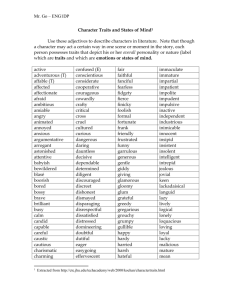
Part 3: Biological Distance
advertisement

1 Chapter I INTRODUCTION The purpose of this study is to examine the biological distance between native Californian populations involved in a series of prehistoric wars in the Santa Clara Valley. The wars occurred between AD 500 and AD 700, and it was during this era that the bow and arrow were introduced to the region (Moratto 1984), possibly as a result of largescale population movements between California’s Central Valley and the San Francisco Bay and Delta. One of these movements has been characterized by Bennyhoff (1968) as the Meganos Intrusion. While controversial in several respects, the basis of the Meganos hypothesis is evidence of a westward expansion (ca AD 500) by a Proto-Miwok group from the San Joaquin Delta into the Livermore and Diablo Valley regions, the Walnut Creek area, the East Bay, and then southward into the Santa Clara Valley. The Meganos intrusion into the South Bay lasted perhaps two centuries. Then, at the end of the Middle to Late Period transition (ca AD 700), the Meganos people were in turn displaced by a population movement from the north and returned to the Delta. There, they became the probable ancestors of the Plains Miwok (Bennyhoff 1961, 1968). The course of California prehistory remains unclear in many respects because of persistent problems in establishing an accurate chronology for most of its cultural sequences and in identifying the nature and dimensions of population movements prior to European Contact. Thus far, the clearest evidence of these movements has been derived 2 from linguistic studies, which indicate that much of California was inhabited by peoples who spoke languages in the Hokan family (Moratto 1984) prior to 4300 BC However, between 4300 BC and AD 1700, the Hokan speakers were apparently displaced by several intrusions of peoples from the north who spoke languages in the Penutian family (Dixon and Kroeber 1913, 1919) and represented a more robust physical type (Bennyhoff 1986). Later linguistic studies (e.g., Shipley and Smith 1979) suggest that the four major California Penutian stocks (Utian, Yokuts, Wintuan and Maiduan) represent four distinct migrations into California from the north, with an original center of dispersal located in either the Columbia Plateau or the northwest Great Basin. The Meganos people, according to Bennyhoff (1968, 1986), were one such Penutian-speaking group. The term ‘Meganos’ is borrowed from the Spanish phrase for ‘sand-hill people’ and describes the sites of most Meganos-style burials along the waterways of the Delta. The people they displaced in the Santa Clara Valley were Hokan-speakers who may have been ancestral to the Esselen (Breschini and Haversat 1997). These intrusions transformed the cultural landscape of prehistoric California. The introduction of the bow and arrow to Central California in or about AD 500, for instance, appears to have coincided with the end of the Meganos Intrusion (Bennyhoff 1968, Moratto 1984), and with peaks in interpersonal violence, as seen in analyses of the violent trauma recorded in human remains (e.g., Jurmain and Bellifemine 1997). It is increasingly clear that the Penutian intrusions were gradual and peaceful in some areas, but stiffly resisted in others by some of the Hokan-speaking groups already living in California (Atwood and DiGiuseppe 2005). By using the linguistic evidence as a 3 framework for further research in other fields, we may be able to clarify the events involved and document the changes in physical and cultural traits that occurred here as a result of conflict, displacement, cultural diffusion and gene flow. The Meganos Intrusion provides an interesting test case. Thus far, the differences between sites linked to the ancestral Esselen and the Meganos people have been based almost entirely upon burial practices and grave goods. The biological distance between the two groups has not been adequately quantified. Two sites in Contra Costa County (CA-CCO-139 and CA-CCO-141) that were first characterized as Meganos sites had both been looted extensively before a formal excavation could be carried out by Heizer (1949) and no osteological data were recorded for the remaining undisturbed burials. At more recently discovered sites, such as the one at Santa Rita Village excavated by Wiberg (1984), restrictions imposed by the Native American Graves Protection and Repatriation Act under Public Laws 101-185 and 101601 prevented osteological analysis prior to reburial. A limited analysis (Marshall 2002) of long bone metrics collected from a single Meganos site (CA-ALA-343) in Alameda County attempted to quantify the biological distance between the remains from Meganosstyle and non-Meganos burials at that site. However, the results were inconclusive due to extensive post-mortem damage to the majority of the bones in question. The study proposed here will instead rely on a comparative meta-analysis of metric and nonmetric (discrete) cranial traits seen in samples from six sites spanning the Central Valley, San Francisco’s East Bay and the Santa Clara Valley. The temporal range of the study will extend from the Early Middle Period (ca. 900 BC) to the Late 4 Period Phase 2 (ca. AD 1850). See Chapter 2, pp. 6-9, for a discussion of the chronological schemes applied to California archaeological sites by various authors. See Table 1, p. 8, for a summary of the chronological scheme developed by Millikan and Bennyhoff (1993) that was used in this study, Of the six sites included in this study, one from the southern reaches of San Francisco’s East Bay yielded burials with Meganos-style characteristics as well as others with non-Meganos characteristics. That location is the site already mentioned, CA-ALA343. Two sites in the Santa Clara Valley that lack Meganos characteristics include CASCL-674 and CA-SCL-137. Sites in the Central Valley associated with proto-Miwok, Miwok and Yokuts groups include CA-SAC-117, CA-SJO-91 and CA-SJO-105. If the Meganos Intrusion described by Bennyhoff and Fredrickson (1994) did occur, then remains from the Meganos-style burials at CA-ALA-343 will demonstrate less biological distance from those at proto-Miwok, Miwok and Yokuts sites in the Central Valley and more from those at the two proto-Esselen sites in the Santa Clara Valley. The purpose of this study is, therefore, twofold: (1) to measure the biological distance between prehistoric populations in Central California associated with the socalled Meganos Intrusion; and (2) to assess the relative merits of using metric versus nonmetric cranial traits in this analysis. 5 Chapter II LITERATURE REVIEW Part 1. Archaeological Background In California, archaeological investigations have made great progress in characterizing the material culture and resource usage of the state’s prehistoric populations since their inception in the late 19th and early 20th centuries. However, those investigations have yet to establish a clearly defined chronology for most of California’s cultural sequences. Indeed, the absence in large part of maize agriculture, large scale or durable architecture, sophisticated ceramics, and large scale tribal organization led early researchers such as Kroeber (1925) to conclude that human occupation of the state probably only extended back over the space of perhaps three or four thousand years. Kroeber (1925) further concluded that any prehistoric cultural changes that had occurred were minor in nature. In his view, cultures lacking agriculture as well as the highly developed technologies and social organizations he’d seen in the Southwest were primitive and therefore probably of recent derivation. It did not occur to him that in an environment so rich in food and other resources, the material and social ‘sophistication’ achieved by the Pueblo peoples was simply unnecessary. Better dating techniques and excavation technologies have since made it clear that California prehistory extends back to at least the Clovis period of 9,000-11,000 BC (Chartkoff and Chartkoff 1984). It has also become quite clear that extensive changes in 6 subsistence, settlement patterns, social organization and technology have taken place since that time (Heizer 1949, Elsasser and Breschini 1968, Fredrickson 1974, Moratto 1984). However, the exact sequence of change and the population movements associated with them are still hotly debated. Chronology Any discussion of prehistoric population movements will require reference to a chronology system, the first of which was proposed by Lillard, Heizer and Fenenga (1939) and later refined by Heizer (1949). Heizer’s (1949) chronology divided California prehistory into a time frame with four divisions, the Late Horizon, Phases I and II, the Middle Horizon and the Early Horizon. Since then, various modifications and refinements of this scheme have been proposed (e.g., Moratto 1984, and Bennyhoff and Frederikson 1994). For the purposes of this study, a modified form of the Central California Taxonomic System (CCTS), originally devised by Robert Beardsley (1948:49), will be used to provide temporal reference points. The modifications are those proposed by Milliken and Bennyhoff (1993), and are based on their analysis of stylistic changes seen over time in grave goods and in ornamental shell beads in particular. A summary of this chronology is shown below in Table 1. A single sequence, as shown here, is probably unrealistic, given the complexity of the population movements and pattern changes now known to have occurred in some parts of the state. However, the CCTS corresponds fairly well with Heizer’s (1949) original system, and simplifies discussion of archaeological data from several different research paradigms if one simply substitutes the term ‘period’ for ‘horizon’ as needed. 