RESPONSES TO SHRUB LOSS, ANNUAL GRASSLANDS, AND CRESTED WHEATGRASS SEEDINGS: MANAGEMENT
advertisement

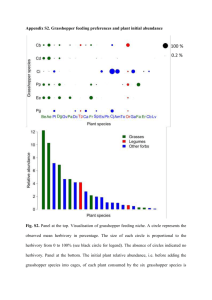
This file was created by scanning the printed publication. Errors identified by the software have been corrected; however, some errors may remain. GRASSHOPPERCO~TY RESPONSES TO SHRUB LOSS, ANNUAL GRASSLANDS, AND CRESTED WHEATGRASS SEEDINGS: MANAGEMENT IMPLICATIONS Dennis J. Fielding MerI~ A. Brusven ABSTRACT Grasshopper density and species composition were sampled at 42 sites arrayed along a disturbance gradient. Grasshopper density was lowest and species diversity was highest in vegetation types with shrub cover. Annual grasslands had the highest grasshopper densities and the lowest species diversity~ and were dominated by generalist species with wide diet breadths. Management concerns that arise from the different characteristics (food habits~ migratory propensity) of the dominant grasshopper species associated with the various plant communities were discussed. Management ofgrasshopper populations by habitat manipulation may be a viable alternative strategy. INTRODUCTION Grasshopper populations periodically reach outbreak proportions in the Intermountain region (Hewitt and Onsager 1983). In 1985, during a massive grasshopper outbreak, about 2.5 million ha of rangeland across southern Idaho were treated with broad-spectrum insecticides. This type of sledge-hammer approach to the control of insect pests applied on a landscape scale is becoming less acceptable. The undesirable aspects of broad-spectrum biocides (effect on nontarget arthropods, expense) make it compelling to find ways to manage grasshopper populations so that outbreaks are less frequent and of smaller extent. This project was undertaken to assess the role that range management actions have on grasshopper populations and to provide insights into the management of grasshoppers through habitat manipulation. Like many areas of the Intermountain region, southcentral Idaho has suffered extensive habitat degradation and shrub loss due to increased fire frequency associated with the invasion of cheatgrass. The diminished resource values that result from this process of shrub loss have been documented by many studies. The response of rangeland grasshoppers to the conversion of native vegetation to annual grasslands has not been studied in detail. There are well over 100 species of grasshoppers in the Intermountain region. Only four or five species attain very high densities and account for most of the outbreak populations. One of these, Melanoplus sanguinipes L., is particularly troublesome. In south-central Idaho, where there is an extensive interface between publicly owned rangeland and privately owned irrigated cropland, migration of grasshoppers from rangeland to cropland is a major problem. Melanoplus sanguinipes is well known for its propensity to migrate (McAnelly and Rankin 1986) and its broad range of food plants (Mulkern and others 1969) makes it a threat to a wide variety of crops, as well as rangeland forage species. Another abundant species in the Intermountain region is Aulocara eUiotti. It differs from M. sanguinipes in several respects: it is restricted to feeding on grasses only (Mulkern and others 1969) and is seldom found in cultivated crops. The objective of the present study is to identify patterns of grasshopper species composition among different vegetation types, and to determine whether the conversion of native plant communities to annual grasslands has an affect on the abundance of the major grasshopper pest species in south-central Idaho. MATERIALS AND METHODS Forty-two sites were sampled for plant and grasshopper species composition. These sites were all within the Davis Mountain SW USGS 7.5' quadrangle map (north of Bliss, ID). This area was selected because a wide spectrum of plant communities, from relatively undisturbed to dominance by exotic annual grasses (cheatgrass, Bromus tectorum, and