trypanosomoses_complete
advertisement

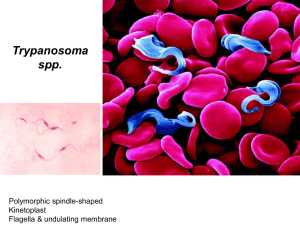
Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Trypanosomoses Author: Vincent Delespaux Adapted from: R.J. CONNOR and P. VAN DEN BOSSCHE, 2004, African animal trypanosomoses, in Infectious diseases of livestock, edited by J.A.W. Coetzer & R.C. Tustin. Oxford University Press, Cape Town, 12: 251 – 295 Licensed under a Creative Commons Attribution license. TABLE OF CONTENTS Introduction ................................................................................................................... 2 Epidemiology................................................................................................................. 4 Pathogenesis ............................................................................................................... 12 Diagnosis and differential diagnosis ......................................................................... 15 Diagnosis and differential diagnosis .....................................................................................15 Clinical signs and pathology ........................................................................................15 Laboratory confirmation ...............................................................................................21 Differential diagnosis ............................................................................................................22 Control / Prevention .................................................................................................... 23 FAQs............................................................................................................................. 26 References ................................................................................................................... 27 1|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses INTRODUCTION The trypanosomoses are diseases of humans and domestic animals that result from infection with parasitic protozoa of the genus Trypanosoma. Trypanosomes parasitize all classes of vertebrates: fish, amphibians, reptiles, birds and mammals. The parasites, with the exception of Trypanosoma equiperdum, the cause of dourine, are transmitted from host to host by haematophagous vectors, and usually cause little appreciable harm to either the vector or the vertebrate host. However, several species of trypanosomes which parasitize mammals are less well adapted and commonly cause disease. Trypanosomosis is generally characterized by the intermittent presence of parasites in the blood and intermittent fever. Anaemia usually develops in affected animals, and this is followed by loss of body condition, reduced productivity and, often, high mortality. The first report that associated trypanosomes with disease was made from India in 1880. In 1895 a major discovery was made in Zululand, South Africa that trypanosomes were the causal organisms of ‘nagana’, or tsetse fly disease. Two forms of human trypanosomosis exist: Chagas' disease occurs in Central and South America and is transmitted by bloodsucking reduviid bugs, certain small wild animals and dogs harbouring the infection. The second form is human sleeping sickness. This occurs in Africa and is transmitted by bloodsucking flies of the genus Glossina, commonly known as ‘tsetse flies’ or simply as ‘tsetse’. The majority of animal diseases caused by trypanosomes occur in the tropics. In Africa, several species of tsetse-transmitted trypanosomes cause African trypanosomoses in domestic animals, which in southern Africa are collectively known as ‘nagana’, a word derived from the Zulu word ‘nakane’ meaning tsetse fly disease. ‘Surra’ is transmitted by biting flies other than tsetse flies and, although it occurs in many parts of the tropics, including northern Africa, it is not present in southern Africa. The large populations of wild animals, which have thrived for millennia in the tsetse-infested tracts of Africa have evolved with these flies and the trypanosomes they transmit. Hosts and parasites have become mutually adapted and co-exist in a balanced relationship. Humans first brought domestic animals into the tsetse belts of Africa relatively recently. Because of this recent introduction, the relationship between tsetse-transmitted trypanosomes and domestic animals has not fully evolved and infection with these parasites frequently produces disease. The devastation which resulted from the rinderpest pandemic of the 1890s destroyed almost entire populations of wild animals and millions of cattle. Without hosts on which to feed, tsetse disappeared from large areas. However, a few decades later, tsetse were dispersing from residual pockets, and trypanosomosis again became a problem for livestock owners. By 1931, tsetse were spreading at a rate of 2500 square kilometres (1000 square miles) annually, and game elimination to control tsetse began in 1932. Since then strenuous efforts have been made to contain the tsetse fly. In many other parts of southern Africa, livestock owners have also had to live with the tsetse fly and its consequences. 2|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Tsetse infest 10 million square kilometres and affect 37 countries, which makes African animal trypanosomosis a problem of truly continental magnitude. They live in frost-free areas that have an annual rainfall of 650 mm or more. In arid, marginal habitats, tsetse only exist in the better wooded and better watered strips where the host species concentrate during critical times, such as in the late, hot, dry season. Most of the settled areas of the tsetse fly belts of southern Africa are used for traditional mixed farming, but the presence of tsetse seriously handicaps development. Tsetse fly Cattle Trypanotolerant cattle General distribution of tsetse flies and cattle in Africa Concerted efforts to control tsetse over the past 50 years have resulted in significant changes in the distribution of tsetse and tsetse-transmitted trypanosomosis. Unfortunately, few of these achievements have been sustained. In many countries of southern Africa, the current distribution of tsetse and, hence, tsetse-transmitted trypanosomosis is not much different from the ecological limits of the fly distribution. Early work on trypanosomosis, much of it conducted in southern Africa, concentrated on describing the trypanosomes and studying the natural history of the parasites, their vectors and their hosts. The greatest 3|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses advances in knowledge of trypanosomosis over the past two decades have been made in the areas of molecular biology and immunology. EPIDEMIOLOGY Trypanosomes are protozoan parasites of the genus Trypanosoma, order Kinetoplastida, and have, as characteristic organelles, a kinetoplast and a flagellum. Typically, trypanosomes are digenetic parasites and thus require two hosts to complete their life cycle: they multiply in the blood, tissues or body fluids of a vertebrate host and, with the exception of T.equiperdum which is venereally transmitted, are ingested by a haematophagous invertebrate vector. With a few notable exceptions, a cycle of development and maturation occurs in the vector, after which the parasites are transmitted to another vertebrate host as the vector feeds. Transmission is either by inoculation of trypanosomes with saliva or by contamination of mucosa or broken skin with trypanosomes in the vector's faecal material, voided during the blood meal. The type of development cycle within the vector determines whether or not infective, metacyclic parasites are present in saliva or faeces. On this basis mammalian trypanosomes are classified into the two broad sections of ‘salivaria’ and ‘stercoraria’. In Africa, the pathogenic trypanosomes that cause sleeping sickness in humans and nagana in domestic animals are salivarian, and cyclical development occurs in tsetse flies. Transmission of any trypanosome species can take place mechanically without cyclical changes occurring in the