographic position of these wetlands in the Smith Creek watershed and thus their likely position near the top of regional groundwater flow paths (Tóth, 1963; Rózkowski, 1969; Devito et al., 2000). Topography, however, is not always a good indicator of water table elevations in the prairies (van der Kamp, pers. comm.). Transects studies by Barica (1978) indicate that closed basin lake salinity varied considerably regardless of the general topographic slope. Also, wood wetlands may, in part, have lower Ca, K, and Mg concentrations than grass or crop wetlands because they uplands are comprised primarily of aspen (Populus tremuloides), which are known to store these solutes in their standing biomass (Wang et al., 1995). SC and solute concentrations measured throughout Smith Creek were lower than maximum values measured at other lakes and wetlands in the prairie pothole region (Rózkowska and Rózkowski, 1969; Barica, 1975; LaBaugh et al., 1987; Detenbeck et al., 2002; Waiser, 2006) and were comparable to values observed by Nicholson (1995) in a northern Alberta transition zone between semi-arid prairie and moister boreal forest. These lower concentrations may be the result of dilution caused by the relatively higher annual precipitation at Smith Creek watershed compared to other parts of the prairie pothole region (Millet et al., 2009), which would minimize the precipitation to evaporation ratio and the effects of evapoconcentration. Additionally, the mid-summer precipitation events that occurred in the region during the 2008 summer prior to wetland sampling caused many potholes to fill with dilute rain water and remain relatively full at the time of freeze-up. SC was also likely low because samples were collected in late spring shortly after the wetlands have filled with dilute snowmelt runoff and before solutes have become concentrated by evaporation. Dilute salt concentrations in precipitation have been measured 220 km southwest of Smith Creek at Bratt’s Lake station, which is part of the Canadian Air and Precipitation Monitoring Network (CAPMoN, 2007). Although not measured, it is likely that some of the HCO3 dominated wetlands with SC near 400 μS/cm became SO4 44 dominated later in summer. This inference is based on the work of LaBaugh et al. (1987) and Detenbeck et al. (2002), which showed some temporal progression in seasonal wetland ion dominance patterns. Some of the spatial variation in some wetland nutrients can be attributed to variations in land cover and permanence classes. The higher concentrations of TP, TDN and DOC in seasonal wetlands likely results from the pronounced periods of flooding and drying that affect them. Mineralization of organic matter is enhanced during dry periods, resulting in leaching of nutrients from standing dead litter and sediment when the next inundation occurs (Bärlocher et al. 1978; Neill, 1995; Baldwin and Mitchell, 2000; Aldous et al., 2005). In contrast, the lack of vegetation and continuous flooding of sediments located within the open-water zones of permanent wetlands and some semi-permanent wetlands would not lead to the same annual release of nutrients (Brinson et al., 1981). The near continuous flooding of semipermanent and permanent wetlands suggests that conditions may be sufficiently reduced for denitrification to occur, also leading to comparatively lower total N concentrations (Neely and Baker, 1989; Crumpton and Goldsborough, 1998). Seasonal wetlands have been shown to have higher TP concentrations compared to semi-permanent ones due to leaching from standing dead plants in the marsh zones surrounding them (LaBaugh and Swanson 2004). Further, seasonal wetlands are often cropped when they are dry, thus they can have higher N and P concentrations than semipermanent or permanent wetlands because of direct fertilizer application (Cowardin et al., 1981). In addition to differences in wetland nutrients among wetland permanence classes, differences in some nutrients were found among wetland land cover types. The greater TP and K concentrations in crop than wood and grass wetlands may have resulted from fertilizer inputs being transported to wetlands from their cropped watersheds (Hansen et al., 2002; Little et al., 2007; Tiessen et al., 2010) during snowmelt. The finding that N and DOC did not differ significantly among land cover types was surprising given that other researchers have shown that wood litter contains fewer nutrients and DOC than grass litter (Fuller and Anderson, 1993; Köchy and Wilson, 1997), and that nutrients stay stockpiled in the woody biomass located throughout wood wetland watersheds for extended periods of time (Wang et al., 1995). The lack of significant difference in N among land cover types may be attributed to the fact that the majority of wetlands sampled, similar to shallow prairie lakes, were eutrophic and characterized by low DIN:DIP ratios and were thus N limited (Barica, 1990; Hall et al., 1999). The greater TP and K concentrations measured in crop than wood and grass wetlands may also be the result of varying amounts of surface runoff from different land cover types. Surface runoff and the nutrients transported with it are likely greatest in cropped areas partly because of higher overland flows during spring (van der Kamp et al., 2003; Bodhinayake and Si, 2004). The infiltration potential of frozen soils is higher if the soils are dry and have a well developed macropore structure compared to saturated soils with poor macropore development, which have very low infiltration potential (Gray et al., 2001). Cultivation reduces macroporosity and infiltration capacity (Bodhinayake and Si, 2004), which increases infiltration of snowmelt water and rain, and decreases surface runoff to depressions (van der Kamp and Hayashi, 2009). Further, snow and rain interception by forest canopies reduce snowpacks and throughfall (Pomeroy et al., 1997). Consequently runoff in wood areas is likely lower than grass and crop areas. It is unlikely that the observed differences in wetland salinity and nutrient concentrations among land cover types were attributed to differences in soil characteristics. While spatial 45 variation in soil types occur across the Smith Creek watershed (Figure 4.1), the majority of wetlands resided in soil units characterized by orthic or calcareous soils on lower slopes or poorly drained depressions. In all, 100%, 46%, and 50% of crop, grass, and wood wetlands, respectively, were located in soil units with orthic soils in depression, 17% and 36% of grass and wood wetlands were located in soil units with calcareous soils in depression, and 38% of grass wetlands and 14% of wood wetlands were located in soil units with carbonated and mixed (orthic and calcareous) soils in depressions. There was a trend, however, between SC-SO4 dominance and soil type. Forty one percent of wetlands characterized by high SC and SO4 dominance were located in soil units characterized by slightly larger proportions of sandy loams (OxWh2, OxWs2, and Yk12). These soils are expected to have a relatively higher hydraulic conductivity which means that the wetlands would likely experience greater shallow groundwater and salt fluxes. Results also show that the interaction between land cover and permanence classes did not significantly influence nutrient or salt water quality parameters. This result is not surprising given that the control mechanisms on water quality parameters attributable to wetland permanence and land cover type are likely additive. The lack of significant interaction may have also resulted from the relatively high variability of wetland water quality measured. Other factors not considered in this study that could also influence prairie wetland water quality include grazing, cultivation, and tillage practices, varying groundwater fluxes, and the presence of willow rings surrounding wetlands. 6.2 Factors controlling temporal patterns in wetland water quality The LR3 wetland effectively trapped N, P, DOC, coliforms, and major ions during runoff events and exchanged them with the surrounding uplands between events prior to the construction of the drainage ditch. Intensive temporal measures at the LR3 wetland suggest hydrological processes are the dominant control on all the studied ionic water quality parameters, except for HCO3. However, the lack of significant correlations with Cl for most nutrient variables and differing seasonal dynamics of concentrations, masses and normalized masses between major ions and DOC compared to N, P, and bacteria suggest there are other processes important in determining wetland nutrients. This concept of differing control mechanisms for major ions and nutrients was hypothesized by LaBaugh et al. (1987). Seasonal fluctuations of ions and DOC appear to be primarily linked to hydrological processes due to the significant correlations with Cl. Instead, variable seasonal dynamics observed between major hydrological events and the lack of significant correlations between Cl and TP, orthoP, NH4, NO3, and bacteria suggests that wetland N, P, and bacteria are linked to both hydrological processes and biotic/sorption processes. For example, sequences of algae and plant uptake and decay, microbial processing (i.e. mineralization, nitrification, and denitrification), sedimentation, and waste from waterfowl and semi-aquatic mammals have been all shown to influence wetland nutrient availability (Barica, 1974; Neely and Baker, 1989; Neill, 1995; LaBaugh and Swanson, 2004). Removal mechanisms for bacteria in wetlands include sediment retention and natural die-back (Hemond and Benoit, 1988; Auer and Niehaus, 1993). Temporal variations in solute concentrations and masses in the wetland can largely be attributed to the hydrologically isolated nature of the wetland. The estimate of ~6 mm/day of water lost from the wetland is within the range of those for other isolated prairie wetlands (1.7 – 7.4 m) via shallow groundwater infiltration and evapotranspiration (Shjeflo, 1968; Millar 1971; Woo and Roswell, 1993; Hayashi et al. 1998a; Su et al., 2000). Shallow, lateral flows, driven by 46 hydraulic gradients and transpirative demand by upland plants, occur at wetland margins within the top 5-6 m of till in this landscape where the hydraulic conductivity is relatively high due to fractures (Hayashi et al., 1998a). These flows transport solutes with them (Hayashi et al. 1998b; Parsons et al., 2004), explaining the reduction in mass of most water quality constituents in the wetland between rain events. Transport with lateral flows can effectively concentrate solutes in uplands (Arndt and Richardson, 1993; Winter and Rosenberry, 1995). They are then likely to be leached and cycled back to the wetland during snowmelt and rainfall runoff events. Precipitation events can also cause the water table beneath the wetland margin to rise above the pond level causing a reversal in shallow groundwater flow toward the pond (Gerla 1992; Winter and Rosenberry, 1995; Hayashi et al., 1998b; Parsons et al., 