medusahead, Taeniantherum asperum) and introduced perennial grasses (primarily crested wheatgrass, Agropyron cristatum), were all present within a small geographic area. The small scale of the study was intended to minimize the effects of local weather patterns on grasshopper community composition. The current year's standing crop by plant species was estimated by the weight-unit method (USDA-SCS 1976). Ten, 1-m2 quadrats were estimated at each site in July of 1990 and 1991. Both years' data were averaged for subsequent analysis. Paper presented at the Symposium on Ecology, Management, and Restoration of Intermountain Annual Rangelands, Boise, ID, May 18-22, 1992. Dennis J. Fielding is Postdoctoral Fellow and Merlyn A Brusven is Professor of Entomology, Department of Plant, Soil and Entomological Sciences, University of Idaho, Moscow, ID 83843. 162 Grasshoppers were sampled twice per year in June and late July and August. Densities were estimated by counting the number of grasshoppers flushed from 50, O.l-m2 quadrats. Species composition was determined by capturing and identifying at least 30 grasshoppers at each site. Density of individual grasshopper species was estimated by multiplying the species' proportions by overall density on the site. Pooled grasshopper data from the four sampling dates were used for subsequent analysis. Plant data were summarized and primary gradients identified using detrended correspondence analysis (DCA) (Hill 1980), an eigenvector ordination technique. Primary gradients in plant species composition were identified by nonparametric correlation (Spearman's r., Zar 1984) of shrub biomass, native perennial grasses (excluding Poa spp.), annual vegetation, and the percent exotic plant species with plant DCA axis-! scores for the 42 sites. Trends in grasshopper community composition were also examined by nonparametric correlation with the plant DCA a.xis-1 scores. Table 1-Mean plant aboveground biomass (gram dry weight/ square meter) by vegetation type Vegetation~~ Artr/ Agsp Artr/ Brte Artr/ Agcr Agcr/ Brte Taas/ Brte Bromus tectorum Bromus japonicus Taeniantherum asperum Agropyron cristatum Poa sandbergii Agropyron spicatum Other native grasses1 Artemisia tridentata Chrysothamus spp. Annual and biennial forbs Perennial forbs 5.2 2.8 <.1 .4 .8 14.9 4.8 34.4 1.9 .2 .7 11.5 1.5 1.4 .7 1.5 <.1 .8 33.6 3.2 .7 <.1 3.0 .5 .3 19.0 4.8 <.1 <.1 26.5 4.8 .2 <.1 5.4 <.1 1.8 21.8 5.7 <.1 .3 2.2 .5 1.3 .3 15.0 1.0 22.2 .4 2.5 <.1 <.1 .3 .2 3.4 <.1 Number of plant species 12.1 9.3 5.6 8.0 7.6 Plant 11ncludas E/ymus clnereus, Sitsnlon hystrlx, Slips thurberiana, and Agropy- ronsmithli. RESULTS The DCA ordination of the vegetation is shown in figure 1. The first axis, which accounted for 53 percent of the variation in plant data, may be interpreted largely as a disturbance gradient. The biomasses of shrubs and native grasses were negatively correlated with the plant DCA axis-! scores (r. =-0.79 and --0.72, respectively, N =42, P < 0.01), indicating less disturbed plant communities at the low end of axis-! (fig. 1). Biomass of annual vegetation and the percentage of aboveground biomass represented by exotic plant species were positively correlated (r. 0.59 and 0.91, respectively, N 42, P < 0.01) with the plant DCA axis-1, indicating the dominance by introduced plant species at the high end of plant DCA a.xis-1. Five somewhat subjective but nonoverlapping vegetation types were delineated on the ordination diagram and were labeled according to the two plant species with the greatest mean aboveground biomass within the vegetation type. Table !lists the mean composition of the five vegetation types. Grasshopper species composition also differed among the vegetation types (table 2). Melanoplus sanguinipes showed a strong affinity for the annual grassland sites where it accounted for 66 percent of all grasshoppers collected (table 2). Melanoplus sanguinipes did not comprise more than 15 percent of the population in any of the other vegetation types. Density of M. sanguinipes was strongly = = Table 2-Mean density and percentages of grasshopper species collected by vegetation type . - - - - - · ---------------------. ..... 