vector. In nature, this is effected by biting flies, such as Tabanus, Stomoxys and Lyperosia spp., which feed on more than one animal before repletion. Surra is a disease that affects a wide range of host animals, and it occurs in North Africa, the Near and Far East, Central and South America, Philippines and Mauritius. It is caused by Trypanosoma evansi, a dyskinetoplastic form of which — known as Trypanosoma equinum — also causes disease in equids in Central and South America where it is known as ‘mal de Caderas’ or ‘Murrina’. These parasites have adapted to an entirely mechanical, non-cyclical mode of transmission by blood-sucking flies other than tsetse. Trypanosoma theileri is a stercorarian parasite of cattle which deserves greater mention. It was first reported by Theiler in South Africa in 1903, and has since been found to occur in cattle throughout the world. It is transmitted by tabanid flies and is widely regarded as being non-pathogenic, but in certain circumstances it has been associated with disease. Human sleeping sickness is caused by T. brucei gambiense and T. b. rhodesiense. Whilst these two subspecies do infect some domestic and wild animals, there are other, more significant pathogens of livestock. The remarkable alternate adaptations of these extracellular parasites to mammalian and insect hosts are reflected in morphological changes which are readily detectable by light microscopy. Bloodstream forms 4|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses are trypomastigotes; from the posterior portion of an elongated body, some 8 – 35 µm long, arises a flagellum which extends anteriorly, and which is connected to the body by an undulating membrane. Beyond the anterior extremity of some species, the flagellum may extend free of attachment to the undulating membrane. The beating of the flagellum pulls the trypanosome forwards, imparting characteristic motility. Within the cell, in a posterior position and at the base of the flagellum, a kinetoplast is found, and a single nucleus is located almost halfway along the body. In the tsetse fly, trypomastigotes transform to epimastigotes in which the kinetoplast has migrated anteriorly, to a position adjacent to the nucleus. Differences in the morphology of the trypomastigote stages of the various species form the basis for differential diagnosis. The major characteristics are clearly seen in thin blood smears, stained with Giemsa's, Leishman's or other Romanovsky stains. a b c d Trypanosomes in thin blood smears, x1 000 stained with Diff-Quick. a = Trypanosoma congolense: note absence of free flagellum; b = Trypanosoma vivax : note long free flagellum and large kinetoplast; c = Trypanosoma brucei : note polymorphism, prominent undulating membrane and free flagellum; d = Trypanosoma brucei dividing by longitudinal binary fission. (Unpublished photomicrographs by courtesy of Dr L. Logan-Henfrey, International Laboratory for Research on Animal Diseases, PO Box 30709, Nairobi, Kenya) 5|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Electron microphotograph of Trypanosoma congolense: cross-section showing flagellum (F), nucleus (N), mitochondrion (M) and variable surface glycoprotein coat (VSG), x86 000. Bar represents 0,2 μm. (Unpublished electron micrograph by courtesy of Dr P. Webster, Yale University School of Medicine, Department of Cell Biology, New Haven, CT) 6|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Electron micrograph of Trypanosoma brucei : section through the flagellar pocket (FP) region of the cell. Microtubules are longitudinally sectioned, x44 000. Bar represents 0,4 μm. (Unpublished electron micrograph by courtesy of Dr P. Webster, Yale University School of Medicine, Department of Cell Biology, New Haven, CT) 7|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Scanning electron micrograph of an intermediate (bloodstream) form of Trypanosoma brucei from the blood of a mouse. Note the prominent undulating membrane, pointed posterior end and long, free flagellum. A ‘streamer’ or filopodium can also be seen. (By courtesy of Dr P. Gardiner and reprinted by kind permission of Vinand Nantulya and Parasitology Today ) Trypanosomes show remarkable adaptation. They survive not only in the turbulent blood stream, where they face vigorous immunological assault, but they also withstand the digestive enzymes of the tsetse fly's alimentary tract. Trypanosomes reproduce by longitudinal binary fission, both in the bloodstream and in the fly, although a sexual process can apparently occur in the tsetse fly. Multiplication in each host culminates in the presence of mature trypanosomes, which stop dividing and are pre-adapted to the conditions that they will encounter in the next cyclical host. As a tsetse fly takes its blood meal from an infected host it ingests trypanosomes. Pre-adapted parasites survive in the fly, but trypanosomes that are not metabolically adapted to the new physiological conditions die. The transformation of bloodstream trypanosomes into procyclic or midgut forms within the fly’s midgut is a crucial first step in the establishment of a trypanosomal infection. The mechanism of maturation of a midgut infection is complex and, once established does not always progress to maturation. Before the infection is mature, procyclic forms 8|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses transform into epimastigote and then to metacyclic forms. From the midgut, trypanosomes migrate to the mouthparts or salivary glands. As the infective tsetse fly feeds, metacyclic trypanosomes and saliva pass through the hypopharynx and are inoculated intradermally; it is here that infection is established. From the skin, the trypanosomes reach the blood via the draining lymphatics within a few days. Trypanosomes multiply in the bloodstream, and although initially their low numbers make detection difficult, the generation time of only a few hours soon leads to high levels of parasitaemia. Trypanosomes may leave the bloodstream to reach various extravascular sites. The ability of trypanosomes to establish prolonged infections is attributable to the phenomenon of antigenic variation. Each bloodstream trypanosome is completely clad in a dense surface glycoprotein coat. Within a population of trypanosomes originating from a single infection, almost all bear the same glycoprotein coat and are thus of the same antigen type. As parasitaemia rises, a swift antibody response is elicited against the antigen type exposed on the surface of the bloodstream trypanosomes. These specific antibodies attach to the surface glycoprotein and produce complement-mediated lysis of all trypanosomes of that antigen type. However, before antibodies reach trypanolytic levels, some trypanosomes — as few as one in 100 000 — switch off the gene that controls the production of the initial surface glycoprotein and activate a gene that codes for a different protein. Trypanosomes which bear the new surface glycoprotein are of a different antigen type and are not destroyed by antibody against the first antigen type; they survive to produce another parasitaemic wave, which in turn is removed by antibody specific for that antigen type. By this time a third variant has arisen, and, escaping the effect of host antibody, it survives to produce the next parasitaemic peak. This antigenic variation is the result of sequential expression of variable surface glycoproteins (VSGs) which constitute a repertoire of variable antigen types (VATs). Infections arising from a single trypanosome may have a repertoire of more than 100 VATs. Thus shielded from total destruction, trypanosome infections usually run prolonged courses, since each VAT is present for several days before being removed. Although within a parasitaemic peak there is a mixture of a small number of VATs, the sequence of expression of VATs tends to be quite stable in clonally-derived trypanosomes. This imparts immunologically distinct characteristics to a strain of trypanosomes, the distinct strain being called a ‘serodeme’. In the course of successive parasitaemic waves, some trypanosomes stop dividing and transform to the pre-adapted form able to survive in the tsetse. After ingestion by the tsetse, pre-adapted trypanosomes shed the glycoprotein coat, transform, multiply and finally mature. Infective tsetse then transmits metacyclic trypanosomes to another host. Irrespective of the VAT of the bloodstream trypanosomes ingested by a fly, the metacyclic VATs of a serodeme are relatively constant. The antigenic diversity within a species leads to the possibility of animals in a tsetseinfested area being exposed to a large number of antigenically distinct trypanosomes, but although trypanosomes within a species may be antigenically dissimilar, they are morphologically indistinguishable. 