2004), which would also transport solutes back to the wetland. Daily water level increases in the LR3 wetland exceeded daily precipitation by >5 mm on 14 days during the study period, indicating that surface and/or subsurface runoff likely contributed to the increase in wetland water storage. Specifically, runoff contributions to the wetland likely occurred on at least nine days during the midsummer rain events as well as on May 28, June 12, June 23, October 6 and October 12. Surface and/or subsurface runoff are thus likely responsible for the high solute masses and bacteria loads observed during the snowmelt period and the increases that occurred following the rain events. Concentrations of DOC and major ions increased during rain free periods likely due to evapoconcentration and mass decreased likely due to the transport with water out of the wetland by means of shallow groundwater seepage (Hayashi et al. 1998b; Waiser, 2006), as is described above. Data from Bratt’s Lake station of the Canadian Air and Precipitation Monitoring Network (CAPMoN), which is 220 km southwest of the Smith Creek watershed, show precipitation is dilute with regards to major ions (CAPMoN, 2007). As a result, inputs during the midsummer rain events probably led to DOC and major ion dilution. It is thus likely that the increases of DOC and major ions mass measured immediately following rain events were caused primarily by runoff and shallow groundwater inputs. Following snowmelt, the wetland had high nutrient concentrations. While transient, elevated N and P concentrations immediately following snowmelt are important to note because ditched wetlands drain at this time. Similar to results from Batt et al. (1989) and LaBaugh and Swanson (2004), these elevated N and P concentrations coincided with periods of use by breeding waterfowl. They also are coincident with a 2.5-fold increase in wetland volume, relative to the previous fall. Freshly flooded above ground litter leaches stored N and P to the water column (Reddy and Patrick, 1975; Davis and van der Valk, 1978; Neill, 1995) and nutrients accumulated in soils of the wetland periphery are released upon re-wetting (Reddy and Patrick, 1975; Murkin et al., 2000; Aldous et al., 2005). Soils in the region are naturally nutrient rich, especially in P (Anderson, 1988). In addition, nutrients were likely released from the cropped areas of the catchment that received fertilizer in fall 2007 and May 2008, as has been shown to occur in other agricultural systems (Hansen et al., 2002; Little et al., 2007). The midsummer runoff events led to increased TKN and TP mass in the LR3 wetland. Prior to the rain events orthoP represented 13% on average of TP and increased to 60% following the rain events. This increase in the proportion of TP as orthoP may have resulted from the transport of orthoP with sediment (Neely and Baker, 1989) during overland flows. Although NH4 and NO3 are predominantly transported with surface runoff and subsurface flows, respectively (Neely and Baker, 1989), their percentages of TN were similar before and during the midsummer rains. Average concentrations of NO3 and NH4 in precipitation at the CAPMoN 47 Bratt’s Lake station (2007) are 2.1 mg/L and 0.95 mg/L, respectively. These concentrations are much greater than those measured in the wetland throughout the study period. Thus wet deposition in addition to upland runoff and leaching from soil and plants located within the freshly flooded wetland periphery could have contributed to the increase in nutrient concentrations and mass loads measured in midsummer. The normalized mass data (Figure 4.4) suggest that reductions in TP, orthoP, and NH4 mass following the midsummer rain events were due to biotic uptake and/or biogeochemical reactions rather than hydrological processes. Furthermore, these data also suggest that NO3 was removed from the wetland relative to Cl, likely by biotic uptake and denitrification. Although measurements of nitrogen cycling in prairie wetlands are limited (e.g. Moraghan, 1993), researchers such as Neely and Baker (1989) have noted that conditions are likely suitable for denitrification to occur: anaerobic conditions and the presence of a large organic carbon stock. Temporal variations in HCO3 were likely influenced by carbonate equilibrium relationships as Heagle et al. (2007) identified carbonate mineral dissolution to be an important geochemical reaction in a recharge prairie wetland. Normalized mass data showed that proportions of HCO3 and Ca became elevated relative to Cl. This result suggests that the increase in these ions was not attributable to water inputs, marked by Cl variations, alone. Sulfate reduction was also identified as a key geochemical reaction by Heagle et al. (2007). The proportional mass data did not indicate that SO4 was removed from the wetland differently than was Cl, meaning sulfate reduction was likely not an important driver of SO4 concentrations at this site. A major non-point source of disease causing coliforms and indicator coliforms in agricultural landscapes is runoff containing animal wastes from pastures or fields fertilized with manure (Hyland et al., 2003). While the wetland catchment was not fertilized with manure over the course of the study period, the high coliform densities measured have been observed in comparable agricultural systems following snowmelt and large precipitation events (Ontkean et al., 2003). Semi-aquatic mammals and waterfowl that commonly occupy the wetland can also contaminate wetland water and uplands with fecal matter (Hyer and Moyer, 2004; Kadlec et al., 2007). Muskrats occupied the wetland as evidenced from newly constructed lodges some time between June 25 and September 24, 2008. 6.3 Water quality characteristics of a newly constructed drainage ditch Construction of the drainage ditch at LR3 transported solutes previously stored in the wetland downstream. Wetland water storage decreased exponentially when the drain was completed. Within four hours ~80% of solutes and 30% of water exported had exited the wetland. The newly constructed ditch acted primarily as a conduit, transporting solutes downstream directly from the wetland. Although some solute concentrations were significantly correlated with distance along the newly constructed ditch and had slopes that differed from the Cl slope, these relationships were not consistent across time. For example, concentrations of HCO3 were higher along the ditch length 4 hr and lower 6 hr since the start of the drainage experiment and concentrations of orthoP were often constant along the length of the ditch with the exception of the first point that differed. These data thus suggest that no consistent biotic or abiotic processing occurred along the length of the new ditch. This result is in direct contrast with previous studies that have shown that agricultural drainage ditches can act as solute sources 48 and/or sinks where changes in solute concentrations are attributed to sedimentation/resuspension, adsorption/desorption, biotic uptake/release, and microbial mediated reactions such as mineralization and nitrification (Skaggs et al., 1994; Sharpley et al., 2007; Strock et al., 2007). However, the conditions reported in this study differed from existing studies by: i) the experimental drainage ditch was shorter than ditches typically studied, such that the residence time may be too short for processing to occur; ii) the ditch was new and lacked well established aquatic vegetation and microbial communities; and iii) drainage occurred over a very short period in late fall when water temperature was near freezing. Cold temperatures have been shown by others to restrict nutrient uptake by vegetation, microbially mediated reactions, as well as sorption and diffusion rates (Kadlec and Reddy, 2001). The drainage conditions studied here are common in the prairies, and thus results given herein should be transferable to other watersheds in the PPR, although rigorous quantitative testing is recommended. Although there was no change in concentration along the length of the new ditch, there was an increase in concentration of most solutes studied with time since the start of drainage. This trend suggests that a vertical concentration gradient existed in the wetland such that the water closest to the sediment and in the sediment porewater had the highest constituent concentrations. Barica (1974), Fisher and Reddy (2001), and Barker et al. (2010) have all found vertical concentration gradients and elevated concentrations in sediment pore water in similar marshes and shallow lakes. Fecal coliforms also tend to concentrate in sediment where they survive longer, potentially due to the greater organic matter present in the sediment than water column (Karim et al., 2004). A vertical gradient in concentration may also explain why the masses of TP, orthoP, NO3, NH4, E. coli and T. coli exported via the drainage ditch exceeded estimates of those in the wetland. Wetland water samples were obtained from the center of the wetland at half the water depth and thus the masses calculated from these samples would be underestimated if a vertical concentration gradient existed. Another explanation for the increase in concentration of water quality parameters is that many wetlands in the prairies have salt rings around their perimeter. Salts are likely concentrated by shallow groundwater fluxes at this transition zone between wetlands and their uplands. Constructing a ditch that traverses this ring may have also contributed to the elevated concentrations and mass exceedances observed at LR3. Further research is needed to determine the cause of the mass exceedances because accurately estimating the amount of solutes leaving the wetland is the most important factor for predicting downstream export and associated ecological consequences, given that the ditch acted as a simple conduit. Had it not been for the uncharacteristically high midsummer rainfall, constituent concentrations leaving the wetland would probably have been higher and constituent mass exported lower at the time of drainage. Proportions of nutrients transported by snowmelt runoff typically exceed exports during rainfall events given that snowmelt runoff typically dominates prairie hydrology (Timmons and Holt, 1977; Tiessen et al., 2010). Decreases in specific conductivity and by association ion concentrations, have been attributed to rainfall and decreased evaporation (Barica 1978; LaBaugh and Swanson, 2004). The increases in ion concentrations between October 22 and the time of drainage were likely a result of their exclusion from the overlying ice and their freezing-out into a reduced water volume (Barica, 1975; Schwartz et al., 1978; Lilbaek and Pomeroy, 2008). In comparison, low coliform densities at the start of the drainage experiment were likely the result of sedimentation on the wetland bed (Auer and Niehaus, 1993; Wang and Doyle, 1998). 