260 • ·. 200. Artr/Agcr C\1 150 .• ' / ..\ Agcr/Brte • " ' ...... ~ /. tr/Agspt • < 50 ··· ... ·-. ~... :.~,~ :. Overall density (per m2) • ··.··.···I·····-..~··-. ·. . ·.~·.·. -.·.~- -· / 100 Density and species • ~·· .. \ Percentage of population: Agensotettix deorum Amphitomus coloradus Aulocara elliotti Clrcotettix undulatus Cordillacris occipitalis Cratypedes neglectus Dlssostelra spurcata Hespsrotettix viridis Melanoplus cinereus Melanoplus sanguinipss Oedaleonotus enigma Phoetaliotes nebrascsnsis Spharagemon equale Stenobothrus shastanus Trachyrachys kiowa Trimerotropis gracilis Trimerotropis psuedofasciata .···· • ........ ······~···a; ,./ • . ./_..-·· Ta;/Br.·te ~-··' • ....... I ~r/Brte _./ • oL---~----J-~·~=~·'----~·-~·~··--~--~----~ 0 50 100 150 200 25'0 300 350 Axls1 Figure 1-Detrended Correspondence Analysis ordination of 42 sites based on plant species aboveground biomass. Vegetation types are labeled according to the dominant plant species: Artr, big sagebrush (Artemisia tridentata); Agsp. bluebunch wheatgrass (Agropyron spicatum); Agcr. crested wheatgrass (Agropyron cristatum); Brte. cheatgrass (Bromus tectorum); Taas. medusahead wildrye (Taeniantherum asperum). 163 Vegetation type Artr/ Artr/ Artr/ Agcr/ Taas/ Agsp Brte Agcr Brte Brte 0.22 0.31 0.68 1.18 16 25 5 14 6 2 34 2 5 50 4 63 8 3 9 3 10 9 • 15 13 7 6 15 5 66 14 2 4 5 • 3 • 1 1 13 8 1 • • 5 2 1 2 .. 1.63 • • 13 2 • • • DISCUSSION correlated with the plant DCA axis-1 scores (fig. 2). Aulocara elliotti dominated the crested wheatgrass sites (table 2). Density of A elliotti also was positively correlated with axis-1 scores, although it reached its highest densities near the middle of plant DCA axis-1, where the Agcr/Brte sites were located (fig. 3). Both overall density and diversity (Shannon's H') of grasshoppers were strongly correlated with the plant DCA axis-1 scores (figs. 4 and 5). Sites high on plant DCA axis-1, the sites lacking sagebrush cover, had the highest grasshopper densities and the lowest diversity. In the lesser disturbed Artr/Agsp sites, 13 species comprised 95 percent of the grasshoppers collected from these sites (table 2). Progressively fewer species comprised 95 percent of the grasshoppers in the vegetation types arrayed from left to right along plant DCA axis-1 (fig. 1). Only four species accounted for 95 percent of the grasshoppers collected from the annual grassland sites. Striking differences were observed in the grasshopper assemblages associated with natural and introduced plant communities in south-central Idaho. The pattern of reduced biodiversity associated with the conversion to exotic annual grasslands (Whisenant 1990; T. Rich, these proceedings) was reflected in the grasshopper assemblages within the study area. Grasshopper assemblages were composed of progressively fewer species along the primary disturbance gradient in the plant communities. Sagebrush was probably the single most important factor affecting grasshopper community structure. The vegetation types with sagebrush were all characterized by low overall density and high diversity without any single species dominating, except A elliotti in the Artr/Agcr sites. Much of the observed patterns were the result of the distribution of two of the most common species, M. sanguinipes 2.5 4 r8 E ~ Cl) rs • 0.68 N E 3 Q) 1.5 ~ ~ CD 1 • ::l z • 0.5 Q) .c 2 •• • • E •• ::l 150 200 250 300 •• 0 350 • . . . • •• • .• 0 • '•• I • 50 100 Vegetation DCA Axis 1 r s • 0.53 CD c. 2 ~ - • • ••• >- ·a; E ~ • ~ 1.6 ~ 1.5 i5 ~ CD • •• ::l 0.5. • •• • ~ .. • 0 50 100 150 200 250 • • . . J . . _ - - L - - - L - - . - - - 1 - - . . L ... 200 250 .c 1 0 • 0 &1S ~ " • 300 • •• • • •• •• • • • 0 ._...JJ!..L.JI.1d~_!_l __ !.J_t_~___1_ 0 •• 150 ,., • • • • c. c. • .