7 Table 1. Chronology for the Prehistoric San Francisco Bay Area. California Cultural Horizons After Heizer 1949 Late Horizon Phase II Late Horizon Phase I Central California Taxonomic System (CCTS) After Beardsley 1948 Historic period European Contact (End Augustine Pattern) Late period Phase 2-B Late period Phase 2-A Late period Phase 1-C Late period Phase 1-B (End Meganos Pattern) Late period Phase 1-A Middle to Late Period Transition Middle Period Terminal Phase (Begin Augustine Pattern) (End Berkeley Pattern) Middle Period Late Phase Middle Period I Intermediate Phase Middle Period Early Phase Early to Middle Period Transition (Begin Meganos Pattern) (Begin Berkeley Pattern) (End Windmiller Pattern) Middle Horizon Early Horizon Revised Dating Scheme After Bennyhoff and Hughes 1987 Beginning ca. AD 1800 AD 1775 Temporal Categories Used In this Study After Milliken and Bennyhoff 1993 Late Period Phase 2 AD 1700 AD 1500 AD 1300 AD 1100 Late Period Phase 1 Middle to Late Transition AD 900 AD 700 AD 500 Upper Middle Period AD 300 AD 100 Lower Middle Period 200 BC 500 BC Early Middle Period 1000 BC 1500 BC Early Period (Begin Windmiller Pattern) 2500 BC 3000 BC 3500 BC Within this framework, Fredrickson (1973) defined various patterns of material culture. Within the bounds of this study, the Augustine, Berkeley and Windmiller Patterns are those seen at the selected sites. The earliest of these is the Windmiller Pattern, named for its typological site on the Sacramento River (Ragir 1972). The 8 Windmiller culture emerged sometime between 3,500 and 2,500 BC (Moratto 1984) and lasted until AD 1000 to 1500. Windmiller sites are usually located on low rises or knolls in the flood plains of creeks and rivers, and Windmiller material culture, in general, reflects an adaptation to wetlands habitats. It is associated with a seasonal settlement pattern, burials dominated by primary inhumation in a ventrally extended position, large non-obsidian stemmed projectile points (spear and dart tips), and the use of manos and metates to grind grass seeds. Baked clay net sinkers, as well as bone fish hooks and spears, and bone awls and needles have been recovered, although bone artifacts in general are rare. Charm stones, quartz crystals, Haliotis shell ornaments, and Olivella shell beads are commonly found in Windmiller burials (Ragir 1972, Moratto 1984, Fenenga 2009). The Middle Horizon Berkeley Pattern also demonstrates a seasonal settlement pattern (Moratto 1984) and is frequently associated with coastal shellmounds. Berkeley traits include the use of fishhooks and spear points, burials (often in village middens) with remains curled up in a flexed position, dart projectile points, and a shift from small lightweight manos and metates to mortars and pestles, indicating a greater reliance on acorns. At Berkeley Pattern sites (Moratto 1984), grave goods are less frequent than at either Windmiller or Meganos sites. When grave goods are present, shell ornaments and beads, and bone artifacts such as fish hooks and spears, occur with greater frequency than at Windmiller sites (Fenenga 2009). The Augustine Pattern (Moratto 1984) is the most recent, arising between AD 500 and AD 1000 and persisting until European Contact, which took place in AD 1775 in the 9 San Francisco Bay area. The Augustine Pattern is associated with the development of semi-permanent villages, more intensive reliance on acorns, the use of beads as money, and the use of the bow and arrow. At one time, the patterns mentioned here were thought to correspond closely with Heizer’s (1949) Culture Horizons. However, more recent work has shown that prehistoric cultural successions in Central California were much more complex (Moratto 1984, Johnson 2003). Population Movements A major point of interest for this study is the hypothesized intrusion of Penutianspeaking peoples into California from the north, possibly originating in the Columbia River Basin (Chartkoff and Chartkoff 1984, Moratto 1984). Penutian speakers are thought to have moved into the marshy lower Sacramento Valley between 3500 BC and 2500 BC, a movement associated with the development of the Windmiller Pattern (Fredrickson 1974). It is now generally accepted (see Breschini 1983 for a thorough review of the issue) that over the following centuries, as the Penutians spread out and diversified, the earlier Hokan-speaking groups were forced into peripheral resource zones and higher elevations. The Penutian-speaking groups eventually occupied the entire Central Valley. They are also believed to have expanded westward into the San Francisco Bay Area and southward as far as the Monterey Peninsula (Moratto 1984). The map in Figure 1 shows the linguistic groupings of Central California tribes at the time of European Contact as currently understood (Breschini and Haversat 1997). This displacement of earlier peoples was neither swift nor straightforward. In some cases, Penutian-speaking groups appear to have migrated south and then moved 10 northward again (Breschini 1983, Moratto 1984). Gerow (1974) describes the migration process as gradual and states his belief that many Hokan speakers gradually ‘coalesced’ into Penutian-speaking groups, but many others report that warfare was common at various points throughout this period (Chartkoff and Chartkoff 1984). Peaks of interpersonal violence were associated with some transitions, particularly in the San Francisco Bay Area (Jurmain and Bellifemine 1997, Jurmain 2001, Atwood and DiGiuseppe 2005), where excavation and osteological analysis has revealed an increased frequency of injuries consistent with warfare as well as projectile points found embedded in bone. 11 Figure 1. Linguistic Families of the California Culture Province (after Kroeber 1925). As noted by Breschini (1983:70), the intrusion of Penutian speakers into areas held by Hokan speakers appears to have taken place along the Central California Coast only where a specific combination of environmental factors was present. The necessary factors appear to have been a combination of relatively level areas of oak grassland in proximity to either the ocean or the San Francisco Bay, and sizable areas of marshes. 12 One element of this complex and multiform Penutian intrusion is that which Bennyhoff (1968) termed the ‘Meganos Intrusion.’ Probably an offshoot of the larger Penutian movement, it occurred along the eastern shores of San Francisco Bay and then moved south into the Santa Clara Valley. Basing his hypothesis on differences in burial type and position, grave goods, and the more robust physical type he observed in Meganos remains, Bennyhoff (1968, 1986, 1987, 1994a, b) believed that during the Middle Period’s Late Phase (AD 300 to AD 500), the Walnut Creek Valley and a region stretching westward to the shores of San Francisco Bay, and southward past the Monterey Bay area, was inhabited by, among others, a Hokan-speaking group that may have been ancestral to the Esselen (Breschini and Haversat 1997). Around AD 500, these people were displaced by the Meganos people, a Penutian-speaking group which originated in the San Joaquin Delta and expanded westward into the Livermore and Diablo Valley regions, the Walnut Creek area, the East Bay, and southward into the Santa Clara Valley (Bennyhoff 1986). Bennyhoff (1994a:7) thought the sites used by the Meganos people exhibited a fusion of the Windmiller (see Figure 2) and Berkeley (see Figure 3) patterns of material culture. The fusion resulted in a distinct new pattern. 13 Projectile Points Shell Ornaments and Beads Bone Artifacts Ground Stone Art ifacts Figure 2. Windmiller Pattern Artifacts from CA-ALA-343 (Hylkema 2002) GRAVE GOODS Projectile Points Shell Ornaments and Beads Bone Artifacts Ground Stone Artifacts AD 500 UPPER MIDDLE AD 300 LOWER MIDDLE 500 BC EARLY 2000 BC Figure 3. Typical Berkeley Pattern Artifacts (Hylkema 2002) Meganos grave goods (Bennyhoff and Frederickson 1994: 81) are characterized by large projectile points, and by shell and stone beads. Bone artifacts (Fenenga 2009) are far more common than is seen in Windmiller sites, and there are fewer ground stone 14 artifacts. Charm stones (Bennyhoff and Frederickson 1994: 81) are present, and exhibit greater complexity in stylistic elements than is generally seen in the Windmiller Pattern, frequently including the presence of a hole drilled through one end of the stones. According to Moratto (1984:120), a typical burial position for remains associated with the Windmiller Pattern is extended, with the deceased sometimes placed in a prone or face-down position, but more commonly in a supine or face-up position in a long, narrow grave (as shown below in Figure 4). The head is oriented toward the west, and the remains are often accompanied by a variety of grave goods which may include shell beads, charm stones or quartz crystals. Female burials may also contain manos and metates, while male burials may contain projectile points and knives. Bennyhoff (1986) describes Meganos burials in similar terms, with one exception: he specifies a northerly rather than a westerly orientation for the head. Figure 4: Typical Windmiller burial position (after Moratto 1984:120) According to Beardsley (1971:167), the skeletal remains from typical Windmiller burials indicate “a large-boned, rugged population with long, high heads and broad noses” and Bennyhoff (1986) describes Meganos remains in nearly identical terms, noting a difference in physical type as one of the chief characteristics distinguishing Meganos from non-Meganos burials. 15 In contrast, the sites used by the ancestral Esselen are characterized by the Berkeley Pattern (Bennyhoff and Frederickson 1994:83), including flexed burials, sparse grave goods, ceremonial burials of birds and animals, and burials within village middens. Grave goods may include some of the Berkeley Pattern artifacts shown in Figure 3. A typical burial position for remains associated with the Berkeley Pattern (Moratto 1984) is flexed, as shown in Figure 5, with the deceased placed on one side, and the head oriented in any direction. The remains themselves are described by Bennyhoff (1986) and several others as being more gracile than those of either the Windmiller or the Meganos people, and as having more rounded heads and narrow noses. Figure 5. Typical flexed burial position (from Grady, Latham and Andruschko 1999:124) The Meganos intrusion into the South Bay lasted perhaps two centuries. Then, at the end of the Middle to Late Period transition (ca AD 700), the Meganos people were in turn displaced by population movement from the north. According to Bennyhoff (1986), this intrusion was the result of pressure from a third group, the ancestral Patwin. The Patwin supposedly moved into the Solano area, bringing the Augustine pattern along with them, and forcing the ancestral Bay Miwok into the Walnut Creek Valley. In turn, the Bay Miwok pushed the Meganos people southward, out of the Walnut Creek Valley. A Meganos site adjacent to the Carquinez Straits in modern-day Fernandez (CA-CCO-2), as 16 well as another in El Sobrante, were then also abandoned by the Meganos people, who all ended up moving east to reoccupy ancestral lands in the Stockton area. There they purportedly became the Plains Miwok and/or Valley Yokuts (Bennyhoff 1994a, 1994b). The outlines of these hypothetical movements are depicted in Figure 6, in maps redrawn from Bennyhoff and Fredrickson (1994). Significance of the Meganos Intrusion The Meganos Intrusion is significant for several reasons as it appears to demarcate the boundaries of a lengthy and presumably episodic war in the East and South Bay caused by intense competition for resources (Bennyhoff 1994b). Ongoing research (Beardsley 1954, Jurmain and Bellifemine 1997, Jurmain 2001, Schwitalla 2010, Atwood and DiGiuseppe 2005) indicates high levels of violence occurring in conjunction with these movements, and includes evidence of trophy-taking (Andrushko, Latham, Grady, Pastron and Walker 2005) at CA-ALA-343, a site central to this study. Indeed, Bennyhoff (1986) thought the success of the Patwin invasion was largely due to their introduction of a new weapon – the bow and arrow – which gave them an edge in the ongoing conflict and eventually replaced the atlatl throughout California. Chartkoff and Chartkoff (1984) consider the arrival of the bow and arrow ‘a cultural watershed’ and likewise place its date as approximately AD 500 in the San Francisco Bay Area. While the Meganos Intrusion is considered by some researchers to be an artifact of poor sampling techniques and insufficient sample size (e.g., Johnson 2003), other researchers 17 A: Emergence of the Meganos (500 BC – AD 100). C: The Meganos retreat to the Delta (AD 700-900). B: Meganos expansion during the Middle Horizon (AD 300-700). D: The end of the Meganos era (AD 900-1100). Figure 6: The Meganos intrusion into and departure from the San Francisco Bay Area (maps redrawn from Bennyhoff and Fredrickson 1994). 