49 6.4 Comparing artificial ditches and natural spills Similar to the results for the newly constructed drainage ditch, water quality was not altered during transport along ditch or spill connections. Consequently, the significantly greater concentrations of TDN, DOC, HCO3, K, and Ca observed in ditches than spills were most likely due to differences in wetland water quality rather than differing physical attributes of drains and spills. Ditching permanent and semipermanent wetlands effectively turns them into seasonal ones because they are drained each year. Further, drained wetlands are predominantly located in cropped areas, a situation typical across the PPR (Gunterspergen et al., 2002). In contrast, spills flowed predominantly from permanent wetlands and were located only in grassland and wooded areas. As outlined in section 4.1 of this report, seasonal wetlands are characterized by greater concentrations of TDN and DOC, and K concentrations are greater in cropped wetlands than grassland or wooded wetlands. The ditches and spills also had significantly different physical characteristics. The ditches were more channelized, longer, and had higher flow velocities. Although this had no apparent direct influence on the quality of water moving along them, the differences are likely to influence the impacts on downstream water bodies. Wetland drainage ditches are created to connect wetlands to the watershed drainage network. In contrast, the short length of spills means they often are transiently connecting wetlands to other wetlands. These different connection characteristics suggest that wetland drainage has a higher likelihood of potentially enhancing downstream nutrient, salt and bacteria loading than spills. To date, the only study of prairie pothole drainage effects on downstream water quality (nutrients only) was a modeling exercise that compared different scenarios of wetland restoration at Broughton Creek watershed, Manitoba (Yang et al. 2008). Using the SWAT model, the researchers ran a wetland restoration scenario in which the 2005 wetland area was increased to match 1968 conditions. They predicted a 23% reduction in TN and TP loads to the stream using an empirical nutrient export coefficient. In contrast, results presented herein suggest that a nutrient export coefficient other than 1 is not warranted. This is likely the result of generally low temperatures during the snowmelt period when ditches and spills flow and when new ditch construction occurs. Fall ditch construction following harvest is more common than spring ditch construction because wetter soils in spring impede access to wetlands by heavy machinery and farmers are not preoccupied with seeding. Instead, the volume of water and mass of constituents exported from wetland drainage will depend on: i) how effectively the ditch drains the wetland; and ii) the water quality characteristics of the wetland that is influences by the combined permanence and land use setting. 6.5 Stream water quality There was a trend toward higher discharge for the subbasins with medium high and high wetland drainage compared to the one with low wetland drainage. Higher discharge is comparable to other studies that show wetland drainage increases effective contributing area and stream discharge (Campbell and Johnson, 1975; Saskatchewan Watershed Authority, 2008; Yang et al., 2008). Peak flows may have been delayed in the subbasin with low wetland drainage because of increased water storage potential (Pomeroy et al., 2009; Fang et al., 2010). The dominant anion switch from HCO3 to SO4 on the falling limb of the freshet hydrograph coincident with an increase in SC to >520 μS/cm was apparent in all subbasins. Therefore, it was not associated with wetland drainage. Instead, it likely occurred as proportion 50 of input from dilute snowmelt runoff decreased relative to that of saltier groundwater and more permanent surface water bodies. Subbasins characterized by moderately high and high wetland drainage had significantly higher mean export coefficients of TP, orthoP, HCO3, SO4, Mg, and Ca than the low drainage subbasin. These nutrients and salts have been shown in this report and by other researchers (Waiser and Robarts, 2004) to be concentrated in isolated prairie potholes located in cropped watersheds. The potholes most frequently drained in Smith Creek watershed are semi permanent to permanent ones located in cropped areas. The differences in water quality found among subbasins may have been partly attributed to variations in the land use distribution. Effects of nonpoint source pollution on stream water quality, particularly its eutrophic effect on downstream water bodies, have been widely documented; see Carpenter et al. (1998) for a comprehensive review. However, it is more likely that the differing mean export coefficients of TP, orthoP, HCO3, SO4, Mg, and Ca among subbasins was related to wetland drainage degree rather than land cover variations. Guo et al. (in press) report land covers in Smith Creek watershed as extracted from 2007 and 2008 classified SPOT5 images. Their data show land covers were very similar among subbasins; albeit they lumped subbasins SC4 and SC5 rather than reporting them separately (see Figure 4.3 for areas covered by these basins). Generally, no differences in the water quality parameters studied were found for subbasins characterized by medium high and high wetland drainage. This is not surprising given the relatively small difference (7%) in the proportion of wetlands lost for the two subbasins. Unfortunately, site selection preceded wetland loss determination from remote sensing imagery. It is recommended that future studies examine water quality in relation to stream flows across a wider range of wetland drainage scenarios. Some subbasins of Smith Creek watershed have much higher wetland loss than those studied here; for example, SC3 experienced ~81% wetland loss between 1958 and 2007-2008. Total export of the various water quality constituents at Smith Creek outlet, which is upstream of the town drainage (Langenburg Creek), was higher than the sum of the three studied subbasins. This suggests the lower part of the watershed, where many wetlands have been lost, is a significant source of nutrients, salts and bacteria to Smith Creek. Future examination of water quality in the subbasins that make up the lower part of the Smith Creek watershed is thus warranted. It has been shown in other agricultural systems outside of the PPR that wetland restoration reduces downstream nutrient loading (Woltemade, 2000). Water quality improvements are not necessary linearly related to numbers or areas of wetlands gained as the cumulative effects of wetland loss on downstream water quality are not usually additive and instead tend to be nonlinear (Whigham et al., 1988). Stream flows are also important in determining how effective wetlands are in maintaining good quality water downstream. For example, Johnston et al. (1990) reported that wetlands removed more TP, ammonia and suspended solids at high flows but removed more NO3 at lows. It would be very interesting to compare water quality in Smith Creek under different wetland restoration scenarios. 6.6 Exceedance of Federal and Provincial water quality guidelines for wetlands, ditches, spills and streams TP concentrations in the newly constructed ditch, 6 of 7 ditches, and 3 of 5 spills exceeded the Saskatchewan Watershed Authority (2007b) objective of 0.1 mg/L for the Lake Stewardship Program water quality index. This objective was also exceeded in 40 of 67 wetlands 51 sampled. Exceedances occurred in 71%, 50%, and 59% of crop, grass, and wood wetlands, and 77%, 65%, and 40% of seasonal, semi-permanent, and permanent wetlands. The TP objective was also exceeded for 45% of the LR3 wetland sampling instances. The TP objective was exceeded for all samples from the streams draining the subbasins with medium high and high wetland drainage and 63% of stream samples from the subbasin with low wetland drainage. The majority of NO3 concentrations measured in the wetlands, newly constructed ditch, ditches and spills, and streams did not exceed the Canadian Council of Ministers of the Environment (CCME) Canadian Environmental Quality guideline for the protection of aquatic life (2.9 mg/L; Canadian Council of Ministers of the Environment, 2003). However, this guideline was exceeded twice in the LR3 wetland following snowmelt, in 2 of 7 ditches, and in 1 stream sample from the high wetland drainage subbasin. The CCME guideline for NH3 + NH4 is greatly affected by temperature and pH; for typical conditions the guideline ranges from 18.5 mg/L (0oC, pH = 7.0) to 0.07 mg/L (15oC, pH = 9.0) (Canadian Council of Ministers of the Environment, 2003). Concentrations of NH4 did not exceed these guidelines, with the exception of the first two samples obtained following snowmelt in the LR3 wetland. The use of water containing salt concentrations that exceed CCME guidelines for water used for irrigation (100 – 700 mg Cl/L) and for livestock (1000 mg SO4/L and 1000 mg Ca/L) is not recommended. Concentrations of Cl, SO4, and Ca measured in the 67 wetlands, ditches and spills, and streams sampled in 2009 were below the CCME guidelines; thus drainage water would not be anticipated to degrade water quality with regards to these parameters. The guideline for SO4 was exceeded twice in the LR3 wetland when the wetland volume was lowest and in the newly constructed drainage ditch 1 hour after the start of drainage. The CCME indicator bacteria guideline for recreational water quality (200 E. coli per 100 ml) was only exceeded by the samples collected in the newly constructed ditch 23 hours after the start of drainage. The CCME guideline for the protection of agricultural water for crop irrigation (100 E. coli per 100 ml) was exceeded July 23, 2008 in the LR3 wetland. With the exception of the November 20, 2008 sample in the LR3 wetland, all samples collected in the LR3 wetland and the new ditch exceeded the animal-specific CCME guideline for livestock watering (2 E. coli per 100 ml of sample). E. coli densities exceeded the livestock watering guideline for 29%, 27% and 17% of the stream samples taken from subbasins with low, medium high, and high wetland drainage, respectively 6.7 Stream ecosystem health 6.7.1 Effects of wetland drainage stress levels on receiving lotic ecosystem health Data from 2008 and 2009 show that wetland drainage stress does not have a significant effect on the ecosystem health of the Smith Creek. However, the Langenburg Creek East of Langenburg is significantly impaired relative to reference, as two of the three samples from this site reveal significantly different assemblages (through CA Axis 1, Table 4.3) and one with significantly impacted diversity. This impairment may be the result of stressors from the town of Langenburg rather than wetland drainage as the samples are dominated by taxa such as Nematoda, Ostracoda, Oligochaeta, and Chironomidae, which typically reflect high organic pollution (Barbour et al., 1999). However, these results should merely be taken as an alarm that 52 warrants further study into the how drainage from the town of Langenburg may influence the ecosystem health of Smith Creek. The ditches included in this study did support aquatic benthic macroinvertebrate assemblages, and in the instance of ditch DT 3 it even had healthy assemblage. The other three ditches included in this study revealed stressed ecosystem health, particularly due to the richness and diversity measures of their condition. The assemblages in these three stressed ditches were dominated by the midges Chironomidae, the freshwater worms Oligochaeta and the biting midge (or no-see-ums) Ceratopogonidae with few other species. These are all organisms tolerant to environmental extremes and capable of multiple generations per year. Further, as the ditches dry these organisms either die and leave resting eggs, burrow deep into wet areas, or emerge and disperse as in the case of Chironomidae and Ceratopogonidae. 