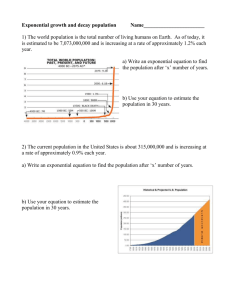
Exponential growth, etc.
advertisement

In this lecture we will look in greater depth at both exponential and logistic growth. This material parallels chapter 1 of the text. We all know the basic equation for exponential growth, but it is important to recognize that there are different forms for the equation to fit different life history/reproductive patterns. The simplest pattern is for populations that have nonoverlapping generations, i.e. parents grow, reproduce, then die. We call such species semelparous. The equation that tells us numbers in such a population is the integrated form of the usual differential equation for exponential growth: dN/dt = N0ert Nt = N0t or Nt = N0Rt In the differential equation: N0 is the starting population size r is the instantaneous rate of increase t is time measured in some continuous unit However, in the integrated equation, reflective of our semelparous population: is the “finite rate of increase” by definition this is the number of individuals ‘replacing’ each adult in the previous generation, and sometimes written as R, the ratio of numbers in two sequential generations. t is the number of generations This is the way to getting you to think of reproduction that does not occur continuously. Most species produce offspring in discontinuous bursts (e.g. once a year or once in a lifetime). Very few living things reproduce in the essentially continuous way assumed as characteristic of the exponential model. Single celled organisms (and possibly other very simple ones) basically fit that assumption. Multicellular ones rarely do. At latitudes where seasonality is characteristic, i.e. at temperate latitudes or more extreme areas, almost all plants reproduce once per year. Responding to resultant food availability, most animals reproduce during the plant growing season. Some reproduce throughout that period, some have two or a few reproductive cycles related to that period, others reproduce once each year. Thus, most species reproduce in discrete bursts. The basic exponential model described continuous growth; that pattern is one of the model's assumptions. An example from your text shows how to apply discrete growth equations: Gypsy moths are insects that reproduce annually, following which the adults die, i.e. they are semelparous. They feed on various tree species, but one of their favourites and the subject of the example is oaks (Quercus spp.). A technician counted the number of egg masses per hectare (each mass averages containing 40 eggs). In 2003 there were 4 egg masses/hectare, or 160 eggs. In 2004 there were 5 such masses per hectare on average, or 200 eggs. What was the finite rate of increase (R or ) for this population? An example from your text shows how to apply discrete growth equations: Gypsy moths are insects that reproduce annually, following which the adults die, i.e. they are semelparous. They feed on various tree species, but one of their favourites and the subject of the example is oaks (Quercus spp.). A technician counted the number of egg masses per hectare (each mass averages containing 40 eggs). In 2003 there were 4 egg masses/hectare, or 160 eggs. In 2004 there were 5 such masses per hectare on average, or 200 eggs. What was the finite rate of increase (R or ) for this population? R = 200/160 = 1.25 What would the population size (number of eggs/hectare) be in 2005? N2005 = N2004R = 250 And in 2006? N2006 = N2004R2 = 200 x 1.5625 = 312.5 Apparently, spraying to control this pest insect is permitted only when the density of eggs reaches 1000/hectare. When would this density be reached as a result of population growth? Begin with the basic equation for growth: N t = N 0R t Now make the natural log transform of the equation: ln Nt = ln N0 + t ln R Using the starting (2003) data, ln 1000 = ln 160 + t ln 1.25 6.91 = 5.07 + .223t 1.84 = .223t t = 8.25 Since the rules say the density must be >1000, and gypsy moths reproduce once per year, spraying will only be permitted after 9 years of population growth, or in 2012. There is a second distinction that is needed: Some species reproduce only once, then adults die (semelparity). Other species reproduce repeatedly. That strategy is called iteroparity. Different mathematical models are needed for each of the four ‘categories’: 1) continuous semelparous reproduction – this produces the simple model of exponential reproduction you are used to seeing. 2) discrete semelparous reproduction 3) continuous iterparous reproduction – also can fit the simple model. 4) discrete iteroparous reproduction So, we need to consider what is called ‘discrete time’ models, meaning growth that occurs in bursts, and a mathematical form called ‘difference equations’. Let’s look at the way difference equations present ‘exponential’ growth… If populations are semelparous (reproduce once and adults die), then… Nt+1 = R0Nt This is the version you are used to seeing. This is the growth equation for single-celled organisms. The models are different when adults survive (i.e. seasonal reproduction in iteroparous organisms): Nt+1 = RNt + Nt or Nt+1 = (R + 1)Nt This need not look too different from the usual… Nt+1 = Nt With adult survival = (R+1). If we follow growth for a second generation… Nt+2 = Nt+1 = Nt = 2Nt And after T time steps (generations)… Nt+T = TNt Compare this to the usual solution for continuous growth… dN/dt = rNt Rearranging: dN/N = r dt Integrating over a time interval beginning at t=0 and ending at time = T… The left side of the equation integrates to: ln Nt – ln N0 And the right side to: rT – r0 = rT ‘Exponentiate’ each side of the equation, and you get the familiar result: Nt/N0 = erT = T or Nt = N0erT = N0T The two forms, continuous and discrete, are essentially similar, and = er. It is particularly important in discrete reproductive strategies (but really in all strategies) that we consider not only births, but also deaths between bouts of reproduction. Graphs of population size over time when reproduction occurs in bursts are typically ‘sawtoothed’, with rapid increase at the time of reproduction, then gradual decrease in the intervals between bouts of reproduction… That is the growth of a population of pheasants introduced onto Protection Island off the coast of the state of Washington. There was abundant food and were no predators; the island was far enough from mainland that there were no pheasant dispersers onto or off of the island. The size of the pheasant population