9|Page Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses The characterization of trypanosomes relied for a long time on comparisons of their morphology, motility, host specificity, tsetse transmissibility and their development within the fly, but more recent characterization methods include isoenzyme typing, analysis of kinetoplast DNA by polyacrilamide gel electrophoresis, pulsed field gradient electrophoresis of chromosomal digests and DNA hybridization. The sequel to infection with salivarian trypanosomes is not always disease. The outcome is determined by many factors, frequently related to the susceptibility of the host and the pathogenicity of the trypanosome. In the case of wild animals, a natural cycle of trypanosome transmission occurs which is not associated with disease. Similarly, in some breeds of domestic animals, infection with salivarian trypanosomes is tolerated, and host and parasite reach an equilibrium. Disturbance of the equilibrium may precipitate disease a long time after establishment of the infection. Thus, although the tsetsetransmitted trypanosomes are aetiological agents of African trypanosomosis, infection is not always synonymous with disease. The occurrence of T. theileri in healthy cattle throughout the world exemplifies a well-developed host-parasite relationship. Similar events occur in many wild animals which harbour tsetse-transmitted trypanosomes. Infected animals show no clinical signs, but when they are subjected to the stress of capture, for example, their immunity is reduced, parasitaemia flares up and clinical disease may be precipitated. The epidemiology of African trypanosomosis is almost entirely dependent on tsetse flies. African trypanosomes are well-adapted parasites of many species of wild animals, and sylvatic cycles of trypanosome transmission occur throughout the 10 million square kilometres infested by this unique vector. The natural hosts of salivarian trypanosomes usually show no clinical signs of infection, host and parasites being in equilibrium. The large numbers of naturally infected wild animal hosts constitute a huge reservoir of trypanosomes. Once infected, tsetses remain so for life and thus they too form a reservoir of infection. Consequently, when domestic animals are introduced into areas in which sylvatic cycles of trypanosome transmission occur, trypanosomosis always emerges as a serious disease. Wild animals are the natural hosts of T. brucei rhodesiense, the aetiological agent of human sleeping sickness in central, eastern and southern Africa. Thus, people living and working in tsetse areas are at risk of contracting the disease, but for animal trypanosomosis to occur it is not always necessary for livestock to enter tsetseinfested areas; tsetse also move. Changes in land use may also alter the extent of tsetse infestation. The abandonment of cultivation, for various reasons, permits the regrowth of vegetation, which may then provide suitable tsetse habitat. Conversely, the intensive settlement and cultivation seen in some areas destroy tsetse habitat. The trypanosomal infection rate in tsetse is of prime importance. The ease with which infections develop in tsetse depends upon the fly’s vectorial capacity and specific factors related to the blood of host animals. Generally speaking, the duration of the development of trypanosomes in tsetse increases with increasing complexity of the developmental cycle. 10 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses The prevalence of trypanosomal infections in tsetse is also affected by host preference. Two aspects are important in this context: first, there is diversity of host preference among Glossina and, second, different species of hosts vary in their susceptibility to infection with the different species of trypanosomes. Of particular importance is the relationship between absolute tsetse density and biting rate. A good understanding of this relationship is essential when predicting the impact of tsetse control interventions. Usually the challenge increases with the tsetse population density but even at low densities tsetse can still cause a substantial disease problem. This is partly attributed to the often observed, increased frequency with which flies that have metacyclic infections in their mouthparts probe. Management practices may also alter the challenge to which livestock are subjected, and in this sense management is central to the epidemiology of trypanosomosis. The different management of calves and adult cattle can reduce significantly the level of challenge to which young animals are subjected. Observations on the grazing ranges of livestock in West Africa showed that while cattle foraged widely in tsetse-infested habitat, sheep, goats and donkeys remained closer to the villages. As a result, small ruminants and equids were less exposed to attack by tsetse than cattle. A major influence on the epidemiology of trypanosomosis is the use of trypanocidal drugs. Whilst permitting the use of tsetse-infested land, chemotherapy may alter the prevalence of trypanosome species in the area. In Zimbabwe, it was reported that the use of diminazene increased the prevalence of T. vivax in cattle. Furthermore, the repeated use of trypanocides may result in the emergence of drug resistant strains of trypanosomes, and the epidemiological picture then changes. Wild animal hosts of tsetse, such as kudu, warthog, bushbuck, bushpig, buffalo, elephant and rhinoceros, are vital to sylvatic cycles of trypanosome transmission and they form a major reservoir of infection. The increasing popularity of game ranching or farming frequently entails the translocation of these animals from tsetse habitats to tsetse-free areas. As a result, clinical disease may occur in animals normally regarded as ‘immune’, in the absence of the tsetse vector. There is also the possibility that trypanosomes could be mechanically transmitted from these species to livestock. Wild animal hosts of tsetse and certain West African taurine cattle are tolerant of tsetse-transmitted trypanosomes. Carefully controlled experiments have shown that the trypanotolerance of N’Dama cattle and of African buffalo (Syncerus caffer) is an innate characteristic. This trait also occurs in some breeds of sheep and goats. Lower trypanosomal parasitaemias and less severe anaemia occur in trypanotolerant livestock than in trypanosusceptible animals. The epidemiological significance of this trait lies in the lower morbidity and mortality due to trypanosomosis in tolerant breeds. Although trypanosome-infected animals may effect self-cure, the usual sequel to infection in tolerant animals is the establishment of a balance between host and parasite. If the host is stressed, the equilibrium is disturbed and a clinical episode of variable severity is precipitated. Stress takes many forms. Animals in late pregnancy or which are lactating are more susceptible to trypanosomosis. Overwork also constitutes a stress, which in infected trypanotolerant stock may precipitate disease. This has serious consequences for animal traction and for hard-working bulls used in restricted breeding seasons in tsetse-infested areas. In a similar 11 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses manner, intercurrent disease is stressful; trypanosome-infected animals with