18 (Wiberg 1984, 1992) have continued their efforts to characterize the Meganos sites and to explore the levels of violence associated with the Intrusion (Atwood and DiGiuseppe 2005, Schwitalla 2005). Burial types and grave goods from the Upper and Lower Middle Periods (AD 300-900) and this area are now known to be more variable than Bennyhoff more gracile foothill-dwelling Hokan speakers has been described by several believed at the time. However, the osteological data from the relevant burials have not undergone a rigorous archaeologists, including Heizer (1949), Beardsley (1948) and Johnson (1994), and confirmed by osteological analyses, which will be described in detail in Chapter 2, Part 2, in the section on previous cranial studies. However, with the exception of Suchey (1975), all previous studies of biological distance between the various prehistoric populations in Central California have been quite limited in scope. In most cases, relatively small osteological samples were used from no more than three or four individual sites. With the improved analytical and statistical techniques now available, it would be informative to compare both metric and discrete traits of crania recovered from several relevant sites in Central California. 19 Part 2. Previous Studies of Prehistoric California Cranial Types The earliest studies of skeletal material from prehistoric California sites were done by Gifford (1926), Hrdliçka (1927), and Von Bonin and Morant (1938). All of these studies used craniometric measurements and indices to examine geographic variability among prehistoric inhabitants of the Central Valley. Despite a preconceived notion that the aboriginal physical type was wide-headed and broad-faced, both Gifford (1926) and Hrdliçka (1927) found that cranial morphology varied geographically among Central Valley populations. However, these preliminary analyses gave no consideration to the age of the subjects, temporal variation or possible genetic relationships among populations, and because of this, the results were strongly biased by sample selection. The first study of a skeletal series from Central California that did take time and space into consideration was conducted by Fenenga (1939). He found significant variation in crania from the Early, Middle and Late periods in that cranial and nasal indices both underwent a progressive change over time. Fenenga found that individuals dating to the Early Horizon were characterized by a dolichocephalic (elongate) head and platyrrhine (wide) nose. Crania dating to Heizer’s (1949) Phase I and Phase II of the Late Horizon were characterized by a rounder brachycephalic shape and mesorrhine (narrow) noses. On this basis, Fenenga concluded that Central California populations included two physical types during the Middle Horizon – one dolichocephalic and one brachycephalic. This difference in cranial shapes, as it turned out, correlated with variations in artifact typology in the same sites, and was later used by Heizer (1949) to 20 help define the Early, Middle and Late Horizons in his chronological scheme (Moratto 1984). Another study of skeletal series from Central California was carried out by Beardsley (1948), who compared the cranial attributes of samples from the Central Valley with those from the San Francisco Bay Area. His longitudinal results were based on the temporal origins of the specimens he examined, and showed that individuals associated with the Windmiller Pattern during the Early Horizon were large-boned and had long heads and broad noses. Individuals from the later Middle Horizon samples were transitional, and individuals from the Late Horizon had dolichocephalic (elongate) heads and narrow noses. When Beardsley (1948) considered space as well as time, he found two physical types present in the Central Valley and the San Francisco Bay Area during the Late Horizon: individuals from Alameda County were consistently mesocephalic while individuals from the Cosumnes area were brachycephalic. Even so, Beardsley saw no evidence of wholesale replacement or sudden changes. Instead, he suggested a gradual change had taken place through a genetic admixture of interior and coastal groups, possibly in the course of a partial population replacement of Hokan speakers by Penutian speakers. Additional studies were conducted by Newman (1957) on selected skeletal measurements from early Sacramento Valley populations. Expanding on an earlier study, Newman used craniometric data and morphological observations of 140 crania from 17 sites in California’s Central Valley. The sites were chosen to represent three cultural time frames: the Early, Middle and Late Horizons proposed by Heizer (1949). In order 21 to reduce bias caused by inaccuracies in sexing the crania, he also decided to use only the crania of adult males over 20 years of age, and to use only those crania that came from fairly complete remains, so that the pelvic bones could be used to verify the sex of each individual. Using both craniometric measurements and derived indices, Newman examined biological distance over time and attempted to construct a typological series. He found a significant decrease in skull size and general robustness from the Early Horizon (typified by the ‘Large Broad-faced’ cranial group) to the Middle Horizon, in which the ‘Long-faced’ and ‘Small Short-faced’ cranial types appeared and increased in number. He also found that cranial breadth increased during the transition from the Middle to the Late Horizon, when ‘Round-vaulted’ crania appeared. Newman (1957:20) concluded: “The evidence… favors the hypothesis that each horizon may have seen a new group enter and mix with the preceding people. In no case does this new group form as much as 50% of the population of the next horizon.” In this, Newman (1957) agreed with Beardsley (1948), who thought that some population intrusions had occurred, but did not result in wholesale replacement of earlier populations. Suchey (1975), however, criticized Newman’s sample selection, noting that Newman’s crania from Central Valley sites were not sufficiently separate in either time or space. Newman’s study lumped crania from several sites into one time scheme without consideration of geographical aspects of the sites. Because of this lumping, differences between Middle and Late Horizon crania might be due to either geographical or temporal variation. Suchey (1975) also noted that Newman did not fully discuss his 22 hypothesis concerning the possibility that an immigrant population had moved into Central California, nor did he say much concerning the possible nature, physical types or origins of any such group(s). Suchey found the Late Horizon data to be the most problematic because Newman lumped together some sites whose populations are known to have been culturally and linguistically distinct. Gerow (1974) also criticized Newman’s failure to fully describe the progressive changes in physical types over time. A more definitive model of Central California’s population dynamics was proposed by Gerow and Force (1969), who compared osteometric data from a site in University Village (CA-SMA-77) with previously excavated material from the Central Valley. Their results showed that individuals from the Bay Area series represented a small and narrow-nosed population with low cranial vaults while those from the Delta series were larger and had high cranial vaults. On this basis, Gerow and Force (1969) proposed an independent cultural succession sequence for the Bay Area during what they called the ‘Early San Francisco Bay Period.’ According to this model, people with two distinct physical types and cultural traditions coexisted during the Middle Horizon, from 1500 to 1000 B.C. However, Gerow and Force also thought their data supported a model of convergence over time among Central California populations. Another study by Gerow (1974) compared skeletal series from sites in the San Joaquin Delta and the Santa Barbara region. In this study, Gerow likewise found that a large and high-vaulted group inhabited the Central Valley, while a small, low-vaulted group inhabited the Santa Barbara coastal sites during the Early Horizon. During the Middle and Late Horizons, Gerow (1974) found evidence of convergence between these 23 two types as cranial sizes and proportions grew increasingly similar, a result he ascribed to gene flow between the groups. All of these studies were relatively small in scale, but a much more comprehensive study was conducted by Suchey (1975), who used 30 discrete traits derived from Berry and Berry (1967) to assess biological distance between sites along the coast in Southern California, the Bay Area and the Central Valley. Suchey scored a total of 1200 crania for this study, taking care to maintain the same ratio of males and females in all samples tested. She also restricted her sample subjects to those that could be sexed using postcranial skeletal elements and to those at least 16 years of age at time of death, based on closure of the spheno-occipital synchondrosis. Suchey then calculated a Mean Measure of Divergence (MMD) from her results. Suchey (1975) found that the coastal sites in Southern California showed no significant differences from each other. Suchey found more variation in data from the Central Valley sites, where remains from 63% of the sites shared similar cranial attributes. In particular, samples from a site in the San Joaquin Delta (CA-SJO-68) and a coastal site (CA-ALA-329) had a Minimum Means of Divergence (MMD) of 0.009, indicating great similarity. However, 37% of the Central Valley samples did differ significantly from each other. From this, Suchey concluded that there was as much biological diversity within the Central Valley as there was in the larger area including both the Central Valley and Southern California. This conclusion was then tested by comparing the range and distribution of individual traits from Delta crania with the variability seen in more than 100 samples drawn from all major geographic races, world-wide (Suchey 1975:82). The comparison showed that the range 24 and distribution of traits in the Delta were just as variable and diverse as they were in major geographic groupings from around the world. Suchey (1975) then used a multi-dimensional scaling program to categorize the skeletal samples according to the MMD results. Scores for discrete traits showed developmental differences among individuals and between interior and coastal sites, which Suchey (1975:114) attributed to intraregional variation that was probably due to both genetic and environmental influences. In