• • • 300 350 • •• CD E • ra • -0.70 ~ .c z •• • Figure 4-Relationship of overall grasshopper density to to vegetation DCA axis-1 soores. 2.5 2· • Vegetation DCA Axis 1 Figure 2-Relationship of M. sanguinipes density to vegetation DCA axis-1 scores. N • • • • 1. .--S--~~~~:--~--~----~-~ 100 • • z • • 50 • c. • • • ~ c. ~ 0.72 • 2 N • • • • • • • • • • •• 0.6 350 0 Vegetation DCA Axis 1 50 100 150 200 260 Vegetation DCA Axis 1 Figure 3-Relationship of A. e/liotti density to vegetation DCA axis-1 scores. Figure 5-Relationship of grasshopper diversity to vegetation DCA axis-1 scores. 164 300 350 all vegetation types. Examination of nymphal survey data from 1985, a year of extremely high densities, indicates that, although densities were high across south-central Idaho, the relationship between annual grasslands and higher grasshopper densities prevailed (Fielding and Brusven in press). In the Shoshone Bureau of Land Management district in 1985, annual grasslands averaged 41 grasshoppers/m2 compared to about 221m2 in sagebrushcovered areas (Fielding and Brusven in press). It may be argued that when densities exceed a certain threshold it does not matter whether there are 20 or 40 grasshoppers per square meter, it will be enough to cause hardships for farmers and ranchers. However, the evidence to date suggests that outbreaks would be less frequent, less intense, and cover less area in habitats with shrub cover than on frequently burned, cheatgrass-dominated landscapes. If further research confirms that annual grasslands do experience more frequent grasshopper outbreaks, then rehabilitation of annual grasslands with shrubs and perennial grasses should be considered as a means of noncatastrophic management of grasshoppers. Because migration of M. sanguinipes from rangeland to irrigated croplands is a primary rationale for control operations in south-central Idaho, some of the highest priority areas for rehabilitation would be those areas adjacent to croplands. Although the costs of grasshopper control alone may not justify the expense of planting shrubs and perennial grasses over large areas, the benefits to game birds such as pheasant (Sands, these proceedings), other wildlife, and livestock may be enough to justify rehabilitation of high-priority areas. The management of grasshoppers by the manipulation of vegetation has the advantage of being an environmentally sound, long-term strategy that could benefit many other resources. and A elliotti. The annual grasslands were dominated by M. sanguinipes. Aulocara elliotti dominated the crested wheatgrass seedings. The relative abundance of their preferred host plants may account for much of their observed habitat preferences. Both species can subsist largely on cheatgrass early in the season, then switch to other foods as the cheatgrass dries. Crested wheatgrass is readily accepted by A elliotti later in the season, while M. sanguinipes feeds largely on weedy forbs later in the season (Fielding and Brusven 1991). Annual and biennial forbs were most abundant on the annual grasslands (table 1). The different life history strategies exhibited by M. sanguinipes and A elliotti result in different management implications for these species. Aulocara elliotti is more of a specialist adapted to exploit a perennial grass resource. As a member of the grass-feeding subfamily Gomphocerinae, it is restricted in its host range to grasses. Phenologically it is well adapted to the perennial grasses of the area, maturing at about the same time as the plants. It appears that the life history strategy ofA elliotti is to specialize on a perennial resource, remain in a resource patch and tolerate conditions during adverse periods. Because A elliotti tends to mature at about the same time as its host plants, it is less likely to migrate off the rangeland in search of more suitable habitat. In contrast, M. sanguinipes is a much more opportunistic feeder, and populations hedge their bets with a wide spread in hatching dates. A large proportion of the population