helminthosis or other diseases are more severely affected than those with either disease alone. The combined effect of poor nutrition and increased exercise is often associated with an increased incidence of trypanosomosis in the dry season. Age also has a significant effect on resistance to trypanosomosis. It is widely recognized that cattle born in an infested area do not immediately succumb to disease, even though they acquire trypanosomal infections when young, whereas cattle brought into the area readily succumb. Despite the antigenic complexity of trypanosomes, infected animals do mount immune responses which, especially when supported by chemotherapy, can confer specific protection against homologous serodemes. Thus, within a defined area, animals may acquire protective immunity against locally prevalent serodemes. However, the movement of these animals to another area may expose them to different strains, or serodemes, to which they may succumb. This type of tolerance requires continuous challenge and explains the low mortality of cattle in high challenge, endemic areas (such as the Eastern Province of Zambia) where trypanocidal drugs are used to prevent animals from succumbing to nagana. The introduction of tsetse control measures in such areas would result in a loss of immunity that would render the population highly susceptible to infection and disease. If tsetse control measures breakdown severe outbreaks of trypanosomosis would ensue, probably with high mortality. The African continent is experiencing considerable environmental changes as a result of an unprecedented demographic growth.The increasing human pressure and the demand for arable land is a major driver of land-use change resulting in deforestation, erosion and loss of biodiversity and suitable habitats for tsetse flies Like many other arthropods, tsetse flies rely for their survival on habitats with suitable conditions.The destruction and fragmentation of habitats has had important repercussions for the density and distribution of tsetse flies. Insome instances,the drastic changes have resulted in the disappearance of certain species In many cases, however, tsetse flies persist at lower densities in what might appear to be unsuitable environments probably occupying microclimatic niches. The effects of anthropogenic environmental changes on the epidemiology and impact of livestock trypanosomosis are largely driven by the degree with which the tsetse flies adapt to an environment where domestic animals are becoming increasingly important for their survival.These changes have been demonstrated in southern Africa where epidemiological studies have clearly revealed the effect of such anthropogenic changes on the impact of the disease on livestock and the appropriatenesss of particular control strategies. PATHOGENESIS The precise pathogenesis of the trypanosomoses remains far from clear. Three features — anaemia, tissue damage and suppression of immune responses — dominate the pathology of trypanosomosis. 12 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Infection becomes established at the site of inoculation of metacyclic trypanosomes in the skin, where a chancre may form. Multiplication of the parasites induces an inflammatory response, the severity of which depends upon the size of the inoculum, the species of parasite and the breed and species of the host. These may originate from either the host or the parasites. The chancre reaches a maximum diameter, of some 100 mm, ten to 14 days after an infective tsetse fly feed, its development preceding invasion of the bloodstream by trypanosomes, and is accompanied by enlargement of the draining lymph nodes. Trypanosomes are detectable in the blood 13 to 16 days after an infective tsetse fly has fed. At this time the chancre begins to regress, and the characteristic series of intermittent parasitaemias begins. Bone marrow impression smear from a calf with an acute Trypanosoma vivax infection, showing a macrophage which has phagocytosed several erythrocytes x1 200. Stained with Wrights-Leishman. (Unpublished photomicrograph by courtesy of Dr L. Logan-Henfrey, International Laboratory for Research on Animal Diseases, PO Box 30709, Nairobi, Kenya) After an infection has become established, a protracted battle ensues as the parasite provokes an immune response. Anti-VSG antibodies destroy large numbers of trypanosomes. The destruction of large numbers of parasites releases lysosomal and other enzymes as well as structural proteins. Some of these enzymes have been identified and are thought to be directly harmful to the host. Many biological 13 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses mediators such as the vasoactive amines are released from activated or damaged host cells. Anaemia is a cardinal sign of trypanosomosis in many domestic animals, and the aetiology is probably similar in all species. There is no single cause of the anaemia in trypanosomosis; the pathogenesis is complex and involves a variety of mechanisms. Accompanying anaemia is leukopenia, which possibly arises from direct inhibition of stem cell differentiation. There are several possible sequelae to the early phase of infection, which depend largely on the nature of the parasite and the susceptibility of the animal. There may be spontaneous recovery or death, but very often there is a chronic phase which is characterized by infrequent, low-grade parasitaemias. Animals lose weight and condition and, as a result of dyshaemopoiesis, remain anaemic. Extensive haemosiderosis occurs as a result of erythrophagocytosis, and the trapping of iron in phagocytes is believed to contribute to the failure of erythropoiesis. Despite the apparent absence of parasites in the circulation, red blood cell destruction continues, and insufficient erythropoietic compensation results in persistent anaemia. The pathogenesis of tissue lesions varies with the species of trypanosome. Trypanosoma congolense and T. vivax are mainly intravascular parasites; they induce changes in the endothelium of capillaries, and so indirectly cause damage to adjacent tissues. Trypanosoma brucei, on the other hand, has an affinity for tissues. Its presence in the extravascular compartment is associated with marked lesions in parasitized tissues. The severity of endothelial injury also depends on the interaction of host and parasite. Trypanosoma congolense often attaches to erythrocytes and capillary endothelium. Damage to endothelial cells by parasite products, immune complexes, vasoactive amines and cytokines increases vascular permeability. In T. congolense infections a generalized dilatation of capillary beds, which alters the haemodynamics, is observed. The concomitant anaemia and more sluggish tissue perfusion affect the exchange of metabolites and are associated with intracellular oedema of capillary endothelial cells. Fibrinous microthrombi form in response to endothelial damage. These changes can be prominent in T. vivax infections, with which disseminated intravascular coagulation is more commonly associated. Alterations to the microcirculation produce secondary degenerative changes in tissues. As capillary permeability increases, phagocytes and products of the parasite extravasate more readily and are responsible, in part, for some of the tissue lesions. Cytokines and parasite-derived substances, such as proteases, may directly injure host cells, an effect that is exacerbated by the formation of immune complexes in tissues. In the case of T. brucei, parasites localized in tissues cause mechanical disruption of host cells and probably also have some direct toxic effect on host cells, possibly mediated by their enzymes. The cellular response provoked by parasites within the tissues causes further damage, and auto-antibodies are also thought to play a role in inducing lesions. The influence of all of these changes on the course of the disease depends upon their severity and upon the degree of impairment of the affected organs. 