particular, Suchey found that dietary elements and methods of subsistence were likely to be important factors in convergent evolutionary forces affecting discrete traits. For example, while coastal populations made great use of shellfish, and obtained substantial amounts of iodine from their diet, Sacramento River-based groups maintained a mixed hunting/fishing/gathering lifestyle and probably had less access to iodine, which can affect growth patterns. On the other hand, individuals inhabiting some interior Central Valley sites could obtain iodine from anadromous fish, such as salmon. Suchey (1975:117) concluded that nonmetric traits, by themselves, cannot provide a “clear understanding of the dynamics of prehistoric populations.” However, thus far, no complementary regional study using craniometrics has been published. Several small scale cranial studies have been done since Suchey’s (1975), using either craniometrics, nonmetric traits, or a combination of the two. Agler (1986), for example, used nonmetric traits to examine the biological distance between a site in Northern California (CA-TEH-10) and six Central California sites that were all included in Suchey’s (1975) study. In the process, Agler further examined these data for the 25 effects of inter-observer error in scoring nonmetric traits. Agler (1986) concluded that inter-observer error was high and sometimes causes significant distortions in the Mean Measure of Diversity. For this reason, more accurate results were obtained when problematic traits, such as the presence of a bony partition in the hypoglossal canal, were excluded from the meta-analysis. Agler (1986) also found that the Tehama County sample did differ significantly from half of the Central California samples. Overall, Agler (1986:114) concluded that “biological homogeneity increases through time in California prehistory.” This conclusion concurs with those of Beardsley (1948), Gerow and Force (1969), and Suchey (1975), who all found evidence of an overall change from heterogeneity in the Early Horizon to homogeneity in the Late Horizon. A study of biological distance using discrete cranial traits was conducted by Donahue (1993) on three prehistoric populations from the Chico area (Butte County) in order to assess their heterogeneity. In addition, Donohue examined the biological distance between these populations and others at sites located along the Sacramento River south of Butte County and in the Delta. The sites chosen by Donohue span the Early, Middle and Late Horizons, and one of her goals was to evaluate change over time in the levels of homogeneity seen in all these groups. Donohue (1993) concluded that nonmetric traits could provide useful measures of biological distance, especially when skeletal materials are damaged and fragmented. Donohue (1993:116) also found substantial heterogeneity between the three Butte County sites, which she described as being: “as different from each other as they are from the Delta sites.” In addition, Donohue’s results support Agler’s (1986) conclusion that homogeneity increased over 26 time across the region sampled. However, the analysis could not determine whether that increase in homogeneity was the result of convergence or population replacement. A craniometric study by Oeh (1996) looked for patterns of biological distance across the Early, Middle and Late Horizons (2500 B.C. to A.D. 1700), using samples from two coastal sites (the Ryan and Patterson Mounds) and one interior site (the Blossom Mound). The Ryan Mound is a San Francisco Bay shellmound of uncertain affiliation, with a Late Horizon component that yielded 61 crania. The nearby Patterson Mound is a Middle Horizon site that yielded 22 crania, and is associated with the Costanoan people. The Blossom Mound is an Early Horizon Central Valley site associated with the Plains Miwok which yielded 27 crania. Using measurements of 69 variables obtained from the 110 samples, Oeh (1996) used a multivariate statistical analysis to measure intergroup relationships. The study also tested measurement replicability, and the effects of age and sex on adult cranial morphology. Oeh’s results showed that intraobserver error was a problem for small measurements such as mastoid length, but there were no significant differences in cranial measurements that could be ascribed to age. Sexual dimorphism was significant, but the pattern of dimorphism was consistent across all three temporal horizons. Analysis of biological distance showed that the crania sampled from the Ryan Mound were smaller in size and less robust than those from both the Blossom and Patterson Mounds. However, the two neighboring shellmound groups are morphologically more different from each other than either one is from the Early Horizon site (the Blossom Mound) in the Central Valley. In fact, the Patterson Mound and the 27 Blossom Mound samples showed sufficient similarity that Oeh (1996:123) concluded there were probably genetic interactions between the two populations during the Early Horizon, possibly in the course of trade relations. Oeh further suggested that differences between the two shellmound sites might have resulted from different migration routes into the South Bay such that political independence, resource competition and/or language differences might have worked to maintain group separation despite the environmental similarity resulting from their proximity to each other. A geographic study conducted by Morgan (2001) examined biological distance between four Late Horizon Central California populations using cranial metric and discrete traits as well as dental metric traits. The four sites represented two distinct but related cultural groups within the Penutian language group: the Plains Miwok and the North Valley Yokuts. The four sites also represent two geographic settings, two deltaic and two riverine. Morgan’s results showed while biological variability persists in these four populations, there is also a degree of homogeneity that tends to confirm the Late Horizon convergence seen by other researchers in Central California, including Beardsley (1948), Gerow and Force (1969), Gerow (1974), and Agler (1986). However, Morgan (2001) found that Suchey (1975) was incorrect in suggesting that geography played a larger role than genetic relationships in shaping the divergence of Central California populations, at least in the Late Horizon. Instead, Morgan’s metric analysis showed that while there is a trend toward geographic regions being distinctive, the difference never accounted for more than 20% of the variation seen in the traits she examined. 28 Part 3: Biological Distance: Metric vs. Nonmetric Biological distance studies have become a mainstay of bioarchaeology, although the goals of such studies have undergone continual changes due to emerging techniques, new statistical methods and shifts to new centers of interest. Buikstra et al. (1990) reviewed recent trends in biological distance studies and identified the following major thrusts: (1) answering basic questions of evolutionary history; (2) addressing basic archaeological questions, e.g., about residence patterns or the stability of local cultures; and (3) assessing the biological basis of questions about paleodemography and paleopathology. An example of the evolutionary history approach would be Sciulli’s (1990) study of crania from the Duff site cemetery, a Terminal Late Archaic site in Ohio. He used both metrics and nonmetrics, and determined that all of his subjects were related. Variation was geographical in nature rather than temporal, with samples geographically nearer to each other being more closely related than samples from more distant sites. Another example would be Rothhamer and Silva’s (1990) study of craniometric variation across prehistoric South American populations, in which they found several correlations between population profiles and sex, temperature minima, and altitude. An example of the archaeological approach would be Conner’s (1990) study of nonmetric variability in Late Woodland sites in west-central Illinois. In this study, he found evidence of lower variability between sites than is seen in the Middle Woodland period, confirming a ceramic-based hypothesis about increased interaction between local populations in the later era. Another example would be Steadman’s (2001), who also 29 looked at gene flow in west-central Illinois, but during the transition from Late Woodland to Mississippian. Looking at metric and nonmetric data, she found that distances of 100 km or more between sites did not stop gene flow in the Late Woodland, but did in the Mississippian, and thought this was due to the period’s sedentary lifestyles, agricultural practices and social hierarchies. An example of the paleodemographic approach would be Konigsberg’s (1988) work on nonmetric traits of crania from the same area as Steadman (2001), wherein Konigsberg used an algorithm to split male and female data. He found more variability among the females at most prehistoric sites in west-central Illinois and Konigsberg interpreted this to mean the females were more mobile than the males, and therefore the residence pattern was patrilocal. There is no question, then, that biological distance studies have posed and attempted to answer some fascinating questions. What is still being debated is whether one should use metric or nonmetric traits for these kinds of studies. Some, such as Berry and Berry (1967), have touted the superior value of nonmetric to metric studies in various types of population studies, especially in archaeological contexts. They reviewed a number of early studies on nonmetrics, and found them superior to metrics for the following reasons. Discrete traits: (1) are highly genetic in nature; (2) vary even between closely related populations; (3) exhibit constancy under environmental variability; (4) do not vary with age; 30 (5) show no sex differences; (6) show virtually no correlation with each other; and (7) are easily defined, standardized and scored. Therefore, say Berry and Berry (1967), it is not really necessary to go to all the work of measuring bones, and data from subjects of different sexes and ages can be pooled for analysis. Others, like Sullivan (1922), have promoted the use of discrete traits in cases where artificial cranial deformation limits the use of metrics, while Saunders (1989) argues for their use where only bone fragments are available for study. Several studies and reviews have now qualified all of the assertions made by Berry and Berry (1967) concerning nonmetric traits. Others have pointed out a number of problematic aspects which need to be addressed in designing studies which include nonmetrics. A summary of these studies follows. Heritability Problems with heritability affect both metric and nonmetric traits. As Buikstra et al. (1990:6) stated, there are both environmental and polygenic components involved in the development of any trait, and in the cranium “we are dealing with quantitative traits that are probably only moderately heritable.” However, Buikstra et al. (1990) state that this does not mean that