6.7.2 Findings on the influence of time on ecosystem health Reassuringly, sampling efforts from 2008 whereby multiple samples from Smith Creek North of Marchwell, Smith Creek East of Langenburg, and Langenburg Creek East of Langenburg were taken all provided relatively consistent precision in conclusions about the ecological condition at each site. As such, we are confident that single samples from each site in this watershed are relatively accurate in estimating condition, and that am certain that comparisons can be made through time both between years and between sampling periods within a year. In particular, the site Smith Creek East of Langenburg from 2008 and SC 3 from 2009 are overlapping sites and can be compared between the two years of study. The condition of this site in the river did not change between years despite a few samples appearing “stressed” in 2008; closer inspection shows that they are stressed because their diversity is higher than reference thus can be interpreted as a positive measure of impact. Therefore, this mainstem Smith Creek site did not vary in its ecosystem health between years. The second temporal component of this study, comparing between collection periods in the spring in 2009, produced results suggesting that ecosystem health may degrade as the stream slows and dries. Specifically, our two most northerly sites, and thus lower order and ergo earlier to cease flowing in the spring, dropped from a healthy assemblage of macroinvertebrates in the early spring to impaired at SC 4 and stressed at SC5. Interestingly, SC 4 has medium high drainage, while SC5 has low drainage. A potential explanation for the difference in impact between these two sites is that although they are lower order, the medium high drainage of the SC 4 site may have lead to a more flashy flow this spring while the low drainage of SC5 may have provided for a more prolonged and sustained flow into the later spring. The two remaining stream sites, TV1 and SC 3 are further downstream in the watershed and may have still had adequate flows to support this healthy macroinvertebrate assemblage through into mid May. Further investigation into the relationship between the flow in this watershed through time and the timing of ecosystem health degradation is warranted based on these preliminary observations. 6.7.3 Unanswered aspects of drainage-biotic relationships and future directions The lack of baseline data makes it difficult to place sites such as this in perspective, and such information would allow the monitoring of ecosystem health through time as drainage practices in the watershed are either increased or decreased. Further, lacking is any information on the effect drainage may have had on the wetland ecosystem health in the Smith Creek watershed. Certainly completely draining a wetland and ploughing it into terrestrial agricultural 53 land will result in a complete loss of aquatic ecosystem health, but intermediate drainage to control the expansion or merely to reduce the size of wetlands may have negative effects on the benthic macroinvertebrates due to loss of habitat complexity, and loss of habitat quantity. Developing a reference condition approach for wetlands would be extremely useful in setting ecosystem health objectives for wetlands, and assessing the impacts of activities such as drainage and land-use. 7 Conclusions and recommendations 7.1 Conclusions Project results show that prairie wetlands act as traps for nutrients, ions and bacteria. Temporally intensive measures of ions and DOC in one, permanent wetland suggest that hydrology is a strongly regulator of their storage. In contrast, nutrient and bacteria storage in the wetland appeared to be regulated by hydrological and biotic processes. Spatial variations in wetland water quality can be attributed in part to different land cover and permanence classes. Unexpectedly, there was no interactive effect of land cover and permanence classes on wetland solute chemistry. Also demonstrated was that neither SC nor ion dominance can be used to distinguish among wetland permanence classes at Smith Creek watershed. This lack of association is in contrast with previous studies that have linked ion dominance patterns and SC (as a proxy for net groundwater seepage rates) to wetland permanence. Overall, the results mean knowledge of land cover and/or permanence class can be used to provide a reasonable estimate of the water quality of a wetland. Wetland water quality was found to be an important control of water quality in drainage water. Thus, the occurrence of high nutrient concentrations (which were sometimes above federal or provincial water quality guidelines) measured in the LR3 wetland at the onset of the spring freshet has important implications because drained wetlands typically connect with streams or other downstream water bodies at this time. Hence, wetland drainage may augment downstream nutrient loads. Not assessed in the wetland drainage experiment was how changing the permanent wetland to a temporary one will influence nutrient exports in future spring freshet events. For instance, will nutrient concentrations remain high in the wetland from year to year, and similar to those observed in seasonal wetlands at Smith Creek? Or, will nutrients become progressively flushed from the stockpiles established in the drained wetland soils leading to reduced nutrient exports over time? Results also suggest that the efficiency with which a wetland is drained is an important factor in quantifying downstream exports. The temporally intensive measures of drainage water quality suggest that a vertical concentration gradient existed in the LR3 wetland such that the water closest to the sediment and in the sediment porewater of the wetland had the highest constituent concentrations. Thus wetland water samples obtained from the center of the wetland at half the water depth are likely to provide an underestimate of the total solute mass stored in the wetland if calculated from these samples. Ditches had higher TDN, DOC, HCO3, K, and Ca than spills. Similar to the results for the newly constructed drainage ditch, water quality was not altered during transport along ditch or spill connections. Since water quality was not altered during transport along the length of ditch or spill connections, differences in wetland water quality likely controlled ditch chemistry rather than differing physical attributes of drains. Drained wetlands are predominantly located in 54 cropped areas, a situation typical across the PPR, whereas natural connections among wetlands tended to be located in the grassland and wooded areas of Smith Creek watershed. Nearly all water quality parameters studied were higher in streams draining subbasins with greater wetland drainage. The provincial TP objective for protection of The low ratios of DIN to DIP that indicate Smith Creek is usually N-limited at its outlet combined with the finding that ditches export greater TDN than natural spills suggests that even small future increases in wetland drainage in the watershed could lead to enhanced stream algal growth. As Smith Creek is a sub-basin of the Assiniboine River, excess nutrient loadings from draining wetlands could facilitate further eutrophication of Lake Winnipeg, into which the Assiniboine River drains. It is important to remember that wetland drainage is not the only landscape stressor in watersheds located in the Prairie Pothole Region that can impact downstream water quality. This is the first to identify wetland drainage impacts on the instream ecosystem health. Overall, these preliminary results indicate that wetland drainage does not have a dramatic impact on the benthic macroinvertebrate assemblage. Due to the small number of sampling sites, results of this work are best used to inform future hypotheses about the interaction between wetland drainage and instream ecosystem health rather than being treated as a definitive conclusion with which to base management of wetland drainage. Aspects of the hydrology such as reduced permanence of wetlands produced by drainage could impact stream ecosystem health negatively as flashier, less stable freshets may result in quicker drying and less diverse assemblages of macroinvertebrates. Research is needed on this topic. Also lacking is any information about how changes in the water level of the drained wetlands may have altered their benthic macroinvertebrate assemblage or ecosystem health, and comparisons need to be made between the positive gains in ecosystem health downstream receiving waters get due to drainage compared to what may be lost from the wetlands being drained. It is hypothesized that the increase in amount of water in a lotic site may be the cause of increased ecosystem health. Conversely, decreasing water in a wetland via drainage could lead to declining wetland ecosystem health if the benthic macroinvertebrates respond similarly (but inversely) in streams and wetlands. 