of M. sanguinipes will mature well into the summer when most plants have dried, leading to a situation where they will be much more likely to migrate to irrigated cropland. Therefore, where migration to cropland is a concern, then high populations of M. sanguinipes may be considered undesirable. However, if destruction of forage grasses is the primary concern, then populations ofA elliotti will compete directly with livestock for available forage grasses, whereas M. sanguinipes will tend to feed first on less desirable weedy forbs. Because populations ofA elliotti are usually more tightly synchronized than populations of M. sanguinipes (Onsager 1987), it is easier to assess the potential for damage from A elliotti early in the season instead of waiting for hatching to be completed, as is the case forM. sanguinipes where a substantial proportion of the population may already be in the fourth instar (the ideal time for treatment) before all hatching has been completed. Because of its high reproductive potential, it is probable that M. sanguinipes will be the dominant species in most vegetation types during outbreak years. However, in those habitats with a more equitable distribution of species, M. sanguinipes may be not be able to attain its full potential for explosive population growth. Few studies have found evidence of direct competition between grasshopper species; it seems especially unlikely that populations of M. sanguinipes will be inhibited by other grasshopper species in habitats where M. sanguinipes already dominates. These results indicate that areas with shrub cover and an understory of perennial grasses will have lower overall grasshopper densities with a lower proportion of pest species. These data were taken during years of low grasshopper density; it may be expected that during outbreak years grasshopper density may exceed treatment thresholds in REFERENCES Fielding, D. J.; Brusven, M. A.1990. Historical analysis of grasshopper (Orthoptera: Acrididae) population responses to climate in southern Idaho, 1950-1980. Environmental Entomology. 19: 1786-1791. Fielding, D. J.; Brusven, M.A. 1992. Food and habitat preferences of Melanoplus sanguinipes and Aulocara elliotti (Orthoptera: Acrididae) on southern Idaho rangeland. Journal of Economic Entomology. 85: 783-788. Fielding, D. J.; Brusven, M. A. [In press]. Spatial analysis of grasshopper density and ecological disturbance on southern Idaho rangeland. Agriculture Ecosystems & Environment. Hewitt, G. B.; Onsager, J. A 1983. Control of grasshoppers on rangeland in the United States-a perspective. Journal of Range Management. 36: 202-207. Hill, M. 0.; Gauch, H. G., Jr. 1980. Detrended correspondence analysis: an improved ordination technique. Vegetatio. 42:47-58. McAnelly, M. L.; Rankin, M. A.1986. Migration in the grasshopper Melanoplus sanguinipes (Fab.). I. The capacity for flight in non-swarming populations. Biological Bulletin. 170: 368-377. Mulkern, G. B.; Pruess, K. P.; Knutson, H.; Hagen, A. F.; Campbell, J. B.; Lambley, J.D. 1969. Food habits and 165 preferences of grassland grasshoppers of the north central Great Plains. Bull. 481. Fargo, ND: North Dakota State University Agricultural Experiment Station. 32 p. Onsager, J. A. 1987. Integrated management of rangeland grasshoppers. In: Capinera, J. L., ed. Integrated pest management on rangeland. Boulder, CO: Westview Press: 196-204. U.S. Department of Agriculture, Soil Conservation Service. 1976. National range handbook. Washington, DC: U.S. Department of Agriculture, Soil Conservation Service. [n.p.] 166 Whisenant, S. G. 1990. Changing fire frequencies on Idaho's Snake River Plains: Ecological and management implications. In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., compilers. Proceedingssymposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INI'-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 4-10. Zar, J. H. 1984. Biostatistical analysis. Englewood Cliffs, NJ: 2d ed. Prentice-Hall. 718 p.