was monitored annually after the introduction of 8 pheasants in 1937. Growth followed the sawtooth curve of the discrete exponential pattern. In 1938, the population was 30. That gives a of 30/8 or 3.75. From that we can predict the population to be expected if growth were exponential for each succeeding year. The predictions was pretty accurate for 1939 and 1940, but the predicted population for 1941 was larger than that actually observed. The population should have reached 5932, but was actually only about 1800. In 1942 predicted population size was 11,488, but actual size was 1898. These differences are considered to be indications of food limitation, and of logistic growth. We cannot learn what the carrying capacity was, since the American army made Protection Island a training center, and the soldiers got target practice, as well as meals of unexpected excellence, by driving the pheasant population extinct. Doubling Time A simple comparison of population growth rates comes from measuring the time it takes those populations to double in size. If growth is exponential, then: Nt/N0 = 2 = ert ln 2 = rt 0.693/r = t Exponential Growth in Invasive Species When invasive species enter a new environment, they frequently occur in the absence of controlling agents (predators, diseases, even competitors adapted to them). As a result, their growth, at least initially, is exponential. That was certainly the case for Lythrum salicaria (purple loosestrife) and the zebra mussel. Their effects on community structure and food chains was (and may remain) severe. The zebra mussel initially grew very rapidly. However, in the last few years its growth has slowed. It first was seen in the mid-1980s (though it probably arrived a year or two before being found), and reached extraordinary density in Lakes Erie and St.Clair by the mid 1990s. Adding Stochastic Fluctuations The exponential models described are both deterministic models. No matter how many times we 'run' these models, they produce the same result at any point in time. There is a single value of the y-variable, population size, for each x, the time. We know full well that the real world doesn't function like that. If we run an experiment many times and want to report the results, we would report a mean, x , and a measure of variation around that mean, typically the variance s2 or a standard deviation s. In describing population growth, how can we introduce stochastic variation into the model? There are two basic ways. The first suggests that the environment varies stochastically, influencing the growth rate. We report averages for population size and r… The equation for the variance in population size is more complicated, and depends a bit on the method of derivation. The most widely accepted way, shown in May (1974) results in this equation: Even without the messy details, there are a few things to take away from this equation: 1) the variation increases with time, since t appears twice in exponents, once in the 'growth term' e2rt and once in the 'variance term' est. Populations that are growing rapidly, or that have a highly variable growth rate show high variance in numbers; populations growing slowly or with little variation in r have low variance in numbers. Also, 2) populations which begin at larger size have a higher variance in numbers. If a population shows a high variance in numbers, there is a real danger that the population may decrease to extinction by chance. A very simple way to predict whether environmental stochasticity will drive a population to extinction was presented by May in estimating variance. If the variance in r is greater than twice the mean, i.e. s2r > 2r , the probability of extinction approaches 1. The other way in which variability in growth is described is to examine demographic stochasticity. In this view the effects of variability are evident in the births and deaths occurring within a population. Randomness in the sequence of events (births and deaths) that can produce remarkable and counterintuitive results. If birth rate and death rate are equal, then the deterministic model says the population remains fixed in size. The stochastic model says there is a variance in population size: 2N = 2N0bt If b and d are not equal, then the population is either growing or declining on average. If Nt is declining, then the population is headed for inevitable extinction. The more interesting question is what happens to a population which is growing on average, but which suffers from stochastic variation. Pielou (1969) has examined the long term prospects for species whose birth and/or death rates vary stochastically, and while the mathematics are complex, the results can be sorted into 3 cases: Case I - if birth rate is less than death rate, we evaluate population growth as time approaches infinity, and from that the probability of extinction over the long term - extinction is inevitable. Case II - b=d is the equilibrium situation, and a population would seem immune to extinction. Stochastic fluctuations in birth and death change this prediction. The algebraic solution gives a probability of extinction indicating extinction is again inevitable. lim t P(0)t = (bt/1+bt)n(0) 1 Case III - b>d - There is now a non-zero probability that even this population may go extinct. The probability of extinction, which variance indicates should include both the size of the starting population as well as birth and death rates, is, in the limit as time approaches infinity: If the population is growing on average (b is greater than d), then the probability of extinction declines as the initial population size increases. Chance events when the population is near its initial size are unlikely to carry it down to 0. Also, the greater the growth rate, evident in the ratio of death rate to birth rate, the less likely extinction is. Average growth rate increases, making recovery rapid, even from temporarily small populations. These are the characteristics of ‘weeds’. Briefly, let’s remember the graphical characteristics of exponential growth… Nt = N0ert If: And take the ln of both sides: ln Nt = lnN0 + rt Then a plot of numbers (in the form of natural logs) against time is a straight line with slope r and y-intercept lnN0. slope = r ln Nt ln N0 time Bibliography May, R.M. (1974) Ecosystem pattern in randomly fluctuating environments. Progress in Theoretical Ecology 3:1-50. Pielou, E.C. (1969) An Introduction to Mathematical Ecology. WileyInterscience, N.Y.