either metric or discrete traits will not work, only that low heritability will cause a lot of noise in the data. There is good reason to believe that phenotypic and genotypic covariance is proportional in many cases. Cheverud et al. (1979), for instance, compared 48 31 craniometric traits and 25 discrete traits in four Amerindian populations, and found every nonmetric trait was correlated with its threshold (metric) trait in at least one population. Therefore, they concluded that there is no biological reason to favor either type of trait in studies of population structure. Some traits are definitely more heritable than others, but the reasons why are still unclear. In one of the few studies done on other primates, Cheverud and Buikstra (1981a, 1981b) looked at 135 mother-child pairs of macaques and found that heritabilities for hyper/hypostotic discrete traits are significantly higher than for foraminal traits. However, the authors noted that the large standard errors in their results serve to weaken their argument that the differences are caused by trait etiology. Cheverud and Buikstra (1981a,b) argued that hyper/hypostotic traits are produced by ossifications of connective tissue that are necessarily less variable than what is seen in the formation of foramina, largely because foraminal traits have several developmental sources. Therefore, the variation in composite foraminal traits could be lower than the variation in the parts that compose them, producing lower estimates of genetic variability than is really there. In addition, some traits previously considered to be epigenetic are now being linked to various kinds of stress. Mayhall andMayhall (1971), for instance, think that diet, via masticatory stress, affects the development of the mandibular torus more than genetics. Benfer and McKern (1966) have shown that olecranon perforation in the humerus is related to both the robusticity of the bone in question and stress on the fossa. Further study is not likely to simplify the question of relative heritability. Mathematically speaking, as Jantz (1970) points out, we already know that the expression 32 of epigenetic traits is affected by different processes in metric vs. nonmetric traits. For discrete traits, incidence is a function of both the mean and the variance of the underlying genetic variable. Why? Because an individual’s genotypic distance from the mean (in standard deviations) determines his or her possession of the trait. Therefore, a change in the mean or the variance can affect the appearance of a discrete trait, but only a change in the mean will affect metric traits. Furthermore, as Corruccini (1974) pointed out, different modes of natural selection have different effects on the distributions underlying the phenotype. Directional selection changes the mean. Stabilizing selection changes the variance. Drift and hybridization can change both the mean and the variance. Livingstone (1972) notes that heritability differences also arise between polygenic traits and single gene (non-additive) traits, depending on which selection pressures are in operation. Single gene traits and partially additive traits react faster to selection pressures. Polygenic traits react more slowly, but their variance can change even faster than that of single gene traits. Single-gene traits can become more variable via hybridization and/or stabilizing selection pressures, while measurements (which are polygenic) usually do not. This is because random factors tend to affect metric traits in both directions, so that their effects cancel out. Not so for discrete traits, which are more susceptible to genetic drift. In some isolates, metric traits will be more variable because their distribution curves are flattened by the predominance of two homozygous types, while nonmetric variability is reduced because the chance of fixation is steadily increased as fewer and 33 fewer loci are involved (Corruccini 1974). These factors may explain why, for example, Bennett and Hulse (1966) found very little metric difference between skulls from Mesa Verde and those of their neighbors, but did find significant differences with nonmetric traits. This may be an important consideration in analyzing differences between the Meganos and their nearest probable relatives, especially those that are separated by relatively short periods of time. In any case, heritability measures are specific to populations and cannot be used to generalize beyond those populations (Saunders 1989). This is because a change in environmental variability or genetic variability can change the heritability of a trait without changing the mean incidence of the trait in a given population. Similarity and Kinship Berry and Berry (1967) held that nonmetric traits vary even between closely related populations. It must be remembered, however, that similarity does not equal relatedness. Groups that are dissimilar can be assumed to be different, but groups that are similar may be undergoing convergent selection pressures, genetic drift, and other random processes (Relethford and Blangero 1990, Saunders 1989). Environmental Variability Berry and Berry (1967) thought that discrete traits exhibit constancy under environmental variability. Given the state of knowledge in 1967, perhaps it should have been obvious that this was not so. As Corruccini (1974) points out, bone growth and development, in particular, can be strongly affected by environmental variants and 34 nutritional levels. Also, some developmental problems can produce discrete traits that are largely unrelated to genetics. For example, hydrocephaly produces large numbers of sutural ossicles (Hertzog 1968). Intentional cranial deformation also produces higher frequencies of Wormian bones (Ossenberg 1970). Cradleboarding, in particular, results in increased numbers of lambdic, occipito-mastoid and asterionic ossicles, all at points closest to those where pressure was applied to the skull. At the same time, there is a decrease in the frequency of Wormian bones in the lateral vault sutures, where the skull was free to expand, e.g., the parietal notch bone, epipteric bone and coronal ossicles. Both phenomena are more pronounced in females than in males, and according to Ossenberg (1970), the total incidence of posterior Wormians is three times the incidence of lateral Wormians in the deformed, but very nearly one to one in the undeformed. Some specific differences included: (1) facial variations – the os japonicum and infraorbital suture were found less often in the deformed skulls, but only in those of males; and (2) frontal bone variations – the supraorbital foramen and frontal grooves are all seen less in deformed skulls, but again only in the skulls of males. All these effects are of interest in analyzing the discrete traits of crania from California populations because some Penutian groups are known to have practiced cranial deformation. One such is the Modoc, a northeastern California people closely related to the Klamath of Oregon, where the practice was common throughout the Klamath Lakes area as well as the Columbia River Basin during pre- and early historic times (Tasa 1986). 35 Age Berry and Berry (1967) stated that discrete traits do not vary with age. However, Corruccini’s (1974) study of 321 adult skulls from the Terry Collection showed that nonmetric traits actually vary with age much more than most metric traits do. As Gruneberg (1963:267) pointed out, most of the minor skeletal variants are the outcome of conflicts between several agencies and “whatever influences size will indirectly tend to affect the incidence of some of the minor variants.” This means growth, progressive ossification and differential development will all have their effects on both types of traits. The variation that does occur is not unidirectional either. When Ossenberg (1970) looked at deformed versus normal skulls, she found a pattern of age-linked regression for hypostotic variants and of progression for hyperostotic variants. This is a major problem, since age estimation in archaeological samples is even less reliable than sex determination, especially for older subjects. To date, no substantial investigation of advanced age effects on nonmetric variations has been done. Therefore, Corruccini (1974) suggests that the only solution is to use Spearman’s rank-order gradation techniques and separate subjects by age groups prior to statistical analysis. Sex Berry and Berry (1967) stated that nonmetric traits show no sex differences. Their pooled data showed no significant differences between pooled and sex-classed data. However, their study looked at only one population. Larger scale studies like that of Corruccini (1974) clearly show that discrete traits do show sex-linked differences in frequency, but at about half the level seen in metric traits. 36 Others, like Jantz (1970), have also found strong sex links in their data, and some had done so long before Berry and Berry (1967) came along. For example, Schultz (1923) had shown some 44 years earlier that the frequency of bregmatic ossicles shows a strong sex link. The same is true of metopism (Comas 1942). Jantz (1970) simply removed dimorphic traits from his analysis in order to take care of this problem, but Corruccini (1974) considers this a bad idea because the most dimorphic traits also tend to be the most valuable in separating populations. Further problems have arisen in studies like that of Lees and Relethford (1982), who looked at marital residence patterns and found greater variation among nonmigrant females than nonmigrant males. The authors suggested that environmental factors must have produced the difference in variability, and pointed to the higher canalization of female development. However, Hall and MacNair (1972) found greater male than female intra-group variability in a similar study of British Columbian Amerindians. They suggest that the differences between the sexes have an environmental basis, and the phenotypes of females are better buffered against nutritional fluctuation. According to Konigsberg (1988), it does not really matter which of these ideas is correct because there is no choice to be made with archaeological samples – you have to assume that there were no differential environmental effects just because there is no good way to detect or quantify the difference. The effects are also linked to Ossenberg’s (1970) hyperostotic vs. hypostotic split in age-progression, and there remains the basic problem of sex determination. The best anthropologists working today can only claim an accuracy rate of 80% in sexing bones, and Weiss (1972) has shown a persistent 12% bias 37 toward males throughout skeletal studies, so sex-linked data will always be the fuzzier for it. Independence Are nonmetric traits really independent of each other? As noted above, Berry and Berry (1967) only tested this in one population. Others who agreed with their assessment, like Comas (1942) and Sullivan (1922), only tested a few associations in each of their studies. A number of other studies, however, have turned up lots of traits that are the result of the same