14 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses An important feature of the pathogenesis of trypanosomosis is the effect on lymphoid tissue. As the disease progresses, the volume of tissue in the spleen, lymph nodes and bone marrow inreases markedly. This hyperplasia of reticuloendothelial cells reduces lymphoid cell density, and eventual lymphoid depletion can occur. Chronic disease is associated with progressive emaciation and eventually cachexia. This is usually accompanied by low levels of parasitaemia but the pathogenesis is poorly understood. The reduced lifespan of red blood cells, the increased catabolism of many proteins, and the associated negative nitrogen balance of clinically affected animals would appear to be largely responsible for the pitifully thin condition of animals suffering from chronic trypanosomosis. DIAGNOSIS AND DIFFERENTIAL DIAGNOSIS Diagnosis and differential diagnosis Clinical signs and pathology Cattle Recognition of the wide range of clinical signs associated with the disease is important, both within and outside endemic areas. The course of disease due to infection with salivarian trypanosomes is variable and there are no clinical signs specific to bovine trypanosomosis. Trypanosomosis in cattle may be acute, subacute or chronic; acute disease may be fatal after brief illness lasting two to six weeks, but chronic disease lasting many months or even years is more common. Chancres are rarely seen in cattle with naturally-acquired infections. The first signs of disease are due to the fever which accompanies the onset of parasitaemia. As parasitaemia falls, so does fever; the course of infection is therefore characterized by fluctuating parasitaemia and parallel fluctuations in body temperature. Intermittent parasitaemia and malaise are followed by increasingly severe clinical signs. Acute trypanosomosis causes sudden reduction of milk yields and also abortion. Oxen, bulls and young stock with acute infections are reported to be suddenly ‘off-colour’. They are generally in good bodily condition but are dejected, their ears droop and they may walk stiffly. Pulse and respiratory rates are raised in febrile animals and there may be piloerection. Conjunctival mucosae may be congested early in the acute phase but later, as anaemia develops, pallor of the mucous membranes is evident. 15 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Acutely affected animals quickly lose weight and body condition, although they continue to eat. They become weak, dejected and lethargic, often standing alone, away from the rest of the herd, not seeking shade. Excessive lachrymation is sometimes seen at this stage. After a short illness some acutely affected cattle become recumbent for a few days before death occurs. A proportion of acutely affected cattle gradually improve to enter the chronic phase of the disease. Subacute trypanosomosis in an ox: the animal has lost weight and condition and it is dejected, with drooping ears and flaccid tail. (Reprinted by kind permission of FGU Consulting and Engineering GmbH, Königstein, Germany, and the Regional Coordinator, RTTCP, Harare, Zimbabwe) In contrast to the acute disease, subacute trypanosomosis is more common and runs a more prolonged course, with many animals apparently making a spontaneous recovery. In subacute cases, animals are intermittently ‘off-colour’, becoming weak and dejected, with drooping ears and flaccid tail. Within a few weeks they lose weight and condition, the coat becomes dull and a marked jugular pulse develops. Superficial lymph nodes and haemal lymph nodes are frequently enlarged and are readily visible. Gradually, the frequency and intensity of parasitaemia decrease and fever subsides. Dependent oedema, particularly in the submandibular region, may develop. Thin, rough-coated, anaemic, lethargic cattle with generalized lymph node enlargement are said to have a ‘fly-struck’ appearance. Death may occur between 4 and 6 months after the onset of disease, but many animals make a gradual recovery, which is assisted by a good plane of nutrition. 16 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Chronic bovine trypanosomosis is by far the most common form of the disease in endemic areas. Severely affected cattle may be extremely emaciated, having lost a large proportion of their muscle mass; they are often just ‘skin and bone’. Their general condition is very poor — the hair is sparse and the coat is rough, dull and staring. Mucous membranes are very pale, the pulse is rapid, and breathing is laboured; in resisting restraint, the animal may collapse in respiratory distress after brief exertion. The combined effect of anaemia, circulatory disturbance and myocardial damage frequently produces acute cardiac decompensation which leads to sudden death from congestive heart failure. The gross pathology of affected animals varies with the duration and severity of the disease, and many organs and tissues are affected. The overall appearance of the carcass is one of paleness and petechial and ecchymotic haemorrhages are frequently present, especially on serosal surfaces. Lymph nodes are enlarged and, when incised, are oedematous and often have a dark pigmented medullary area. Typically, the spleen is greatly enlarged and dark red. Excessive peritoneal fluid is present which is often blood-tinged. The gross pathology of chronic trypanosomosis is characterized by cachexia and anaemia. The coat is dry and dull, the skin is scaly and inelastic, the eyes sunken and skeletal muscles are atrophied. There is hydrothorax, hydropericardium and ascites, and the carcass, in general, appears oedematous. Residual fat around the heart and kidneys is gelatinous. Lymph nodes, haemal nodes and spleen are generally enlarged and have an appearance similar to that seen in acute trypanosomosis, but after prolonged disease there may be splenic atrophy. The heart is enlarged and flabby, the liver swollen and pale. Whereas red marrow is present in the long bones in acute trypanosomosis, all the bone marrow is yellow and gelatinous in the chronic disease. Gross changes are not always obvious, but microscopic lesions are extensive. These are found particularly in the cardiovascular system and lymphoid tissues, and there is widespread infiltration of many organs by mononuclear inflammatory cells. Dilation of the microvasculature, and oedema and structural changes in vessel walls are usually seen, and trypanosomes are commonly present in the lumens of blood vessels. A constant feature of trypanosomosis is initial hyperplasia of the lymphoid tissues and of the mononuclear phagocytic system. The heart is commonly affected, and macrophages, lymphocytes and plasma cells infiltrate the myocardium. Degenerative changes and focal necrosis of myocytes and, in long-standing infections, fibrosis are evident in the heart. Infiltrations also occur frequently in other organs such as the pituitary, thyroids, adrenals, kidneys and gonads. Congestion and interstitial oedema also occur, and, when the substantia propria is affected, corneal opacity then develops. Sheep and goats The general clinical signs of trypanosomosis of goats and sheep are similar to those in cattle. Acute disease is characterized by intermittent and increasing dullness. During bouts of fever, sick 17 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses animals walk stiffly, or stand with head held low, ears drooping and tail flaccid. Early morning rectal temperatures may exceed 41 C in the febrile periods, when the pulse is rapid and the breathing shallow and fast. Anaemia develops rapidly and the PCV falls from 0,35 l/ – 0,18 l/ or less in two to four weeks, when mucous membranes become pale. Affected animals rapidly lose weight and condition, and may die within a month of the onset of detectable parasitaemia. The signs of subacute trypanosomosis are less marked, and the course of the infection resembles that in cattle. Chronic infections are very common and may develop without marked clinical signs. Parasites are rarely found in the blood and animals are afebrile. Clinically, it is difficult to distinguish such animals from those with helminthosis. Furthermore, the immunosuppression induced by trypanosomosis in goats and in sheep permits the establishment of large helminth burdens, which thus exacerbate the clinical signs in affected animals. The pathology of trypanosomosis in goats and sheep does not differ greatly from that in cattle. A goat with a naturally acquired Trypanosoma vivax infection: prescapular lymph nodes are greatly enlarged Pigs Trypanosoma brucei and T. congolense infections in pigs generally produce only mild disease. Occasionally, anaemia occurs, which is accompanied by loss of condition and progressive weakness leading to incoordination. Trypanosoma suis infections are of minor clinical importance and have only rarely been recorded; infections run a chronic course, but may kill young pigs in less than two months. 