7.2 Recommendations Recommendation 1: Water quality along wetland drains was found to be similar to that in natural spills. Exceptions are that TDN, DOC, HCO3, K, and Ca concentrations were found to be higher along wetland drains. The important difference, however, is that spills typically connect wetlands to one another whereas drains connect wetlands to streams. One option to consider that could reduce downstream export of solutes and bacteria during wetland drainage is having wetland drains emulate spills. Drains could be constructed in such a manner as to connect wetlands to other wetlands so that solutes and bacteria remain stored in the watershed. It would be ideal to consider locating these storage zones in areas of the watershed where land productivity is low. Recommendation 2: Little change in water quality along the length of wetland drains was found. This means that the use of an empirical nutrient export coefficient is not required to be included in a model suited to simulating spring freshet nutrient exports due to wetland drainage at Smith Creek. However, knowing wetland water quality and how effectively the ditch drains the wetland are important to 55 predicting solute exports to streams. Future models should thus include representations of wetland land use and permanence setting, and wetland drainage efficacy. Recommendation 3: Many wetland drains connect directly to roadside ditches that then empty into Smith Creek. An evaluation of solutes losses or gains during transport along roadside ditches was not investigated and future study is warranted. Recommendation 4: At Smith Creek, wetland drainage appears to strongly influence stream water quality, but not stream biotic integrity. This conclusion has strong policy implications but has been determined for only a small number of sites in one watershed in the prairies over a limited time period. Basins in other parts of the prairies with differing wetland and drainage configurations, soils and climate should be investigated to see if this conclusion holds elsewhere or is specific to Smith Creek. Recommendation 5: The wetland drainage experiment performed showed that considerable solute and bacteria exports occur during drainage of a permanent wetland. Needed is an examination of solute release upon re-wetting of the wetland bed and subsequent export along drains for wetlands that have experienced repeated draining. Recommendation 6: This study did not examine benthic macroinvertebrate assemblages in wetlands being drained. As drainage reduces the permanence of a wetland, investigation into linkages between wetland water availability and biotic health is needed. 8 Acknowledgements We are grateful to the Saskatchewan Ministry of Agriculture for funding this project. Additional graduate student support was received from an NSERC PGS-M. We thank Larisa Barber, Deanne A. Schulz, Kevin Kirkham, Nicole Seitz, Logan Fang, Mike Solohub, and Adam Minke for their help in the field. Two Smith Creek watershed residents, Barry Schultz and Don Werle were instrumental in providing the logistical support needed to complete the field work. Sorting assistance for the macroinvertebrate samples was provided by Janet Halpin. We thank Adam Minke for producing several of the GIS maps and computing water storage in the LR3 wetland. Various archived GIS data were provided by Ducks Unlimited Canada, hydrometric data for Smith Creek gauging station were provided by Water Survey of Canada, and most taxonomic identifications were provided by AquaTax Consulting, Saskatoon. Discussions with John Pomeroy and Kevin Shook were very important to the success of this project. This work would not have been possible without the support and cooperation of the land owners and producers of Smith Creek watershed, the Smith Creek Advisory Committee and the Langenburg Regional Economic Development Authority. 56 9 References Agriculture and Agri-Food Canada (2009) SKSIDv4: Seamless digital soil resource information for Saskatchewan. Saskatchewan Land Resource Unit. Aldous, A., McCormick, P. Ferguson, C. Graham, S., Craft, C. (2005) Hydrologic regime controls soil phosphorus fluxes in restoration and undisturbed wetlands. Society for Ecological Restoration International 13(2):341-347. Anderson, D.W. (1988) The effect of parent material and soil development on nutrient cycling in temperate ecosystems. Biogeochemistry 5:71-97 Andres, D.D., Van Der Vinne, G. (2001) Calibration of ice growth models for bare and snow covered conditions: A summary of experimental data from a small prairie pond. 11th workshop on river ice: River ice processes within a changing environment. CGU-HS Committee on River Ice Processes and the Environment (CRIPE). Armstrong, N. (2002) Assiniboine river water quality study: Nitrogen and phosphorous dynamics May 2001 to May 2002. Manitoba Conservation Report No. 2002-10. Arndt, J.L., Richardson, J.L. (1993) Temporal variations in the salinity of shallow groundwater from the periphery of some North Dakota wetlands (USA). Journal of Hydrology 141:75105. Arts, M.T., Robarts, R.D., Kasai, F., Waiser, M.J., Tumber, V.P., Plante, A.J., Rai, J., de Lange, H.J. (2000) The attenuation of ultraviolet radiation in high dissolved organic carbon waters of wetlands and lakes on the northern Great Plains. Limnology and Oceanography 45(2):292-299. Auer, M.T., Niehaus, S.L. (1993) Modeling fecal coliform bacteria - I. Field and Laboratory determination of loss kinetics. Water Research 27(4):693-701. Baldwin, D.S., Mitchell, A.M. (2000) The effects of drying and re-flooding on the sediment and soil nutrient dynamics of lowland river-floodplain systems: A synthesis. Regulated Rivers: Research & Management 16:457-467. Barbour, M.T., Gerritsen, J., Snyder, B.D., Stribling, J.B. (1999) Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, benthic macroinvertebrates, and fish. EPA 841-0B-99-002. Office of Water, US Environmental Protection Agency, Washington, D.C., USA. Barica, J. (1974) Some observations on internal recycling, regeneration and oscillation of dissolved nitrogen and phosphorus in shallow self-contained lakes. Archiv für Hydrobiologie 73(3):334-360. Barica, J. (1975) Geochemistry and nutrient regime of saline eutrophic lakes in the EricksonElphinstone district of Southwestern Manitoba. Department of the Environment Fisheries and Marine Services Research and Development Directorate. Technical Report No. 511. Freshwater Institute, Winnipeg, Manitoba. Barica, J. (1978) Variability in ionic composition and phytoplankton biomass of saline eutrophic prairie lakes within a small geographic area. Archiv für Hydrobiologie 81(3):304-326. Barica, J. (1990) Seasonal variability of N:P ratios in eutrophic lakes. Hydrobiologia 191:97-103. Bärlocher, F., Mackay, R.J., Wiggins, G.B. (1978) Detritus processing in a temporary vernal pool in southern Ontario. Archiv für Hydrobiologie 81:269-295. Barker, T., Irfanullah, H. MD., Moss, B. (2010) Micro-scale structure in the chemistry and biology of a shallow lake. Freshwater Biology 55:1145-1163. 57 Batt, B.D.J., Anderson, M.G., Anderson, C.D., Caswell, F.D. (1989) The use of prairie potholes by North American ducks. In: van der Valk, A.G. (Ed.) Northern prairie wetlands, Iowa State University Press, Ames, Iowa, USA. Birgand, F., Skaggs, R.W., Chescheir, G.M., Gilliam, J.W. (2007) Nitrogen removal in streams of agricultural catchments - A literature review. Critical Reviews in Environmental Science and Technology 37:381-487. Bodhinayake, W., Si, B.C. (2004) Near saturated surface soil hydraulic properties under different land uses in the St Denis National Wildlife Area, Saskatchewan, Canada. Hydrological Processes 18:2835-2850. Bowman, M.F., Somers, K.M. (2006) Evaluating a novel Test Site Analysis (TSA) bioassessment approach. Journal of the North American Benthological Society 25(3):712-727. Brinson, M.M., Lugo, A. E., Brown, S. (1981) Primary productivity, decomposition and consumer activity in freshwater wetlands. Annual Review of Ecology and Systematics 12:123-161. Brinson, M.M., Malvarez, A.I. (2002) Temperate freshwater wetlands: types, status, and threats. Environmental Conservation 29:115-113. Brooks, A.R., Kelton, L.A. (1967) Aquatic and semiaquatic Heteroptera of Alberta, Saskatchewan, and Manitoba (Hemiptera). Memoirs of the Entomological Society of Canada 51: 3-92. Campbell, K.L., Johnson, H.P. (1975) Hydrologic simulation of watersheds with artificial drainage. Water Resources Research 11:120–126. Canadian Council of Ministers of the Environment, CCME (2003) Canadian water quality guidelines. Prepared by the Task Force on Water Quality Guidelines. CAPMoN (2007) Canadian Air and Precipitation Monitoring Network (CAPMoN) Precipitation Chemistry. http://www.msc.ec.gc.ca/capmon/index_e.cfm. Carpenter, S.R., Caraco, N.F., Correll, D.L., Howarth, R.W., Sharpley, A.N., Smith V.H. (1998) Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecological Applications 8:559-568. Clarke, K.R., Warwick, R.M. (2001) Change in Marine Communities: An approach to statistical analysis and interpretation, 1st edition. Plymouth Marine Laboratory, Plymouth, UK, 144pp. Clifford, H.F. (1991) The aquatic macroinvertebrates of Alberta. University of Alberta Press, Edmonton, Alberta. Cowardin, L.M., Gilmer, D.S., Mechlin, L.M. (1981) Characteristics of Central North Dakota wetlands determined from sample aerial photographs and ground study. Wildlife Society Bulletin 9(4):280-288. Crosbie, B., Chow-Fraser, P. (1999) Percentage land use in the watershed determines the water and sediment quality of 22 marshes in the great lakes basin. Canadian Journal of Fisheries and Aquatic Sciences, 56:1781-1791. Crumpton, W.G., Goldsborough, L.G. (1998) Nitrogen transformation and fate in prairie wetlands. Great Plains Research 8(1):57-72. Dahl, T.E. (1990) Wetlands-losses in the United States, 1780's to 1980's. Washington, D.C., U.S. Fish and Wildlife Service Report to Congress. 58 Davis, C.B., van der Valk, A.G. (1978) Litter decomposition in prairie glacial marshes. pp. 99113. In: Good, R.E., Whigham, D.F., Simpson, R.L. (Eds.) Freshwater Wetlands, Academic Press, New York. Detenbeck, N.E., Elonen, C.M., Taylor, D.L., Cotter, A.M., Puglisi, F.A., Sanville, W.D. (2002) Effects of agricultural activities and best management practices on water quality of seasonal prairie pothole wetlands. Wetland Ecology and Management 10:335-354. Devito, K.J., Creed, I.F., Rothwell, R.L., Prepas, E.E. (2000) Landscape controls on phosphorus loading to boreal lakes: implications for the potential impacts of forest harvesting. Canadian Journal of Fisheries and Aquatic Sciences 57:1977-1984. Dosdall, L., Lemkuhl, D.M. (1979) Stoneflies (Plecoptera) of Saskatchewan. Quaestiones Entomologicae 15(1):3-116. Driver, E.A., Peden, D.G. (1977) Chemistry of surface-water in prairie ponds. Hydrobiologia 53(1):33-48. Duff, J.H., Carpenter, K.D., Snyder, D.T., Lee, K.K., Avazino, R.J., Triska, F.J. (2009) Phosphorus and nitrogen legacy in restoration wetland, Upper Klamath Lake, Oregon. Wetlands 29(2):735-746. Environment Canada (2006) Archived hydrometric data: Smith Creek near Marchwell (05ME007) http://www.wsc.ec.gc.ca/. Water Survey of Canada, Canada. Euliss, N.H. Jr., Mushet, D.M., Johnson, D.H. (2001) Use of macroinvertebrates to identify cultivated wetlands in the prairie pothole region. Wetlands 21:223–231. Euliss, N.H., Jr., Mushet, D.M., Wrubleski, D.A. (1999) Wetlands of the Prairie Pothole Region: Invertebrate species composition, ecology, and management. In: Batzer, D.P., Rader R.B., Wissinger, S.A. (Eds.) Invertebrates in freshwater wetlands of North America: Ecology and management. John Wiley & Sons, New York. Jamestown, pp. 471-514. http://www.npwrc.usgs.gov/resource/wetlands/pothole/index.htm (Version 02SEP99). Everitt, B.S., Hothorn, T. (2006) A handbook of statistical analyses using R. Chapman & Hall, Boca Raton, Florida. Fang, X., Pomeroy, J.W., Westbrook, C.J., Guo, X., Minke, A.G., & Brown, T. (2010) Prediction of snowmelt derived streamflow in a wetland dominated prairie basin. Hydrology and Earth System Sciences 14:991-1006. Fisher, M.M., Reddy, K.R. (2001) Phosphorus flux from wetland soils affected by long-term nutrient loading. Journal of Environmental Quality 30:261-271. Fuller, L.G., Anderson, D.W. (1993) Changes in soil properties following forest invasion of Black soils of the Aspen Parkland. Canadian Journal of Soil Science 73:613-627. Gerla, P.J. (1992) The relationship of water-table changes to the capillary fringe, evapotranspiration, and precipitation in intermittent wetlands. Wetlands 12(2):91-98. Gray, D.M., Toth, B., Zhao, L., Pomeroy, J.W., Granger, R.J. (2001) Estimating areal snowmelt infiltration into frozen soils. Hydrological Processes 15:3095-3111. Guo, X., Pomeroy, J., Fang, X., Lowe, S., Li, Z., Westbrook, C., Minke, A. (in press). Effects of classification approaches on CHRM model performance. Remote Sensing Letters. Guntenspergen, G.R., Peterson, S.A., Leibowitz, S.G., Cowardin, L.M. (2002) Indicators of wetland condition for the prairie pothole region of the United States. Environmental Monitoring and Assessment 78:229-252. Hall, R.I., Leavitt, P.R., Quinlan, R., Dixit, A.S., Smol, J.P. (1999) Effects of agriculture, urbanization, and climate on water quality in the northern Great Plains. Limnology and Oceanography 44(3,2):739-756. 