developmental pathways, and do show correlations in incidence. For example, Carpenter (1976) compared metric and nonmetric traits and found the following three correlations: 1. minimum frontal breadth and metopism (also reported by AshleyMontagu 1937); 2. nasal breadth and nasal sill sharpness; and 3. height of the ascending ramus and nasal sill sharpness. Bennett (1965) reported a correlation between basi-occipital length and lambdoidal Wormian bones. Comas (1942) reported another between metopism and epipteric bones, and Woo (1949) reported one between metopism and frontal curvature. Others discrete traits did turn out to be independent. Comas (1942) found that biasteric breadth was not tied to metopism. Benfer (1970) compared Asian, European, Amerindian, African and Indian samples and found that circumparietal accessory ossicles, parietal foramina, and form of the fronto-temporal suture are all largely independent of each other, both within and across geographic races. Hertzog (1968) 38 picked out a single bone and tested all possible associations between traits located along the periphery of the parietal bone, finding the frontal area to be independent of the occipital area. Saunders (1989) and Corruccini (1974) both state, however, that every population has to be tested for these kinds of correlations, because the significance of the correlations varies in different groups. Scoring Berry and Berry (1967) found discrete traits easy to score, but many workers have serious problems in defining nonmetric traits, and in scoring them consistently, particularly those that must be scored by grades rather than simple absence/presence (Corruccini 1974). Fossae, tubercles and tori are continuous in development and seem to produce the same problems. Buikstra (1972) provides a mathematical method for correcting for the problem of partial trait manifestation but Corruccini (1974) doubts its usefulness. Deformation Some workers, like Sullivan (1922), have promoted the use of discrete traits in cases where artificial cranial deformation limits the use of metrics. However, Ossenberg (1970) has shown that cranial deformation also affects nonmetric traits, and not always in the same way. In looking at 1200 undeformed skulls from Arctic sites, Woodland sites, and 19th Century Dakota Sioux, Ossenberg (1970) found two patterns: 1. For hypostotic traits, such as metopism and tympanic dehiscence, incidence is age-regressive till growth ends, and then becomes stable. 39 Ossenberg saw this as a matter of arrested morphogenic development, and found it slightly more common in females and on the right side of the cranium. 2. For hyperostotic traits, such as auditory exostoses, clinoid bridging, pterygo-spinous bridge, and the paracondylar process, the pattern is age progressive. Ossenberg saw this as a matter of excessive ossification and found it slightly more common in males and on the left side of the cranium. According to Torgerson (1951), the right/left differences are probably related to richer ennervation on the right side, associated with cerebral hemisphere dominance and greater blood supply. As Gruneberg (1963:267) put it, “…if one side of the body is ahead of the other in development, asymmetrical manifestations may be the result.” It also serves to further complicate the question of how to score two-sided traits. Unilateral Expression of Bilateral Traits Scoring of some discrete traits is also complicated by variations in their bilateral expression. While some traits, such as the presence of the os bregma, can occur at only one site, many others may occur on one or both sides of the head, e.g., coronal ossicles. Some researchers respond by using scores recorded from the left side only (Corruccini 1974). Others recommend random selection from both the right and left sides (Konigsberg 1990). A recent study of the unilateral expression of bilateral traits like these was conducted by Hallgrimsson, Ó Donnabháin, Blom, Lozada and Willmore (2005), and showed that unilateral expression is linked to trait frequency. Rarer traits are 40 more likely to be expressed on one side only, while more common traits tend to be expressed bilaterally. Hallgrimsson et al. (2005) concluded that this trend is probably the result of small differences in trait formation thresholds on each side of the head. The number of individuals in which one side, but not the other, is pushed over the threshold for trait formation will in general be a larger proportion of the total frequency of expression when that trait’s frequency is low. As trait frequency increases, variation within individuals becomes less significant as a determinant of trait expression in comparison to variance within a population. This confirms an earlier study by Ossenberg (1981) which reported the same relationship between trait frequency and unilateral expression of two mandibular traits. Fragments Another advantage to using nonmetric traits, according to Saunders (1989), is that they can be scored on fragments when metrics cannot. This advantage may be less important now that methods are available for estimating the length of incomplete long bones (Steele and McKern 1969, Wright and Vasquez 2003). However, there are many situations in the archaeological record where cranial fragments are all that is available. The Great Debate A number of researchers have now completed studies in which both metric and discrete traits were used to measure biological distance. In many cases, metric traits provided more clear-cut results than nonmetric traits. 41 For example, Zegura (1975) looked at metric and nonmetric traits in 12 Eskimoid populations. In addition to gender-pooled groups, he subdivided his subjects into male and female groups and examined the effects of sex on his overall results. In general, Zegura found smaller differences between the sexes when using metric traits than while using discrete traits. The metric results were also a better fit than the nonmetric results for the data on linguistic relationships among his populations. But Zegura used 74 metric traits, and only 28 nonmetric traits, admitting that only 15 of the nonmetric traits may really be independent of each other. Rightmire (1972) used both types of trait to look at African skulls. He also found that the metric results fit much better than nonmetric data. However, Rightmire used more than ten times as many metric traits as he did discrete, and did not test even those few discrete traits for true independence. Also, as Corruccini (1974) mentions, there may be a limit to the heterogeneity of nonmetric traits, meaning that if you exceed this limit by comparing truly unrelated groups, your results are going to be off the scale and essentially meaningless. Carpenter’s (1976) study also used both types of traits. He looked at 317 crania from the Terry Collection, using 12 metric traits and 15 nonmetric traits. He found that metrics were better at discriminating sex and race, while nonmetrics were better at parsing age classes. Nonmetrics, he thinks, should only be used to supplement metric data. This is not too surprising given his choice of discrete traits, which included metopism, mandibular and palatine tori – all of which are known-to be sex-linked to 42 some degree. Another trait, sharpness of the nasal spine, is notoriously hard to score with any consistency (Corruccini 1974). A number of other researchers found that nonmetrics worked better for their studies. For example, Ossenberg (1977) looked at one Aleut and four Eskimo populations. She used sex-pooled data but selected discrete traits that are not strongly dimorphic (according to Corruccini 1974). She also chose traits that have been tested for independence by a number of other researchers (e.g., Comas 1942, Benfer 1970, Corruccini 1974). As a result, Ossenberg found that nonmetric traits showed a stronger concordance with linguistic/geographical data than metrics did. Another study, by Steadman (2001), looked at problems related to cranial deformation. She found, as have many others, that metrics can be skewed by even slight deformation, which can be hard to detect. She found that 53% of the 487 crania she looked at showed natural flattening that could not be distinguished from 231 that showed definite artificial deformation. Steadman (2001) also found high rates of inter- and intraobserver errors in detecting slight deformation, leading to her conclusion that nonmetric traits, if carefully selected, were clearly more reliable. Summary Corruccini’s (1974) review seems to sum it up pretty well. He looked for correlations between metric and nonmetric traits by examining 321 adult skulls from the Terry Collection. For this purpose, he scored 72 discrete traits, later reduced to 61 nonmetric traits, and 23 metric traits as well. He then used Pearson’s correlation coefficient to test the data for correlations of individual traits with sexual dimorphism, 43 age dependence, racial grouping, intercorrelation with other nonmetric traits and correspondence between nonmetric and metric analyses of biological distance. His results offer the clearest quantification of the differences between metric and nonmetric traits. In Corruccini’s (1974) study, the strongest correlation for both metric and discrete traits was with sex, but within this pattern, he found significant differences between racial groups. He found 19 of 61 discrete traits differed significantly between male and female Whites, while only three should have differed by chance. He found only nine traits differed significantly between male and female Blacks, where again only three should have differed by chance, and they were different sets of traits. White sex differences were located in the frontal grooves, palatine torus and accessory mylohyoid foramen, while Black sex differences were strongest in the mastoid foramen and bifaceted condyles. The level of dimorphism, then, was different between the two racial groups, and it was based on different parts of the cranium. Corruccini’s (1974) second strongest correlation for both types of variable was with age class. Overall, he found that nonmetric traits only produced about half as much difference between the sexes as metric traits, while age affected discrete traits several times as much as it did metric traits. Overall, about four times as much overall sample separation was afforded by 23 metric traits as by 30 nonmetric traits. Corruccini (1974) recommends treating nonmetric analyses in the same way as metric analyses, with separation of sex and age classes. If handled in this way, he says that metric and nonmetric studies should “behave in the same way.” This seems to be 44 true. There have now been several studies making use of both types of trait, and the majority obtained much the same results from each type. For example, Tatarek and Sciulli (2000) used nonmetrics to test metric results from an earlier study on hunter/gatherers in the Ohio Valley versus early horticulturists, and got the same results. The conclusion seems clear. Analysis of metric traits will usually provide a more precise definition of the biological distance separating sample populations than will analysis of discrete traits, the major exception being studies of very closely related groups in similar environments. However, if nonmetric traits are chosen with care, and tested for independence, and if the same analytical standards are applied as would be used for metric traits, then the outcome will probably be the same. 45 Part 4. Research Strategy The purpose of this study is to examine the biological distance between populations whose skeletal remains have been recovered from six sites spanning California’s Central Valley, the San Francisco Bay and Delta and the Santa Clara Valley (see Table 2). A comparative analysis of metric and nonmetric (discrete) cranial traits seen in these samples may help define the nature and extent of prehistoric population movements associated with the so-called Meganos Intrusion hypothesized by Bennyhoff (1986), and with peaks in violent trauma noted by other researchers (e.g., Jurmain 2001). Table 2. Sample Sites/Components Used in This Study Sample Site CA-SJO-091 CA-SAC-117 CA-SJO-105 CA-ALA-343e CA-ALA-343f CA-SCL-674 CA-SCL-137 Chronology Early Middle Period Late Period Late Period/ Early Historic Upper Middle Period Upper Middle Period Upper Middle to Lower Middle Period Early Middle to Lower Middle Period Approximate Dates 200-900 BC Burial Type Extended Tribal Affiliation Proto-Miwok AD 700-1700 Extended AD 700-1850 Extended AD 500-700 Extended Northern Miwok Northern Valley Yokuts Meganos? 