18 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Trypanosoma simiae infections in pigs eclipse in importance all other trypanosomal infections in this host. This parasite may kill pigs after an incubation period of only four to six days, and infected pigs may collapse and die within 24 hours of the first signs of disease. Affected pigs are dull and inappetant or completely anorexic. They have a stiff, unsteady gait and eventually become prostrate. Dyspnoea is evident and cyanosis may be seen. Some pigs froth at the mouth and there may be diarrhoea. Rectal temperatures reach 41 C but the extremities are usually cold. The necropsy findings vary considerably, and there are no pathognomonic changes. The blood is usually cyanotic, clots slowly, and chicken fat clots may be found in the heart. Haemorrhages affect the epi- and endocardium, kidneys and serosal surfaces. The spleen is usually enlarged and the pulp soft with a ‘strawberry jam’ appearance; there is excessive serous or serosanguinous fluid in the pericardial sac and pleural and peritoneal cavities; the trachea may be filled with froth when there is also pulmonary oedema. Thoracic and abdominal lymph nodes are usually enlarged and oedematous, and may be haemorrhagic. Horses and donkeys The general clinical signs resemble those seen in ruminants. Horses with acute trypanosomosis are very dejected. Intermittent parasitaemia occurs and is accompanied by fever; rectal temperatures may reach 40,5 C and there is tachycardia. Oedema of the lower limbs occurs early in the course of disease, as affected horses are less active than normal. Subcutaneous oedema then affects the ventral thorax and abdomen. Oedematous plaques may form on the flanks, but dependent oedema is more pronounced. Weakness sets in rapidly. Initially, mucous membranes may be congested and icteric but they become pale as the disease progresses. The condition of the animal deteriorates rapidly and weakness may progress to paraplegia. Oedema becomes increasingly severe. Animals die within two to four weeks from the onset of clinical signs. Chronic trypanosomosis commonly occurs in horses and donkeys. Animals lose condition and weight and the coat becomes harsh and dry. Animals become extremely weak and show signs of ataxia, but usually continue to eat. Subcutaneous oedema, initially affecting the limbs and ventral abdomen, may extend to the sheath, scrotum, perineum and occasionally the head. The general pathology of equine trypanosomosis resembles that occurring in cattle. 19 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses A horse with a chronic Trypanosoma congolense infection showing emaciation and ventral oedema Dogs The general clinical signs of trypanosomosis in dogs resemble those in livestock, but the course and severity of the disease are modified by the virulence of the parasite and the susceptibility of the host. In acute disease the dog suddenly becomes dejected and inappetant. Pulse rate, respiratory rate and rectal temperature are raised. Anaemia develops rapidly; mucous membranes become pale and the dog shows signs of weakness and depression. There is progressive weight loss; as the condition deteriorates, the coat becomes dry and subcutaneous oedema, particularly of the head and limbs, may develop. Ocular lesions characterize both T. brucei and T. congolense infections but are more severe in the former case. Trypanosoma brucei invades all tissues of the eye, producing blepharitis, conjunctivitis, keratitis and uveitis. Additionally, T. brucei invades many other tissues, and by entering the cerebrospinal fluid causes ataxia and partial paralysis. Death ensues within four to 10 weeks of the onset of signs. Infections with T. congolense generally follow a more protracted course. In chronic cases, dogs become severely emaciated, and ulceration of the oral and gastrointestinal mucosa may develop, leading to haemorrhage and melaena. Inappetance may only be intermittent, with animals being periodically ‘off colour’. 20 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses Lesions of trypanosomosis in dogs are broadly similar to those found in livestock. Anaemia is the most readily detectable change early in the course of infection, and the PCV drops markedly. Whilst T. congolense infections produce extensive interstitial oedema and ulceration of gastrointestinal mucosa, tissue lesions induced by T. brucei are much more severe. There is marked cellular infiltration and cellular degeneration and necrosis. The heart, choroid plexus and eyes are consistently and severely affected. Laboratory confirmation The specific diagnosis of trypanosomosis is notoriously difficult. Not only are there no specific clinical signs, but the intermittent and frequently low parasitaemias make detection of the parasites difficult. Furthermore, infection is not synonymous with disease; many subclinically infected animals live in delicate balance with potentially pathogenic trypanosomes. An element of clinical judgment is, therefore, necessary when making a diagnosis of trypanosomosis. The detection of infection in a few clinically affected cattle warrants careful examination of the entire herd. The only way to confirm diagnosis in clinically infected animals is to demonstrate and identify the parasites in body fluids. The parasite detection methods are highly specific but their diagnostic sensitivity is relatively low. This is especially the case when results are considered for an individual animal rather than in a herd. As a result, the apparent prevalence of trypanosomosis determined by parasitological diagnostic tests is an underestimate of the true parasitological prevalence. This is a problem in areas where the disease is present at low prevalence or is seasonal, or when attempting to confirm the absence of the disease in a particular area. The body fluid most commonly examined is blood, either capillary blood from the tip of the tail or venous blood from an ear vein or from the jugular vein. Lymph, aspirated from a punctured superficial lymph node (usually the prescapular), provides useful supplementary diagnostic material. Wet blood smears are prepared by placing a drop of blood (about 2 µl), taken directly from a punctured ear vein or the tip of the tail, onto a clean, dry, grease-free slide. This is immediately covered with a cover slip and examined microscopically as a fresh, wet preparation with a x25 or x40 objective lens. Approximately 50 to 100 fields are examined. Live, motile trypanosomes may be seen as they bore their way between blood cells (the name “trypanosome” is derived from the Greek “trypanon” a borer and “soma”, meaning body). More commonly, for routine diagnosis in veterinary practice, thick and thin smears of blood are prepared. A drop of blood taken directly from a punctured blood vessel in the ear or tail tip is placed on a glass slide; a thick and a thin blood smear can then be made on a single slide. Lymph smears are prepared in a similar manner. The unfixed de-haemoglobinized thick smear allows approximately 120 times more blood to be scanned than a thin smear and, thus, has 21 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses higher diagnostic sensitivity