59 Hansen, N.C., Daniel, T.C., Sharpley, A.N., Lemunyon, J.L. (2002) The fate and transport of phosphorus in agricultural systems. Journal of Soil and Water Conservation 57(6):408417. Hayashi, M., van der Kamp, G. (2000) Simple equations to represent the volume-area-depth relations of shallow wetlands in small topographic depressions. Journal of Hydrology 237:74–85. Hayashi, M., van der Kamp, G., Rudolph, D.L. (1998a) Water and solute transfer between a prairie wetland and adjacent uplands, 1. Water balance. Journal of Hydrology 207:42-55. Hayashi, M., van der Kamp, G., Rudolph, D.L. (1998b) Water and solute transfer between a prairie wetland and adjacent uplands, 2. Chloride cycle. Journal of Hydrology 207:56-67. Heagle, D.J., Hayashi, M., van der Kamp, G. (2007) Use of solute mass balance to quantify geochemical processes in a prairie recharge wetland. Wetlands 27(4):806-818. Hemond, H.F., Benoit, R.J. (1988) Cumulative impacts on water quality functions of wetlands. Journal of Environmental Management 12:639-654. Hilsenhoff, W.L. (1988) Rapid field assessment of organic pollution with a family-level biotic index. Journal of North American Benthological Society 7(1):65-68. Holland, H.D. (1978) The chemical evolution of the atmosphere and oceans. Wiley, New York. Hyer, K.E., Moyer, D.L. (2004) Enhancing fecal coliform total maximum daily load models through bacterial source tracking. Journal of the American Water Resources Association 40(6):1511-1526. Hyland, R., Byrne, J., Selinger, B., Graham, T., Thomas, J., Townsend, I., Gannon, V. (2003) Spatial and temporal distribution of fecal indicator bacteria within the Oldman River basin of southern Alberta, Canada. Water Quality Research Journal Canada 38:15-32. Johnson, W.C., Boettcher, S.E., Poiani, K.A., Guntenspergen, G. (2004) Influence of weather extremes on the water levels of glaciated prairie wetlands. Wetlands 24(2): 385-398. Johnston, C.A., Detenbeck, N.E., Niemi, G.J. (2000) The cumulative effect of wetlands on stream water quality and quantity. A landscape approach. Biogeochemistry 10:105-141. Julien, P.Y. (2002) River mechanics. Cambridge University Press, New York. Kadlec, R.H., Pries, J., Mustard, H. (2007) Muskrats (Ondatra zibethicus) in treatment wetlands. Ecological Engineering 29:143-153. Kadlec, R.H., Reddy, K.R. (2001) Temperature effects in treatment wetlands. Water Environment Research 73(5):543-557. Karim, M.R., Manshad, F.D., Karpiscak, M.M., Gerba, C.P. (2004) The persistence and removal of enteric pathogens in constructed wetlands. Water Research 38:1831-1837. Kemp, M.J., Dodds, W.K. (2001) Centimeter-scale patterns in dissolved oxygen and nitrification rates in a prairie stream. Journal of the North American Benthological Society 20(3):347357. Köchy, M., Wilson, S.D. (1997) Litter decomposition and nitrogen dynamics in aspen forest and mixed-grass prairie. Ecology 78(3):732-739. LaBaugh, J.W., Swanson, G.A. (2004) Spatial and temporal variability in specific conductance and chemical characteristics of wetland water and in water column biota in the wetlands in the Cottonwood Lake Area. In Winter, T.C. (Ed.), Hydrological, chemical, and biological characteristics of a prairie pothole wetland complex under highly variable climate conditions – the Cottonwood Lake area, east-central North Dakota. United States Geological Survey Professional Paper 1675, Denver, CO. p. 35-54. 60 LaBaugh, J.W., Winter, T.C., Adomaitis, V.A., Swanson, G.A. (1987) Hydrology and chemistry of selected prairie wetlands in the Cottonwood Lake Area, Stutsma County, North Dakota. U.S. Geological Survey Professional Paper 1431. Larson, D.J., Alarie, Y., Roughley, R.E. (2000) Predaceous diving beetles (Coleoptera: Dytiscidae) of the neoarctic region, with emphasis on the fauna of Canada and Alaska. NRC Research Press, Ottawa, Canada. Legendre, P., Legendre, L. (1998) Numerical ecology. Second English edition. Elsevier Press, Amsterdam, The Netherlands. Leibowitz, S.G., Vining, K.C. (2003) Temporal connectivity in a prairie pothole complex. Wetlands 23(1):13-25. Lilbæk, G., Pomeroy, J.W. (2008) Ion enrichment of snowmelt runoff water caused by basal ice formation. Hydrological Processes 22:2758-2766. Little, J.L., Nolan, S.C., Casson, J.P., Olson, B.M. (2007) Relationship between soil and runoff phosphorus in small Alberta watersheds. Journal of Environmental Quality 36:12891300. McAllistor, L.S., Peniston, B., Leibowitz, S.G., Abbruzzese, B., Hyman, J.B. (2000) A synoptic assessment for prioritizing wetland restoration efforts to optimize flood attenuation. Wetlands 20(1):70-83. Merritt, R.W., Cummins, K.W. (1996) An introduction to the aquatic insects of North America, third edition. Kendall/Hunt Publishing Company, Dubuque, Iowa, USA. Meybeck, M. (2005) Looking for water quality. Hydrological Processes 19:331-338. Millar, J.B. (1976) Wetland classification in western Canada: a guide to marshes and shallow open water wetlands in the grasslands and parklands of the Prairie Provinces. Canadian Wildlife Service Report, Series No. 37, Ottawa. Miller, J.J., Acton, D.F. St. Arnaud, R.J. (1985) The effect of groundwater in soil formation in a morainal l&scape in Saskatchewan. Canadian Journal of Soil Science 65:293-307. Millet, B., Johnson, W.C., Guntenspergen, G. (2009) Climate trends of the North American prairie pothole region 1906-2000. Climactic Change 93:243-267. Minke, A.G., Westbrook, C.J., van der Kamp, G. (2010) Simplified volume-area-depth method for estimating water storage of prairie potholes. Wetlands 30(3):541-551. Murkin, H.A. (1998) Freshwater functions and values of prairie wetlands. Great Plains Research 8(1):3-15. National Wetlands Working Group, NWWG (1988). Wetlands of Canada. Ecological Land Classification Series, no. 24, Environment Canada, Ottawa, Ontario and Polysicience Publications, Inc, Montreal, Quebec, 452p. Needelman, B.A., Ruppert, D.E., Vaughan, R.E. (2007) The role of ditch soil formation and redox biogeochemistry in mitigating nutrient and pollutant losses from agriculture. Journal of Soil and Water Conservation, 62:207-215. Neely, R.K., Baker, J.L. (1989) Nitrogen and phosphorous dynamics and fate of agricultural runoff. In: van der Valk, A.G. (Ed.) Northern Prairie Wetlands. Iowa State University Press, Ames, Iowa. Neill, C. (1995) Seasonal flooding, nitrogen mineralization and nitrogen utilization in a prairie marsh. Biogeochemistry 30:171-189. Nguyen, L., Sukias, J. (2002) Phosphorous fraction and retention in drainage ditch sediments receiving surface runoff and subsurface drainage from agricultural catchments in New Zealand. Agriculture Ecosystems and Environment 92:49-69. 61 Nicholson, B.J. (1995) The wetlands of Elk Island National Park: Vegetation classification, water chemistry, and hydrotopographic relationships. Wetlands 15(2):119-133. Ontkean, G.R., Chanasyk, D.S., Riemersma, S., Bennett, D.R., Brunen, J.M. (2003) Enhanced prairie wetland effects on surface water quality in Crowfoot Creek, Alberta. Water Quality Research Journal of Canada, 38(2): 335-359. Parsons, D.F., Hayashi, M., van der Kamp, G. (2004) Infiltration and solute transport under seasonal wetland: bromide tracer experiment in Saskatoon, Canada. Hydrological Processes 18:2011-2027. Phillips, I.D., Parker, D., McMaster, G. (2008) Aquatic invertebrate fauna of a northern prairie stream: range extensions and water quality characteristics. Western North American Naturalist 68(2):173-185. Phillips, I.D., McMaster, G. (2010) Feasibility of assessing Saskatchewan prairie stream health using macroinvertebrates. Science, Information and Monitoring, Saskatchewan Watershed Authority. Pomeroy, J.W., Fang, X., Westbrook, C.J., Minke, A., Guo, X., Brown, T. (2009) Prairie Hydrological Model Study: Final Report. Centre for Hydrology Report No. 7. Centre for Hydrology, University of Saskatchewan, Saskatoon. Pomeroy, J.W., Granger, R.J., Pietroniro, A., Elliott, J.E., Toth, B., Hedstrom, N. (1997) Hydrological pathways in the Prince Albert Model Forest. A final report submitted to: The Prince Albert Model Forest Association. National Hydrology Research Institute Environment Canada Saskatoon, Saskatchewan. Porter, R.M., van Kooten, G.C. (1993) Wetlands preservation on the Canadian Prairies: The problem of the public duck. Canadian Journal of Agricultural Economics 41(4):401-410. R Core Development Team (2005) R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. Rawson, D. (1944) The saline lakes of Saskatchewan. Canadian Journal of Research: Section D Zoological Sciences 22(6):141-201. Reddy, K.R., Patrick, W.H. Jr. (1975) Effect of alternate aerobic and anaerobic conditions on redox potential, organic matter decomposition and nitrogen loss in a flooded soil. Soil Biology and Biogeochemistry 7:87-94. Rhee, G-Y., Gotham, J. (1980) Optimum N:P rations and coexistence of planktonic algae. Journal of Phycology 16:486-489. Rózkowska, A.D., Rózkowski, A. (1969) Seasonal changes of slough and lake chemistry in southern Saskatchewan (Canada). Journal of Hydrology 7:1-13. Rózkowski, A. (1969) Chemistry of ground and surface waters in the Moose Mountain area, southern Saskatchewan. Geological Survey of Canada Paper No. 67-9. Saskatchewan Watershed Authority (2007a) State of the Watershed Report. Government of Saskatchewan State of the Environment Report. Saskatchewan Watershed Authority (2007b) Lake Stewardship Water Quality Guide. Monitoring and Assessment Branch Stewardship Division. Saskatchewan Watershed Authority (2008) Agricultural drainage impacts on Fishing and Waldsea lakes. Saskatchewan Watershed Authority (2010) State of the watershed report, http://www.swa.ca/Publications/Documents/d_AppendixA-ConditionIndicators.PDF Scarth, J. (1998) Wetland policy in Canada: A research agenda for policy reform. Great Plains Research 8:169-182. 