8_Homing and Trafficking
advertisement

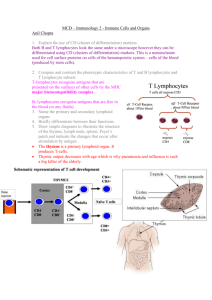
Lymphocyte Activation, Homing, and Trafficking B Cell Activation and Differentiation - Antigen transport to lymph node Lymphatic capillaries within tissue spaces constantly carry interstitial fluid to larger lymphatic vessels, which transport fluid, or lymph, to regional lymph nodes (LNs) The tissue spaces also contain professional APCs (i.e. dendritic cells, Langerhans cells) that engulf foreign antigens and present then to T cells once in the LNs Lymph carries free antigen particles (not engulfed by dendritic cells) to LNs, where they are then engulfed by B cells and presented to T cells - Once free antigen is brought to LN, naïve B cells specific to that antigen bind via surface antibodies, or BCRs IgM and IgD are the BCR antigen receptors of naïve B cells Although BCRs can recognize all types of antigens (i.e. proteins, carbohydrates, lipids), they can only present processed antigen protein peptides to T cells - BCR-bound antigen is endocytosed and processed (chopped up into small peptides) by B cells - Small peptides are loaded onto class II major histocompatibility complex (MHC II) molecules inside B cells and trafficked to the cell surface for presentation to a peptide-specific Th cell This function of B cells demonstrates their role as professional APCs Other professional APCs (dendritic cells, macrophages, basophils) also can only present peptides - MHC I and MHC II presentation helps the body determine when Th and/or Tc cell function is required MHC class II molecules present antigens to Th cells and leads to activation of Th cells MHC class I molecules present antigens to Tc cells and leads to activation of Tc cells - TCRs on a Th cell are specific to and recognize the antigenic peptide presented by B cell MHC II molecules - Following binding, the Th cell secreted cytokines - Under cytokine help from the Th cell, the B cell divides (proliferates) - Divided progeny differentiate into plasma cells and long-lived memory cells - Cytokines from the Th cell also induce class switching and affinity maturation in B cells B cell proliferation, differentiation, class switching and affinity maturation occur in germinal centers - The mechanisms for memory cell generation and survival are not well understood Memory B cells generated in lymph nodes are able to home and traffic through all secondary lymphoid organs (except MALT), where they can encounter and mount rapid response to a subsequent exposure of the same microbe or antigen - Most plasma cells exit the LNs where they were produced via efferent lymphatics and enter blood circulation. During circulation, they migrate and home to other lymph nodes, spleen, and bone marrow Plasma cells in lymph nodes and spleen only live for a couple of weeks, then die by apoptosis Plasma cells of bone marrow live and produce antibodies for years Antibodies get distributed in most body fluids throughout the body. They bind with antigens and microbes and opsonize them for their easy detection and removal by phagocytes Once B cells are stimulated in lymph nodes or spleen, effector B cells no longer go to MALT If naive B cells encounter antigen in LN and spleen, they don’t need to travel to MALT Tc Cell Activation and Differentiation - Dendritic cells engulf antigens within tissue spaces and bring them to LNs. There, antigens are broken down into small peptides that are then loaded onto MHC I and MHC II molecules of dendritic cells for presentation to Tc and Th cells, respectively Antigen presentation to T cells takes place in the paracortex region of the LNs - Antigen recognition leads to cytokine production by both the APC and antigen-specific T cell (either Tc or Th) Which APCs produce cytokines? Dendritic cells, macrophages - Cytokines cause activation of the T cell, which then divides and proliferates Progeny of the parent Tc cell differentiate into effector Tc cells and memory Tc cells o Effector Tc cell are known as cytotoxic T lymphocytes, or CTLs Progeny of parent Th cell differentiate into effector Th cells and memory Th cells - Long-lived memory Tc and Th cells generated in LNs survive for years and are able to home and traffic through secondary lymphoid organs (not MALT) and exit into non-lymphoidal tissue to patrol body organs - CTLs patrol the body for sites where infected cells are present (sites of inflammation) Infected cells express the same antigenic peptide associated with MHC I molecule CTLs recognize these cells by binding to the MHC I/antigenic peptide complex Following binding with the infected