than the thin smears. Trypanosomes are easily recognized by their general morphology but may be damaged during the staining process. This makes it difficult to make a species-specific identification on thick smears. The thin smear, on the other hand, permits accurate speciation of the parasites. However, due to the staining process, results are delayed. The development of anti-trypanosomal antibody detection techniques has been a major improvement in the serodiagnosis of trypanosomosis. The indirect immunofluorescent antibody test (IFAT) has been and still is used widely to diagnose trypanosomosis. The test has undergone several modifications so that it can differentiate, to a limited extent, between trypanosome species in ruminants. The serodiagnosis of trypanosomosis has greatly benefited from the introduction of enzyme immunoassays. The ELISA compares favourably with the IFAT and has been found to give results that correlate with the local history of trypanocide usage. However, even if a trypanosomal infection has been cured, anti-trypanosomal antibodies persist for several months and antibody detection tests do not distinguish between current and past infections. They can only provide a presumptive diagnosis. A polymerase chain reaction (PCR) method has been developed for the diagnosis of infections with African trypanosomes in humans, animals and tsetse flies. Specific repetitive nuclear DNA sequences can be amplified for T. vivax and each of the three T. congolense subgroups. A common primer set is available for detection of the three T. brucei subspecies. Despite the development of more sensitive, more sophisticated and expensive diagnostic methods the clinician will, for the foreseeable future, have to rely upon examination of blood smears, buffy coat preparations and findings at necropsy to confirm diagnoses of trypanosomosis. The diagnosis of trypanosomosis in small ruminants is no different from that already discussed for cattle, but it is much more difficult because of the larger proportion of subclinical infections. In these cases very low parasitaemias occur in the absence of obvious clinical signs. It is undoubtedly these factors which are responsible for the serious underestimation of trypanosomosis in small ruminants. Differential diagnosis In its various stages bovine trypanosomosis resembles a number of other disease conditions, and it frequently occurs at the same time as other infections. In the acute febrile stage, bovine trypanosomosis must be differentiated from redwater (babesiosis), anaplasmosis and East Coast fever. The haemorrhagic syndrome in acute T. vivax infection can be distinguished from anthrax and haemorrhagic septicaemia (Pasteurella multocida infection) by examination of Giemsa-stained blood smears, when large numbers of trypanosomes are found. The main disease entity which resembles trypanosomosis in goats and sheep is helminthosis, especially haemonchosis. Anaemia, illthrift, weight loss, submandibular oedema and high mortality rates are common to the two disease complexes, although diarrhoea sometimes accompanies helminthosis. 22 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses A history of sudden death in a number of pigs, combined with clinical signs as described, is suggestive of T. simiae in areas adjacent to tsetse infestations. Other causes of sudden death that should be differentiated include African and European swine fevers and anthrax. Chronic trypanosomosis in pigs should be differentiated from helminthosis and malnutrition. Acute disease in horses and donkeys must be distinguished from African horsesickness, anthrax and babesiosis. In chronic trypanosomosis the oedema has to be differentiated from that occurring in African horse-sickness and the anaemia from that of equine infectious anaemia. Chronic babesiosis may also produce signs similar to chronic trypanosomosis. The signs of weight loss, emaciation and oedema caused by dourine (T. equiperdum infection), strongylosis, malnutrition and dental disorders should be differentiated from those caused by salivarian trypanosomes. The anaemia in dogs caused by trypanosomosis must be differentiated from that arising from infection with Ancylostoma caninum, Babesia canis or Ehrlichia canis. Hookworm infection may be concurrent with trypanosomosis. Bilateral corneal opacity produced by trypanosomosis must be distinguished from that seen in canine hepatitis (adenovirus-2 infection). CONTROL / PREVENTION For more than 60 years, Governments in southern Africa have made concerted efforts to control tsetse in their battle against trypanosomosis. The ultimate goal, and the basis of many control strategies, was the eradication of the tsetse fly. Despite considerable gains in some countries, few achievements have been sustained because of the prohibitive cost of preventing tsetse from re-invading previously cleared areas in the absence of natural barriers. Nagana, therefore, remains a serious constraint to development throughout much of Africa and new approaches towards its control have been devised. The new control strategy aims at intervening in areas where achievements are likely to be sustained by the beneficiaries. Consequently, in most cases, trypanosomosis will be controlled on a smaller, localized scale in areas that have to be selected carefully. Several methods are available to control trypanosomosis. They may be directed against the vector or the parasite, or towards the livestock and modified management. In choosing a method, technical and economic aspects have to be considered and the most viable option selected. Some of the tsetse control techniques, which have been developed with large-scale eradication in mind, are not well suited for use in localized, small-scale control operations. Currently, tsetse control utilizes odour-baited, insecticide-treated cloth targets and insecticide-treatments of cattle. Odour-baited, 23 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses insecticide-treated targets have been very effective in Zimbabwe and have been employed in other African countries to control various species of tsetse under different ecological conditions. Despite the proven effectiveness of insecticide-treated cattle and odour-baited targets, the success of a tsetse control operation will depend largely on its planning and implementation. Trypanocidal drugs will continue to play an important role in the integrated control of trypanosomosis. They appeal to communal cattle owners because they provide a means of protecting private goods and may achieve impressive results at low costs. Although the low-level usage of these drugs may reduce mortality rates, such usage generally has low impact on animal production. This will be the case particularly in high challenge areas and when farmers use trypanocides therapeutically rather than prophylactically. The development of resistance in trypanosomes to trypanocides is a continuous threat to their sustainable use in the control of nagana. Even in areas where resistance to trypanocides has not yet been demonstrated, the probability of its development should influence the selection of an appropriate control strategy. Millions of domestic animals are kept in tsetse-infested areas, but the degree to which they are exposed to tsetse is frequently determined by management practices. The avoidance of heavily infested watering points or grazing reserves reduces challenge, and thus controls trypanosomosis to some extent, but the availability of alternative water sources and supplies of feed are then necessary. Prevention of livestock movement into tsetse habitats also limits the disease, but the enforcement of livestock movement control in Africa is not easy, because of communal grazing practices, seasonal migrations and local political pressures usurping the law. Under light tsetse challenge a therapeutic approach to control can be successful and economical. There are numerous well-documented examples of susceptible breeds of livestock being successfully reared in many tsetse-infested areas of Africa