500 BC to AD 1000 200 BC to AD 900 680 BC to AD 300 Flexed Unknown Flexed Unknown, possibly Esselen Unknown, possibly Esselen Flexed Of the six sites chosen for this analysis, the site of central interest is an Upper Middle Period site (500 BC - AD 1000) from the South Bay – CA-ALA-343. This site 46 consists of a large cemetery with a number of burials demonstrating Meganos cultural traits, and others clearly demonstrating Berkeley pattern traits. Regrettably, two other sites generally considered ‘type’ sites for the Meganos aspect, CA-CCO-139 and CACCO-141, could not be used for lack of craniometric data. Although such data is mentioned in site records stored at the Northwest Information Center at California State University, Sonoma, the Phoebe Hearst Museum was unable to locate any osteological information for either site. The nonmetric data available for either site were scored on materials from CA-CCO-139 by Suchey (1975). Two Middle Period sites in which the Meganos Aspect is absent and the Berkeley Pattern is present are located in the Santa Clara Valley: CA-SCL-674 and CA-SCL-137. Sites in the Central Valley which were included in this study are CA-SJO-91, CA-SAC-117, and CA-SJO-105. These three sites are assigned to groups that Bennyhoff considered either ancestral to or descended from the Meganos people. If the Meganos Intrusion described by Bennyhoff and Fredrickson (1994) did occur, then remains from the extended burials found at CA-ALA-343 will demonstrate less biological distance from those found in similar burials at proto-Miwok sites in the Central Valley and more from those at the two non-Meganos (and possibly protoEsselen) sites in the Santa Clara Valley. A craniometric analysis comparing all six sites and various subgroups of these sites was carried out to determine the biological distance between these prehistoric peoples and determine whether such measures of biological distance support or refute Bennyhoff’s (1986) hypothesis. However, some of the skeletal materials recovered at 47 these sites are heavily damaged and fragmented, due to a number of taphonomic processes. The number of specimens suitable for a craniometric approach is, therefore, quite limited. Because of this constraint and because some of these groups may indeed be very closely related, a concomitant analysis of nonmetric traits was carried out as a further test of the biological distance between these groups. Analysis of both types of traits also allows an assessment of the relative merits of using metric versus discrete traits in studies of this type, and provides a foundation for related studies of regional population movements in prehistoric California. 48 Chapter III MATERIALS In order to test Bennyhoff’s (1986) hypothesis regarding the population movements he called the Meganos Intrusion, this study will examine crania from burials excavated at relevant sites. According to Bennyhoff (1986), a Penutian group exhibiting a modified Windmiller Pattern in its material culture moved into the East and South Bay regions of San Francisco from the Central Valley in California shortly after the onset of the Early Middle Period (ca. 500 BC to AD 100). This group displaced Hokan-speaking groups and others living in the Bay and Delta, including ancestors of the Esselen and possibly the Costanoan peoples, and then retreated into the San Joaquin Delta at the end of the Upper Middle Period (ca. AD 700-900). Therefore, remains from extended burials identified as exhibiting the Meganos Aspect at CA-ALA-343 will be compared to those from flexed burials at the same site that demonstrate Berkeley pattern traits, and with those from two contemporaneous non-Meganos sites (CA-SCL-137 and CA-SCL-674). These groups will also be compared with three Central Valley sites (CA-SAC-117, CASJO-91 and CA-SJO-105). Of these Central Valley sites, one (CA-SJO-91) is an Early to Lower Middle Period site possibly linked to the ancestors of the Meganos people. The other two (CA-SAC-117 and CA-SJO-105) are Upper Middle to Late Period Phase II sites linked to possible descendants of the Meganos people. A map of the sites included in this study is shown in Figure 7. Metric and discrete traits from suitable crania will be 49 analyzed separately to determine the relative level of biological distance between these populations. Site Descriptions 1. CA-SJO-91 is the earliest Central Valley site, and dates to the Early Middle Period at 900-200 B.C., with carbon-dating of material from one grave resulting in a date of 1834 B.P. Excavation in 1954-55 was carried out under the supervision of Dr. Richard Reeve and yielded a total of 99 burials. Artifact association links this site to the Windmiller Pattern of material culture, and should, therefore, be associated with groups ancestral to the Meganos people if Bennyhoff’s (1986) hypothesis is correct. No information is available concerning which standards were used for osteological analysis, but the data sheets are consistent with the standards proposed by Howells (1973). 2. CA-SAC-117 is a Late Period site (700–1700 A.D.) occupied by the Plains Miwok. The site is located on the eastern shore of the Cosumnes River in the San Joaquin Valley. Test excavations were carried out in the early 1950s under the direction of Dr. Richard Reeve when the site was uncovered by leveling due to agricultural activities. No information is available concerning which standards were used for osteological analysis, but the data sheets are consistent with the standards proposed by Howells (1973). 3. CA-SJO-105 is a Late Period/Early Historic site (700–1850 A.D.) whose residence time frame ended within the last 250 years. The site is located just north of Stockton on Five Mile Slough (Vittitow 1999). This rich riverine setting is in a transition zone between the Delta and the broad plains of the San Joaquin Valley and may have been used by the ancestors of the Northern Yokuts (Vittitow 1999). However, the site 50 Figure 7. Location of archaeological sites to be used in current study. Blue site numbers refer to Central Valley sites possibly related to the Meganos peoples. The red site number refers to the possible Meganos site. Green site numbers refer to nonMeganos sites in the Santa Clara Valley. 51 may also have been used by the Plains Miwok, a closely related Penutian group. Both are possible descendants of the Meganos People. Excavation was carried out by Dr. Richard Reeve in 1953-1954 prior to construction of a housing subdivision. No information is available concerning which standards were used for osteological analysis, but the data sheets are consistent with the standards proposed by Howells (1973). 4. CA-ALA-343 is a Middle Period (500 BC – AD 1000) site with at least two cultural components in evidence, including the Meganos Aspect. The site was registered by T.F. King in 1968. He reported a midden that had been scattered over a wide area due to farming activities, and stated that human remains were present. Although several testing programs and excavations have been carried out on the site, no comprehensive report has yet been written. Existing documentation includes a preliminary report by Wildesen (1968) on the San Francisco State College excavation in 1968, two reports on a testing program carried out by Archaeological Resource Services in 1981 and 1994 (Roop 1997), preliminary reports on testing and burial excavation performed by San Jose State University in 1985 and 1988 (Hall 1985, and Hall, Jurmain and Nelson 1988), and a report on the results of a testing program done by Holman and Associates in 1990. Other unpublished data includes excavation and burial data from the 1988 SJSU excavation (Hall 1988 unpublished notes) and an unpublished analysis of the burials excavated in 2000 by Archaeor (Marshall 2002). Three excavations at CA-ALA-343 have revealed nearly 400 burials. The data used for this study came from the latest, carried out by Archaeor in 2000, in which 270 burials were excavated, and the remains of 224 individuals were analyzed for osteological 52 characteristics. Two cultural components were present, one characterized by extended burial positions and Meganos-type grave goods, and the other characterized by flexed burial positions and Berkeley Pattern artifacts. Craniometric and nonmetric data were collected according to standards published by Buikstra and Ubelaker (1994). 5. CA-SCL-674 is a contemporaneous site in the Hillsdale region of San Jose, in close proximity to both Canoas Creek and the Guadalupe River. Radiocarbon dating and artifact association dates the site to a time frame beginning in the Early/Middle Transition Period, 600 – 100 B.C., and extending into the Middle and Late Periods. The grave goods exhibited the Berkeley Pattern rather than the Meganos, including a tightly flexed burial position. Excavation yielded 218 burials (Grady, Latham and Andruschko 1999). Craniometric data were collected from the remains according to standards published by Breschini and Haversat (1980). Nonmetric data were collected according to standards published by Berry and Berry (1967). 