62 Schwartz, F.W., Gallup, D.N. (1978) Some factors controlling the major ion chemistry of small lakes: examples from the Prairie Parkland of Canada. Hydrobiologia 58(1):65-81. Sharpley, A.N., Krogstad, T., Kleinman, P.J.A., Haggard, B., Shigaki, F., Saporito, L.S. (2007) Managing natural processes in drainage ditches for nonpoint source phosphorus control. Journal of Soil and Water Conservation 62(4):197-206. Shjeflo, J.B. (1968) Evapotranspiration and the water budget of prairie potholes in North Dakota. US Geological Survey Professional Paper 585-B. Skaggs, R.W., Brevé, M.A., Gilliam, J.W. (1994) Hydrology and water quality impacts of agricultural drainage. Critical Reviews in Environmental Science and Technology 24:132. Sloan, C.E. (1972) Ground-water hydrology of prairie potholes in North Dakota. US Geological Survey Professional Paper 585-C. Smith, A. G., Stoudt, J.H., Gollop, J.B. (1964) Prairie potholes and marshes. In: Linduska, J.P. (Ed.) Waterfowl tomorrow. US Fish and Wildlife Service, Washington, DC, pp. 39-50. Spence, C. (2006) On the relation between dynamic storage and runoff: A discussion on thresholds, efficiency, and function. Water Resources Research 43:1-11. Spence, C., Woo, M-K. (2003) Hydrology of subarctic Canadian Shield: soil-filled valleys. Journal of Hydrology 279(1-4):151-166. Stichling, W., Blackwell, S.R. (1957) Drainage area as a hydrologic factor on the glaciated Canadian prairies. International Association for Scientific Hydrology 45: 65-376. Stoddard, J.L., Larsen, D.P., Hawkins, C.P., Johnson, R.K., Norris, R.H. (2006) Setting expectations for the ecological condition of stream: the concept of reference condition. Ecological Applications 16:1267-1276. Strock, J.S., Dell, C.J., Schmidt, J.P. (2007) Managing natural processes in drainage ditches for non-point source nitrogen control. Journal of Soil and Water Conservation 62(4):188196. Su, M., Stolte, W.J., van der Kamp, G. (2000) Modelling Canadian prairie wetland hydrology using a semi-distributed streamflow model. Hydrological Processes 14:2405-2422. Swanson, G.A., Winter, T.C., Adomaitis, V.A., LaBaugh, J.W. (1988) Chemical characteristics of prairie lakes in south-central North Dakota USA - Their potential for influencing use by fish and wildlife. US Department of the Interior, Fish and Wildlife Service Technical Report 18:44. Tangen, B.A., Butler, M.G., Ell, M.J. (2003) Weak correspondence between macroinvertebrate assemblages and land use in prairie pothole region wetlands, USA. Wetlands 23:104-115. Tiessen, K.H.D., Elliott, J.A., Yarotski, J., Lobb, D.A., Flaten, D.N., Glozier, N.E. (2010) Conventional and conservation tillage: influence on seasonal runoff, sediment, and nutrient losses in the Canadian Prairies. Journal of Environmental Quality 39:964-980. Timmons, D.R., Holt, R.F. (1977) Nutrient losses in surface runoff from a native prairie. Journal of Environmental Quality 6(4):369-373. Tiner, R.W. (1984) Wetlands of the United States: Current status and recent trends. US Fish and Wildlife Service, Washington, D.C. 59 pp. Tiner, R.W. (2003) Geographically isolated wetlands of the United States. Wetlands 23(3):494516. Tóth J. (1963) A theoretical analysis of groundwater flow in a small drainage basin. Journal of Geophysical Research. 68(16):4795-4811. 63 van der Kamp, G., Hayashi, M. (2009) Groundwater-wetland ecosystem interaction in the semiarid glaciated plains of North America. Hydrogeology Journal 17(1):203-214. van der Kamp, G., Hayashi M., Gallen, D. (2003) Comparing the hydrology of grassed and cultivated catchments in the semi-arid Canadian prairies. Hydrological Processes 17:559575. van der Kamp, G., Keir, D., Evans, M.S. (2008) Long-term water level changes in closed-basin lakes of the Canadian prairies. Canadian Water Resources Journal 33:23-38. Waiser, M.J. (2006) Relationship between hydrological characteristics and dissolved organic carbon concentration and mass in northern prairie wetlands using a conservative tracer approach. Journal of Geophysical Research 111: G02024, doi:10.1029/2005JG000088. Waiser, M.J., Robarts, R.D. (2004) Net heterotrophy in productive prairie wetlands with high DOC concentrations. Aquatic Microbial Ecology 34:279-290. Wang, G., Doyle, M. (1998) Survival of Enterohemorrhagic Escherichia coli O157:H7 in water. Journal of Food Protection 61(6):662-667. Wang, J.R., Zhong, A.L., Comeau, P., Tsze, M., Kimmins, J.P. (1995) Aboveground biomass and nutrient accumulation in an age sequence of aspen (Populus tremuloides) stands in the boreal white and black spruce zone, British Columbia. Forest Ecology and Management 78:127-138. Watmough, M.D., Schmoll, M.J. (2007) Environment Canada’s prairie and northern region habitat monitoring program phase II: Recent habitat trends in the prairie habitat joint venture. Technical Report Series Number 493, Ottawa: Environment Canada, Canadian Wildlife Service. Webb, J.M. (2002) The mayflies of Saskatchewan. M.Sc. thesis, University of Saskatchewan, Saskatoon, SK, Canada. Wetzel, R.G. (2001) Limnology: Lake and river ecosystems, 3rd edition. Academic Press, San Diego, California. Whigham, D.F., Chitterling, C., Palmer, B. (1988) Impacts of freshwater wetlands on water quality: a landscape perspective. Environmental Management 12(5):663-671. Whigham, D.F., Jordan, T.E. (2003) Isolated wetlands and water quality. Wetlands 23(3):541549. Winter, T.C., LaBaugh, J.W. (2003) Hydrological considerations in defining isolated wetlands. Wetlands 23(3):532-540. Winter, T.C., Rosenberry, D.O. (1995) The interaction of groundwater with prairie pothole wetlands in the Cottonwoods Lake area, east-central North Dakota, 1979-1990. Wetlands 15(3):193-211. Woltemade, C.J. (2000) Ability of restored wetlands to reduce nitrogen and phosphorus concentrations in agricultural drainage waters. Journal of Soil and Water Conservation 55:303-309. Woo, M-K., Roswell, R.D. (1993) Hydrology of a prairie slough. Journal of Hydrology 146:175207. Yang, W., Wang, X., Gabor, S., Boychuck, L., Badiou, P. (2008) Water quantity and quality benefits from wetland conservation and restoration in the Broughton's Creek watershed. Ducks Unlimited Canada publication. Zar, J.H. (2000) Biostatistical analysis, fourth edition. Prentice-Hall, Englewood Cliffs, New Jersey. 64 10 Other aspects 10.1 Communications The research group utilizes several methods to communicate our project and its findings with other researchers, the people living in the watersheds we study, and society. These include conference talks, academic publications, public open houses and website development (www.usask.ca/hydrology). Journal publications are forthcoming and we are currently preparing an open house for the residents of Smith Creek watershed which is expected to be held in early spring of 2011. Ms. Brunet will defend her thesis in March 2011. Below is a list of all scholarly communications of this project thus far. CBC Saskatchewan Radio and Ducks Unlimited Canada have asked for a copy of this report. Brunet, N. and Westbrook, C.J. 2010. Prairie stream water quality in sub-basins characterized by differing degrees of wetland drainage. American Geophysical Union 2010 Fall Meeting, San Francisco, USA, December 13-17. Brunet, N. and Westbrook, C.J. 2010. Prairie wetland drainage effects on water quality. Canadian Soil Science Society Annual Meeting, Saskatoon, Canada, June 20-24. Brunet, N. and Westbrook, C.J. 2010. Prairie wetland drainage effects on water quality. Joint Assembly of the Canadian Geophysical Union and Canadian Meteorological and Oceanographic Society, Ottawa, Canada, May 31 to June 4. Brunet, N., and Westbrook C.J. 2010. Characterization of spatial variations in prairie wetland water quality. CGU-HS Student Meeting, Edmonton, Canada, January 30. Brunet, N. and Westbrook, C.J. 2009. Characterization of spatial variations in prairie wetland water quality. Drought Research Initiative Conference, Saskatoon, Canada, November 18. Westbrook, C.J. Pomeroy, J., Fang, X., Guo, X., Minke, A., Brunet, N., Shook, K., Brown, T., 2009. Smith Creek and the importance of hydrometric data in modeling and water quality research. Water Survey of Canada, Saskatchewan Division Staff Workshop, December, Saskatoon, Saskatchewan. 10.2 Personnel involved Cherie J. Westbrook – Principle Investigator, Assistant Professor, University of Saskatchewan Iain Phillips – Aquatic Macroinvertebrate Ecologist, Saskatchewan Watershed Authority John-Mark Davies – Water Quality Scientist, Saskatchewan Watershed Authority Nathalie Brunet – MSc student, University of Saskatchewan Erin Shaw – Lab technician, University of Saskatchewan Summer Students - Larisa Barber, Deanne Schulz, Kevin Kirkham, Nicole Seitz 65 11 Appendices Appendix A: Nutrient concentrations (May 19-21, 2009) for 67 wetlands in Smith Creek watershed, SK. Notes indicate whether the upland area surrounding the wetland was tilled, used for grazing, and the type of crop harvested in 2008, as reported by land owners. ID Land Cover Notes Permanence Max Depth orthoP TP TDN NO3 NH4 DOC (cm) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) DIN:DIP -5.8 W3 Grass Ungrazed Seasonal 44 0.005 0.09 1.13 0.018 0.011 33.0 W4 Grass Ungrazed Semiperm 79 0.005 0.09 0.91 0.002 0.016 28.9 3.6 W6 Grass Ungrazed Semiperm 63 0.005 0.13 0.90 0.021 0.032 30.3 10.6 W10 Grass Ungrazed Seasonal 62 0.03 0.17 1.08 0.018 0.036 28.4 1.8 W11 Grass Ungrazed Permanent 102 0.01 0.12 1.39 0.013 0.038 36.5 5.1 W13 Grass Ungrazed Permanent 87 0.01 0.07 1.01 0.007 0.010 33.1 1.7 W17 Wood Ungrazed Permanent 54 0.14 0.24 1.17 0.045 0.038 33.1 0.6 W18 Wood Ungrazed Permanent 60 0.06 0.13 0.94 0.009 0.018 19.2 0.5 W21 Wood Ungrazed Seasonal 44 0.12 0.22 1.50 0.014 0.041 38.6 0.5 W22 Wood Ungrazed Permanent 58 0.18 0.29 1.35 0.006 0.041 32.6 0.3 W24 Wood Ungrazed Semiperm 56 0.35 0.43 1.09 0.013 0.014 33.9 0.1 W27 Wood Ungrazed Permanent 100 0.03 0.16 1.05 0.017 0.044 21.6 2.0 W28 Wood Ungrazed Permanent 96 0.01 0.11 1.12 0.008 0.014 27.8 2.2 W30 Wood Ungrazed Semiperm 59 0.47 0.53 1.30 0.007 0.017 35.0 0.1 W32 Wood Ungrazed Semiperm 52 0.02 0.22 0.99 0.002 0.018 24.2 1.0 W34 Crop Canola/Tilled Permanent 67 0.03 0.08 1.16 0.027 0.040 22.3 2.2 W35 Crop Canola/Tilled Permanent 76 0.1 0.18 1.29 0.035 0.025 26.2 0.6 W37 Crop Canola/Tilled Semiperm 78 0.03 0.17 1.31 0.025 0.014 30.9 1.3 W40 Crop Canola/Tilled Seasonal 34 0.005 0.19 1.49 0.002 0.014 37.1 3.2 4.0 W42 Crop Canola/Tilled Seasonal 48 0.005 0.55 0.88 0.006 0.014 26.8 W43 Crop Canola/Tilled Permanent 109 0.005 0.05 0.77 0.006 0.036 21.4 8.4 W45 Grass Grazed Semiperm 74 0.03 0.11 1.41 0.008 0.053 31.3 2.0 1.4 W46 Grass Grazed Permanent 98 0.03 0.08 1.10 0.008 0.035 26.3 W47 Grass Grazed Seasonal 36 0.04 0.18 1.13 0.023 0.012 30.9 0.9 W48 Wood Grazed Permanent 90 0.005 0.02 0.81 0.006 0.012 22.5 3.6 W50 Wood Grazed Seasonal 34 0.005 0.10 1.48 0.010 0.042 30.1 10.4 W54 Wood Grazed Semiperm 44 0.005 0.09 1.05 0.012 0.035 29.8 9.3 W55 Wood Grazed Permanent 70 0.005 0.02 0.88 0.007 0.013 24.0 