as a result of chemotherapy or chemoprophylaxis. To control trypanosomosis successfully by chemoprophylaxis, it is necessary to treat a high proportion of cattle at risk at regular intervals. This is not always possible, especially if owners have to pay for treatment, and consequently, a therapeutic approach to control is widely used. There is considerable evidence that such therapeutic control assists the acquisition of protective immunity to locally prevalent trypanosome serodemes, whereas with chemoprophylaxis immunity to trypanosomes does not seem to develop. The satisfactory treatment of trypanosomosis requires more than a correctly administered trypanocidal drug, and the speed of recovery is largely determined by the plane of nutrition, the amount of exercise during convalescence and the duration of the disease. Well-rested and well-fed animals recover more rapidly after trypanocidal therapy than do undernourished animals which have to trek long distances to pasture and water. However, chronic trypanosomosis often fails to respond to therapy. Diminazene and homidium are therapeutic trypanocides, although homidium does provide prophylaxis for several weeks. Isometamidium, whilst having a curative effect, is widely regarded as being the only trypanocide worthy of being used for prophylactic purposes. Trypanocides are toxic compounds and have 24 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses narrow therapeutic indices, so therefore the dosage rates cannot be greatly increased to obtain greater trypanocidal efficacy. Furthermore, they are not equally effective in the treatment of all species of trypanosomes. A single injection of diminazene cures T. congolense and T. vivax infections at a dosage rate of 3, 5 mg/kg but a dosage rate of 7, 0 mg/kg is necessary to treat T. brucei infections. The efficacy of the phenanthridine compounds, homidium and isometamidium against T. congolense and T. vivax is more marked than against T. brucei at recommended dosage rates. Dosage rates of isometamidium in excess of 2, 0 mg/kg are toxic to cattle, and even below this level, the drug is irritant and has marked local effects. The prophylactic effect of isometamidium depends upon the slow release of the drug from the depot created at the intramuscular injection site. Isometamidium is irritant and at the site of injection, it induces an intense inflammatory response and causes necrosis of muscle fibres. This lesion becomes encapsulated and is later organized by fibrous connective tissue. From this depot, the drug is slowly released. Attempts to overcome this irritant side effect by reformulation of the drug have been unsuccessful to date. The repeated inoculation of cattle kept under tsetse challenge induces extensive muscular scarring, and to reduce carcass damage and subsequent losses from condemnation, the drug is usually administered into the less economically valuable neck muscles, often with untoward side effects. The use of trypanocides at subtherapeutic levels is believed to promote drug resistance in trypanosomes. Since diminazene is rapidly excreted, the risk of trypanosomes being exposed to sublethal levels of the drug is less than is the case with isometamidium. Prophylactic levels of isometamidium wane over much longer periods. Even when cattle receive isometamidium at the correct dosage rate, it may be administered at the wrong time. Tsetse challenge may increase at a time when drug levels have declined and are not protective. True resistance to trypanocides is a spectre which looms large. The development of resistance has led to the withdrawal of many trypanocides in the past, and resistance to the few remaining currently-used trypanocides is well documented. The degree of resistance in trypanosomes is not clear cut; a spectrum of drug sensitivity occurs, and resistance to low levels of a drug may be overcome by increasing the dosage rate. The antigenic complexity of trypanosomes has thwarted attempts to develop a vaccine. Although potential immunological targets within the parasite have been identified, no vaccine will be commercially available in the near future, and the greatest hope for the immunological control of animal trypanosomosis lies in the exploitation of trypanotolerant breeds of livestock. It is intended, through a process of selective breeding of trypanotolerant animals, to reduce the problem of trypanosomosis and to improve livestock productivity. However, this approach offers little hope for southern Africa today: inheritance is complex and the number of trypanotolerant animals is relatively small. On the other hand, the fact that some Zebu cattle survive in tsetse-infested areas provides a basis for attempting to select the more tolerant of these animals for breeding purposes. 25 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses FAQS 1. Why is trypanosomiasis called “sleeping sickness”? The infection by trypanosomes when reaching the brain causes nocturnal sleeping disorders. Patients are then so tired that they are sleeping during the day. 2. In the literature the disease is called trypanosomiasis or trypanosomosis. What is the correct term? In the past, trypanosomiasis was the sole term that was used. The OIE (Office International des Epizooties) proposed to use trypanosomosis which was accepted by the World Assembly of Delegates of the OIE. Some authors now are using trypanosomiasis for the human forms of the disease and trypanosomosis for the diseases affecting animals. 3. Why are tsetse flies called tsetse? Tsetse (tsé-tsé in French) means in a local language of West Africa “fly that kills cattle”. Saying thus tsetse fly is incorrect as it is repeating the term “fly”. 4. Are tsetse always involved in the transmission of trypanosomes? No, T. equiperdum is transmitted sexually. T. vivax can also be transmitted mechanically by blood sucking flies and evidences are accumulating that T. congolense might also be transmitted mechanically. 5. It seems that sleeping sickness might be different from country to country. Is that indeed the case? Human sleeping sickness can be caused by two different trypanosomes: T. brucei gambiense or T. brucei rhodesiense in West and East Africa respectively. In West Africa, the disease is longer and more chronic than in East Africa where the disease is far more acute. 6. The disease is known for more than a century now. Why is there no vaccination available? Trypanosomes developed a very clever strategy to escape the immune system of the host. The proteins that are covering the trypanosomes and that are recognized by the immune system of the host change regularly in composition. The host has to adapt his response at every change. During this adaptation period, trypanosomes can multiply. This phenomenon is called “antigenic variation”. 26 | P a g e Livestock Health, Management and Production › High Impact Diseases › Vector-borne Diseases › Trypanosomoses REFERENCES 1. Bourn, D., Reid, R., Rogers, B., Snow, B., Wint, W., 2001. Environmental Changes and the Autonomous Control of Tsetse and Trypanosomosis in Sub Saharan Africa Case Histories From Ethiopia the Gambia Kenya and Zimbabwe. Information Press, Oxford UK. 2. Courtin, F., Rayaisse, J.B., Tamboura, I., Serdebeogo, O., Koudougou, Z., Solano, P., Sidibe, I., 2010. Updating the Northern tsetse limit in Burkina Faso (1949-2009): Impact of global change. International Journal of Environmental Research and Public Health 7, 1708-1719. 3. Ducheyne, E., Mweempwa, C., De Pus, C., Vernieuwe, H., De Deken, R., Hendrickx, G., Van den Bossche, P., 2009. Assessing the impact of habitat fragmentation on tsetse abundance on the plateau of eastern Zambia. Prev. Vet. Med. 91, 11-18. 4. Van den Bossche, P., 2001. Some general aspects of the distribution and epidemiology of bovine trypanosomosis in southern Africa. Int. J. Parasitol. 31, 592-598. 5. Van den Bossche, P., de la Rocque, S., Hendrickx, G., Bouyer, J., 2010. A changing environment and the epidemiology of tsetse-transmitted livestock trypanosomiasis. Trends Parasitol 26, 236243. 27 | P a g e