6. CA-SCL-137 is also a Middle Period site excavated after several burials were uncovered during construction of a pipeline underneath Snell Avenue in South San Jose (King, Delgado and Roehr 1974). Radiocarbon dating, obsidian hydration studies and artifact association date the site to a time frame beginning in the Early Middle Period, ca. 680 B.C, and approaching the Lower Middle Period, which began ca. 300 A.D. Like CA-SCL-674, grave goods from this site demonstrated the Berkeley Pattern and the Meganos Aspect was absent. Excavation yielded 22 burials (Cartier, Sanfilippo, Van Zandt, Carrico and Laffey 1987). Craniometric data were collected according to 53 standards published by Bass (1971). Nonmetric data were scored according to standards published by Howells (1973). The sample sizes for the raw data collected from these six sites are shown below in Table 3. Table 3. Raw Data Sample Sizes Sample Site CA-SJO-091 (ext. burials) CA-SAC-117 (ext. burials) CA-SJO-105 (ext. burials) CA-ALA-343e (ext. burials) CA-ALA-343f (flexed burials) CA-SCL-674 (flexed burials) CA-SCL-137 (flexed burials) Metric Traits (# individuals) 24 22 25 26 266 57 15 Nonmetric Traits (# individuals) 23 20 28 25 120 52 30 Data Collection The data sets for the three Central Valley sites (CA-SJO-091, CA-SAC-117, and CA-SJO-105) were obtained by reference to the original burial sheets and site records maintained by California State University, Sacramento in the collections of the Anthropology Department, all three sets having been published previously as part of other studies. The data set for the Alameda site, CA-ALA-343, was obtained from the offices of Archaeor in Fremont, CA. The data set for the Rubino site, CA-SCL-674, was published in conjunction with the site report (Grady et al. 1999). The data set for the South San Jose site, CA-SCL-137, was obtained from unpublished materials housed at the Northwest Information Center in Sonoma, CA. In some cases, direct access to the skeletal materials is no longer possible because they have been repatriated and/or 54 reburied according to the provisions of the Native American Graves Protection and Repatriation Act under Public Laws 101-185 and 101-601. For that reason, a metaanalysis based on the data collected by other researchers was required in order to confirm or refute Bennyhoff’s (1886) hypothesis concerning the Meganos Intrusion. As noted previously, the data used in this meta-analysis were obtained by several researchers, using several sets of standards. This multitude of standards made it difficult to compare the data directly from site to site and required the elimination of data for a number of traits, both metric and nonmetric, which had not been collected at all six sites. In general, craniometric data collected prior to 1980 were obtained using the standards set forth by Bass (1971) or by Howells (1973), while some later analyses were carried out using the revised standards set forth by Breschini and Haversat (1980). Data collected after 1999 were obtained using the standards set forth by Buikstra and Ubelaker (1994). Precise definitions of the traits selected for this study are given below, to avoid confusion. Data on nonmetric traits were collected according to standards set forth by Berry and Berry (1967), or by Howells (1973) or by Buikstra and Ubelaker (1994). For three Central Valley sites excavated by Dr. Richard Reeve in the 1950’s, no information is available on which standards were used for osteological analysis, but the data collection sheets appear to be fairly consistent with standards later set forth by Bass (1971) and Howells (1973). Precise definitions of the metric traits selected for this study are also given below. 55 Data from both males and females were used for this analysis, while data from individuals of indeterminate gender, and data from subadults, were omitted. Data from burials demonstrating extended burial positions at CA-ALA-343 were assigned to a test case denoted ‘CA-ALA-343e’ and constitute the ‘Meganos’ sample, while data from burials demonstrating flexed burial positions were assigned to a test case denoted ‘CAALA-343f’, and constitute a ‘non-Meganos’ sample. Burials at this site demonstrating indeterminate burial positions were omitted from the analysis. Definitions Metric Measurements Metric measurements used in this study included the fourteen items listed and defined below. Illustrations delimiting these measurements are shown in Figures 8, 9, 10, 11 and 12. Cranial measurements (as defined by Buikstra and Ubelaker 1994): 1. Maximal Cranial Length: the distance between the glabella and opisthocranion in the midsagittal plane, measured in a straight line (Figure 8). 2. Maximal Cranial Breadth: maximum width of the skull perpendicular to the midsagittal plane (Figure 9). 3. Bizygomatic Diameter: direct distance between most lateral points on the zygomatic arches (Figure 9). 56 Figure 8. Cranial measurements in the sagittal plane. 57 Figure 9. Cranial measurements, frontal view. 58 Figure10. Cranial measurements, frontal view. 59 4. Cranial Base Length: direct distance from nasion to basion (Figure 8). 5. Upper Facial Height: direct distance from nasion to prosthion (Figure 10). 6.. Minimum Frontal Breadth: direct distance between the two frontotemporale (Figure 10). 7. Nasal Height: direct distance from nasion to the midpoint of a line connecting the lowest points of the inferior margin of the nasal notches (Figure 10). 8. Nasal Breadth: maximum breadth of the nasal aperture (Figure 10). Mandibular Measurements: 9. Chin Height: direct distance from infradentale to gnathion (Figure 11). 10. Bigonial Width: direct distance between right and left gonion (Figure 11). 11. Bicondylar Breadth: direct distance between the most lateral points on the two condyles (Figure 11). 12. Minimum Ramus Breadth: least breadth of the mandibular ramus measured perpendicular to the height of the ramus (Figure 12). 13. Maximum Ramus Height: direct distance from the highest point on the mandibular condyle to gonion (Figure 12). 60 Figure 12. Measurements of the mandible, lateral view (after Buikstra & Ubelaker 1994). 61 14. Mandibular Length: distance of the anterior margin of the chin from a center point on the projected straight line placed along the posterior border of the two mandibular angles (Figure 12). Nonmetric Traits In the standards set forth by Buikstra and Ubelaker (1994), the authors recommend 30 discrete traits as being the most useful in separating aboriginal populations in North America. However, because the nonmetric data sets to be used in this analysis were collected under several standards, including Berry and Berry (1967), Howells (1973), Finnegan and Cooprider (1978) and Buikstra and Ubelaker (1994), the traits that will be used for this analysis are limited to 14 traits that are common to all three sets of standards, and were recorded for all six sites. In addition, as noted in their definitions, the scoring of graded traits was simplified to the simple absence or presence of the trait in question. Data recorded under earlier standards can then be directly converted to binary data. Foramina and Canals 1. Condylar Canal: canal opening within the condylar fossa, posterior to the occipital condyles. An open or ‘patent’ canal is directed antero-medially, ending in a foramen located on the superior aspect of the pars lateralis, within the cranial cavity, or within the jugular notch (Figure 15). Scoring: 0 = not patent 1 = patent 9 = unobservable 62 Figure 13. Frontal view of cranial vault showing locations of metopic suture, supraorbital foramina, infraorbital foramina, and zygomatico-facial foramina. 63 Figure 14. Superior and lateral aspect of cranial vault showing locations of parietal foramina, sutural bones and mastoid foramen 64 Figure 15. Posterior and basilar aspects of the cranial vault, showing the location of the condylar canal, and lesser accessory palatine foramina 65 . Supraorbital Foramina: notches and/or foramina may be present at the supraorbital margin of the frontal bone. All foramina must present openings on both orbital and external surfaces to be scored as present. Foramina on the superior, horizontal aspects of the margin are considered supraorbital, while those located at the confluence of the vertical (medial) and horizontal aspects of the orbit are considered supratrochlear. Only supraorbital foramina will be scored. Multiple supraorbital foramina will be scored the same as singular foramina. Supraorbital notches will be scored as present when more than half occluded by spicules, and as absent when less than half occluded (Figure 13). Scoring: 3. 0 = absent 1 = present 9 = unobservable Presence of Infraorbital Foramina: foramina situated on the external anterior surface of the maxilla below the infraorbital margin, above the canine fossa, and extending into a canal that opens on the orbital floor. Divisions observable within the infraorbital canal will be scored the same as true multiple foramina, and multiple foramina will be scored the same as singular accessory infraorbital foramina (Figure 13). Scoring: 4. 0 = absent 1 = present 9 = unobservable Presence of Parietal Foramen: located on the parietal bone, within or near the suture at obelion. Foramen should lead into canal extending to and 66 perforating the internal table Foramina existing only on the internal table will not be scored. Foramina located on the parietal will be scored the same as sutural foramina (Figure 14). Scoring: 5. 0 = absent 1 = present 9 = unobservable Mastoid Foramen: located posterior to the mastoid process, usually on the temporal bone, but occasionally on the occipital or within the occipitomastoid suture. Under Buikstra and Ubelaker (994), both location and number would be reported. For the purposes of this study, simple presence/absence will be recorded, with multiple foramina scored the same as singular foramina. Location with regard to a sutural vs. exsutural position (Figure 14) will not be used. Scoring: 6. 0 = absent 1 = present 9 = unobservable Presence of Accessory Lesser Palatine Foramen: small foramina located posterior to the palatine foramina, on the horizontal processes of the palatine bones. Multiple accessory foramina will be scored the same as singular foramina (Figure 15). Scoring: 0 = absent 1 = present 0 = unobservable 67 7. Zygomaticofacial Foramina: foramina located on facial surface of zygomatic (malar) bone. Size will not be scored. Multiple foramina will be scored the same as singular foramina (Figure 13). Scoring: 0 = absent 1 = present 9 = unobservable Ossicles Scoring for all: 8. 0 = absent 1 = present 9 = unobservable Presence of ossicle at Lambda: located within lambdoid suture; also known as the apical bone (Figure 14). 9. Presence of ossicle(s) in the lambdoid suture on either side of Lambda (Figure 14). 10. Presence of Parietal Notch Bone: ossicle located within the parietal notch, between the squamous portions of the temporal and the parietal bones (Figure 14). 11. Presence of Bregmatic Bone: located at the junction of the paired parietals and the frontal, formed within the anterior fontanelle (Figure 14). 12. Presence of ossicle(s) in the coronal suture: located within the coronal suture (Figure 14). 13. Presence of Asterionic Bone: located at the juncture of the occipital, parietal and the temporal bones (Figure 14). 68 Sutures 14. Presence of Metopic Suture: located on the midline of the frontal bone, from bregma to nasion. The metopic suture usually closes by the eighth year, but may be completely or partially retained in adults. Partial sutures will be scored the same as complete sutures (Figure 13). Scoring: 0 = absent 1 = present 9 = unobservable