3.9 W61 Wood Grazed Permanent 80 0.005 0.02 1.14 0.005 0.014 29.5 3.8 W67 Wood Grazed Semiperm 65 0.005 0.04 1.15 0.004 0.012 31.0 3.2 W68 Wood Grazed Seasonal 20 0.005 0.06 1.61 0.002 0.013 31.7 3.0 W69 Wood Ungrazed Seasonal 22 0.005 0.11 1.82 0.040 0.041 52.8 16.2 W71 Wood Ungrazed Semiperm 66 0.005 0.17 1.40 0.037 0.040 30.1 15.4 W72 Wood Ungrazed Seasonal 61 0.03 0.18 1.48 0.023 0.041 35.5 2.1 W73 Wood Ungrazed Semiperm 64 0.02 0.05 1.77 0.034 0.015 42.4 2.4 W75 Wood Ungrazed Seasonal 28 0.005 0.11 1.66 0.022 0.042 38.3 12.8 2.9 W85 Crop Wheat/Tilled Semiperm 62 0.01 0.19 1.23 0.005 0.024 30.2 W86 Crop Wheat/Tilled Permanent 102 0.03 0.09 1.86 0.015 0.014 51.5 1.0 W87 Crop Wheat/Tilled Seasonal 26 0.26 0.61 1.98 0.027 0.039 50.7 0.3 W88 Crop Wheat/Tilled Semiperm 70 0.3 0.93 0.95 0.222 0.029 24.2 0.8 W89 Crop Wheat Permanent 102 0.005 0.06 0.83 0.007 0.031 20.7 7.6 W90 Crop Wheat Semiperm 87 0.005 0.13 1.64 0.016 0.020 38.8 7.2 W91 Crop Wheat Seasonal 50 0.43 1.10 1.65 0.027 0.020 44.2 0.1 W93 Crop Canola Seasonal 74 0.02 0.11 1.34 0.036 0.002 33.0 1.9 W96 Grass Ungrazed Seasonal 44 0.005 0.10 1.74 0.022 0.013 44.9 7.0 W98 Grass Ungrazed Permanent 90 0.01 0.03 1.15 0.022 0.033 33.2 5.5 W100 Grass Ungrazed Seasonal 32 0.005 0.05 1.25 0.028 0.034 35.3 12.4 W101 Grass Ungrazed Permanent 66 0.005 0.03 0.87 0.004 0.034 27.5 7.6 W102 Grass Ungrazed Seasonal 22 0.03 0.87 2.49 0.007 0.025 55.3 1.1 66 Appendix A: continued ID W103 Land Cover Grass Notes Grazed Permanence Max Depth orthoP TP TDN NO3 NH4 DOC (cm) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) DIN:DIP -- Permanent 97 0.12 0.29 1.00 0.002 0.045 29.7 0.4 W104 Grass Grazed Seasonal 26 0.03 0.22 1.41 0.005 0.009 33.6 0.5 W106 Grass Grazed Permanent 70 0.14 0.37 1.26 0.014 0.012 31.6 0.2 W107 Grass Grazed Permanent 98 0.005 0.07 0.79 0.019 0.023 24.8 8.4 W108 Grass Grazed Seasonal 20 0.01 0.42 2.77 0.026 0.048 49.6 7.4 W110 Grass Grazed Semiperm 46 0.005 0.12 0.85 0.226 0.015 24.2 48.1 W111 Crop Canola Semiperm 69 0.2 0.34 1.70 0.028 0.002 39.7 0.1 W112 Grass Ungrazed Semiperm 31 0.005 0.10 1.09 0.011 0.035 35.5 9.2 W113 Crop Wheat Permanent 91 0.005 0.05 1.06 0.045 0.035 27.5 16.0 W114 Crop Wheat Semiperm 84 0.38 2.80 1.60 0.008 0.039 37.3 0.1 W115 Crop Wheat Seasonal 34 0.13 1.30 1.83 0.022 0.016 42.0 0.3 W116 Crop Canola Permanent 80 0.005 0.12 1.34 0.016 0.084 37.5 20.0 W117 Crop Wheat/Tilled Seasonal 38 0.05 0.53 2.17 0.009 0.018 45.2 0.5 W118 Crop Wheat/Tilled Permanent 87 0.005 0.09 1.34 0.024 0.036 26.5 12.0 W119 Wood Grazed Permanent 70 0.01 0.08 1.31 0.226 0.015 34.7 24.1 W120 Grass Grazed Semiperm 74 0.005 0.07 0.90 0.014 0.015 28.1 5.8 W121 Grass Grazed Seasonal 55 0.02 0.57 0.90 0.025 0.036 19.6 3.0 W122 Grass Grazed Semiperm 60 0.005 0.08 0.92 0.026 0.033 24.2 11.8 67 Appendix B: Salt concentrations (May 19-21, 2009) for 67 wetlands in Smith Creek watershed. ID Land Cover Permanence pH SC Cl HCO3 SO4 Na Mg Ca K (μS/cm) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) W3 Grass Seasonal 6.75 163 2.1 89.8 0.8 1.3 9.7 8.6 16.0 W4 Grass Semiperm 7.24 414 1.8 120.5 95.7 8.9 28.2 21.1 10.0 W6 Grass Semiperm 7.31 656 1.7 124.7 210.0 19.3 68.5 39.2 11.9 W10 Grass Seasonal 7.44 309 3.8 80.2 22.0 2.6 15.3 12.7 14.0 W11 Grass Permanent 7.68 695 1.8 127.8 168.4 21.0 74.2 35.6 16.0 W13 Grass Permanent 7.24 404 1.3 79.3 49.3 5.7 20.3 9.7 7.9 W17 Wood Permanent 6.92 158 2.0 63.0 0.6 0.3 6.2 13.8 20.3 W18 Wood Permanent 6.55 63 1.5 25.7 0.1 0.2 1.6 4.6 11.3 W21 Wood Seasonal 6.78 149 2.3 54.4 6.3 0.3 4.8 11.3 23.4 W22 Wood Permanent 6.86 131 1.9 62.8 0.5 0.2 4.8 10.0 18.9 W24 Wood Semiperm 6.77 127 1.4 63.7 0.1 0.2 3.8 11.0 19.4 W27 Wood Permanent 6.74 111 2.4 48.2 2.3 0.5 4.3 8.7 12.7 W28 Wood Permanent 6.76 150 2.0 70.0 7.8 0.6 6.7 11.3 13.4 W30 Wood Semiperm 6.83 127 1.7 64.7 0.1 0.2 3.8 9.8 21.5 W32 Wood Semiperm 6.84 57 0.0 18.3 0.1 0.1 1.2 4.9 4.6 W34 Crop Permanent 7.19 181 2.0 72.6 9.1 2.0 7.8 13.1 8.3 W35 Crop Permanent 7.19 211 4.9 69.1 18.3 1.9 7.7 14.1 18.1 W37 Crop Semiperm 7.26 296 3.3 95.3 40.0 3.8 10.0 16.3 9.6 W40 Crop Seasonal 7.38 287 4.6 98.4 6.0 5.4 11.8 19.3 18.7 W42 Crop Seasonal 6.80 147 9.5 56.2 1.0 2.3 4.8 11.5 13.4 W43 Crop Permanent 7.59 230 2.9 66.9 2.7 1.7 6.6 10.4 11.2 W45 Grass Semiperm 7.28 413 3.2 98.5 70.2 7.5 29.1 14.5 14.2 W46 Grass Permanent 7.47 314 5.9 77.4 50.7 6.7 22.5 13.9 13.5 W47 Grass Seasonal 7.10 414 12.4 103.4 52.1 6.9 25.4 22.3 12.8 W48 Wood Permanent 7.75 306 2.0 113.2 39.7 4.5 17.5 18.3 12.9 W50 Wood Seasonal 7.19 144 0.7 53.9 14.0 0.5 7.8 12.0 9.1 W54 Wood Semiperm 8.62 451 1.9 62.7 114.6 9.9 41.2 30.7 17.3 W55 Wood Permanent 8.42 374 0.7 66.7 44.3 2.4 14.3 19.6 7.3 W61 Wood Permanent 7.74 422 3.9 94.6 81.7 6.8 24.5 18.2 18.2 W67 Wood Semiperm 6.93 134 2.8 79.9 0.7 0.6 5.0 9.0 19.3 W68 Wood Seasonal 6.88 112 1.2 55.5 1.3 0.4 4.7 10.8 12.6 W69 Wood Seasonal 7.06 211 1.4 87.9 8.1 0.9 7.6 13.3 34.1 W71 Wood Semiperm 7.09 182 2.0 80.4 3.9 0.8 5.9 11.3 16.7 W72 Wood Seasonal 6.90 170 2.7 77.7 5.4 0.9 6.9 11.0 25.8 W73 Wood Semiperm 6.94 196 1.7 110.9 9.2 1.5 10.3 12.7 22.5 W75 Wood Seasonal 7.24 194 0.9 70.2 2.1 1.1 4.9 12.7 13.0 W85 Crop Semiperm 7.07 504 3.5 125.5 111.0 4.2 27.7 32.6 18.9 W86 Crop Permanent 7.91 577 6.1 93.3 140.8 10.6 38.4 23.3 23.9 W87 Crop Seasonal 6.84 399 9.0 99.1 34.5 4.7 16.4 21.7 33.9 W88 Crop Semiperm 7.19 677 7.0 135.4 167.4 11.0 57.8 40.3 42.6 W89 Crop Permanent 8.16 1694 5.1 145.4 505.4 44.0 133.4 40.4 17.0 W90 Crop Semiperm 7.51 506 3.4 68.7 92.6 6.1 20.1 27.0 12.6 W91 Crop Seasonal 6.96 975 13.3 130.0 329.3 18.6 68.7 74.5 39.2 W93 Crop Seasonal 7.09 391 6.8 115.1 31.9 3.2 16.1 24.0 33.2 W96 Grass Seasonal 7.15 868 8.5 157.8 256.1 70.2 55.0 26.3 16.1 W98 Grass Permanent 7.56 1241 8.6 161.6 417.9 120.1 107.8 53.0 20.9 W100 Grass Seasonal 7.27 890 5.2 117.4 284.0 60.8 68.0 47.3 13.7 W101 Grass Permanent 7.06 792 3.7 95.1 248.2 54.7 47.5 53.1 12.7 W102 Grass Seasonal 7.80 865 5.8 212.1 177.6 24.8 54.5 34.3 19.6 68 Appendix B: continued ID Land Cover Permanence pH SC Cl HCO3 SO4 Na Mg Ca K (μS/cm) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) (mg/L) W103 Grass Permanent 7.98 584 7.6 114.1 108.7 12.7 43.5 27.1 18.9 W104 Grass Seasonal 7.94 376 7.7 124.8 6.3 1.2 10.3 18.0 25.9 W106 Grass Permanent 7.86 776 11.2 141.6 180.9 21.9 67.6 36.8 24.1 W107 Grass Permanent 8.12 1303 13.6 160.1 464.5 80.7 128.9 31.1 23.5 W108 Grass Seasonal 8.54 1557 46.3 116.8 742.6 100.9 166.1 135.9 32.2 W110 Grass Semiperm 7.17 520 9.5 53.6 156.8 27.8 28.9 20.4 13.6 W111 Crop Semiperm 7.37 795 15.1 139.5 191.4 12.1 47.0 49.8 55.8 W112 Grass Semiperm 7.00 806 11.5 122.8 273.6 51.2 49.2 49.9 22.0 W113 Crop Permanent 7.58 1396 10.1 163.1 538.6 61.7 137.1 60.3 24.3 W114 Crop Semiperm 7.13 349 8.6 118.8 30.5 4.6 15.1 21.4 33.1 W115 Crop Seasonal 7.08 314 11.6 131.6 8.8 1.5 8.3 19.6 39.1 W116 Crop Permanent 8.05 1780 8.2 232.9 488.9 50.3 149.4 28.7 29.6 W117 Crop Seasonal 7.23 771 4.2 162.0 187.6 6.6 52.9 71.4 22.2 W118 Crop Permanent 7.43 823 1.7 103.6 329.0 20.0 89.9 76.5 16.2 W119 Wood Permanent 8.24 307 1.5 165.5 7.7 2.9 19.5 15.2 16.5 W120 Grass Semiperm 7.73 865 16.0 127.0 284.3 57.5 67.6 34.0 20.5 W121 Grass Seasonal 6.59 98 2.0 33.7 2.9 0.6 3.7 8.3 9.0 W122 Grass Semiperm 6.95 262 3.1 108.6 18.9 1.9 13.5 12.8 11.2 69 Appendix C: Taxa List Continued (Part 1 of 4) Major Taxa To Family To Lowest Identification Amphipoda Gammaridae Gammarus lacustris Taltridae Hyalella azteca Hydrachnidia Platyhelminthes Rhynchobdellida Glossiphoniidae Helobdella elongata Anostraca Collembola Nematoda Oligochaeta Ostracoda Pelecypoda Sphaeriidae Pisidium Gastropoda Lymnaeidae Aplexa hypnorum Fossaria Fossaria/Stagnicola Pseudosuccinea Stagnicola Stagnicola elodes Physidae Aplexa hypnorum Physa Physa skinneri SWA 2008 16 92 6 52 334 185 2 2 15 106 7 30 1 1 SWA 2008 17 3 14 11 16 83 517 103 11 238 1 3 45 9 12 35 SWA 2008 18 5 5 2 8 1642 168 901 2 14 2 9 SWA 2008 19 7 182 1 SWA 2009 4 3 4 67 262 1 38 3 21 1 16 2 5 70 Smith Creek Watershed Site SWA SWA SWA SWA 2009 2009 2009 2009 5 6 7 8 402 4 4031 24 1 5368 2 2 17 5 59 730 20 64 4 5 45 4 9 17 10 14 4 2 1 8 2 6 5 SWA 2009 9 8 28 4 4 80 12 44 52 SWA 2009 10 4 146 6 102 1 SWA 2009 11 2 48 111 2 Total 3 414 4174 11 1 2 5516 30 1882 2402 1199 233 2 47 519 1 10 22 45 38 45 62 8 1 107 Appendix C: Taxa List Continued (Part 2 of 4) Major Taxa To Family To Lowest Ientification Planorbidae Gyraulus Promenetus exacuous Coleoptera Dytiscidae Agabus Colymbetes Colymbetes exaratus Hydroporinae Hygrotus Hygrotus sellatus Ilybius Liodessus Neoporus Oreodytes Rhantus Rhantus sericans Haliplidae Haliplus Haliplus immaculicollis Hydraenidae Hydrophilidae Berosus Helophorus Paracymus Diptera Ceratopogonidae Atrichopogon Pericoma/Telmatoscopus Chaoboridae SWA 2008 16 4 6 1 9 3 4 150 SWA 2008 17 10 2 6 13 2 3 2 3 5 49 9 3 110 2 SWA 2008 18 13 13 2 21 5 105 SWA 2008 19 1 8 SWA 2009 4 27 4 11 1 6 2 2 1 2 2 1 26 10 24 71 Smith Creek Watershed Site SWA SWA SWA SWA 2009 2009 2009 2009 5 6 7 8 3 2 2 1 5 30 1 26 2 2 26 1 2 10 1 3 1 109 26 9 2 14 16 SWA 2009 9 24 4 76 104 SWA 2009 10 4 38 SWA 2009 11 1 39 Total 83 19 2 27 50 1 2 26 5 2 6 2 26 2 3 3 38 4 6 2 71 15 3 698 26 144 Appendix C: Taxa List Continued (Part 3 of 4) Major Taxa To Family To Lowest Ientification Chaoborus Chironomidae Culicidae Aedes Dolichopodidae Empididae Muscidae Psychodidae Sciomyzidae Simuliidae Simulium Simulium venustum/verecundum Simulium vittatum Stratiomyidae Nemotelus Odontomyia Stratiomys Syrphidae Tabanidae Chrysops Tipulidae Tipula Ephemeroptera Baetidae Caenidae Caenis Hemiptera Corixidae Callicorixa audeni Cenocorixa dakotensis SWA 2008 16 497 114 9 1 10 10 3 58 10 1 2 2 41 1 SWA 2008 17 1415 26 6 7 2 16 135 279 120 17 56 1 1 6 203 1 17 2 SWA 2008 18 383 6 2 8 2 2 SWA 2008 19 115 3 40 55 1 SWA 2009 4 1 9 28 4 7 8 532 6 94 2 6 72 Smith Creek Watershed Site SWA SWA SWA SWA 2009 2009 2009 2009 5 6 7 8 26 2 1 1159 164 338 221 1 2 2 17 23 15 14 2 2 1 3 1 14 2 4 2 2 6 2 1 2 1 2136 23 17 17 2 1 SWA 2009 9 2 24 8 4 4 8 4 SWA 2009 10 56 2 SWA 2009 11 Total 30 24 4383 148 40 98 6 15 1 28 32 139 867 6 273 27 10 60 79 1 5 1 17 212 1 2182 79 5 1 Appendix C: Taxa List Continued (Part 4 of 4) Major Taxa To Family To Lowest Ientification Hesperocorixa Hesperocorixa atopodonta Hesperocorixa laevigata Hesperocorixa michiganensis Hesperocorixa vulgaris Sigara bicoloripennis Sigara conocephela Sigara decoratella Sigara solensis Lepidoptera Odonata Anisoptera Libellulidae Zygoptera Coenagrionidae Enallagma/Coenagrion Lestidae Lestes Trichoptera Leptoceridae Oecetis Limnephilidae Anabolia bimaculata Asynarchus Limnephilus Philarctus/Limnephilus Grand Total SWA 2008 16 3 1 1 1 1 2 11 1 4 67 46 2 64 2 1993 SWA 2008 17 2 3 2 4 15 1 46 2 577 2 34 4 4349 SWA 2008 18 1 2 4 3327 SWA 2008 19 1 1 2 15 433 SWA 2009 4 1 1 2 6 2413 73 Smith Creek Watershed Site SWA SWA SWA SWA 2009 2009 2009 2009 5 6 7 8 2 2 2 5 2 2 4 26 6 27 4 2 27 1 2 1 4 14 3 100 1 12712 1263 428 70 SWA 2009 9 12 8 4 4 576 SWA 2009 10 2 1 421 SWA 2009 11 2 Total 5 3 1 3 2 9 2 6 13 17 38 20 43 30 119 66 3 4 760 5 34 4 15 27985