cells, CTLs secrete perforin and granzymes o Perforin facilitates entry of granzymes into the cell o Granzymes induce apoptosis of the infected host cell - Effector Th cells also patrol the body for sites of inflammation Their functions are mediated largely by secreted cytokines Immune cell types (i.e. macrophages) that have phagocytosed pathogens express the same antigenic peptide associated with the MHC II molecule that the Th cell can recognize TCRs on the Th cell bind to MHC II/antigenic peptide complex and secrete cytokines Cytokines are potent activators of macrophages and help them become more efficient pathogen killers Effector Th cells provide cytokine help to other cell types (i.e. B cells, Tc cells, and eosinophils) - Once the antigen/pathogen is eliminated, effector Tc and Th cells die by apoptosis Lymphocyte Activation and Differentiation in MALT - Antigens at mucosal surfaces are transported to GALT by specialized M (microfold) cells that are present in follicle associated epithelium (FAE) - M cells are flattened epithelial cells that lack microvilli. They take up pathogens/antigens from the intestinal lumen by endocytosis or phagocytosis, but do not kill them. The pathogenic material is transported through the interior of the cell, via vesicles, to the basal membrane, where it is released into the extracellular space by transcytosis - Antigen is taken up by dendritic cells and processed for T cell presentation. T cells proliferate and differentiate into memory and effector Th and Tc cells. Antigens are also picked up by B cells, which present them to Th cells to receive cytokine help. Activated B cells proliferate and differentiate into memory and plasma cells - Effector cells (plasma and effector T cells) exit via efferent lymphatics, are drained into mesenteric LNs, and eventually reach the thoracic duct - From the thoracic duct, effector cells circulate throughout the body in the bloodstream, only ever re-entering mucosal tissue (they are never redistributed to non-mucosal sites) - B cells differentiated in MALT produce characteristically high levels of IgA. Thus, migrated plasma cells in mucosal sites mainly secrete IgA class of antibody These antibodies are transported across epithelial cells and released as secretory IgA into the lumen where they block the entry of pathogens - Memory B and T cells strategically home to and traffic through MALT (but not through non-MALT tissues) Unlike plasma cells of the LNs and spleen, there is no evidence suggesting that those generated in MALT can survive for years anywhere in the body Lymphocyte Trafficking and Homing - Naïve lymphocytes do not show preference for a specific type of secondary lymphoid tissue but circulate indiscriminately to various types throughout the body - Lymphocyte homing to LNs and MALT is highly regulated Initial attachment of naïve lymphocytes to HEVs is mediated by the binding of specific molecules o In LNs, L-selectin on lymphocytes binds to GlyCAM-1 on HEVS GlyCAM-1 (glycan bearing cell adhesion molecule-1) o In MALT, L-selectin on lymphocytes binds to MAdCAM-1 on HEVS MAdCAM-1 (mucosal vascular addressin cell-adhesion molecule-1) Naïve lymphocytes exit these organs via efferent lymphatics to re-enter circulation - Lymphocyte homing to spleen is not as regulated as migration into lymph nodes and MALT Spleen does not contain HEVs Naïve lymphocytes are delivered to the marginal zone sinuses by a passive mechanism Naïve lymphocytes exit the spleen via the splenic v. - Following activation of naïve lymphocytes by antigen, newly-produced effector and memory lymphocytes gain expression of different molecules (no longer L-selectin). Because of this new expression, trafficking patterns of effector and memory lymphocytes differ from those of naïve lymphocytes - Memory and effector cells exhibit tissue-selective homing Effector cells tend to home to regions of infection by recognizing inflamed vascular endothelium Memory cell selectively home to the type of tissue in which they first encountered antigen (probably to ensure that cells return to tissues where they are most likely to re-encounter the same antigen) Only lymphocytes that encounter antigen and are activated in MALT gain the expression of α4β7 integrin (molecule that determines mucosa-specific homing) o α4β7 integrin binds to MAdCAM-1 on endothelial cells lining blood vessels of mucosal walls o Because MAdCAM-1 is not restricted to the blood vessels of one type of MALT, priming of lymphocytes in one mucosal tissue can induce protective immunity at other mucosal surfaces o MAdCAM-1 is only expressed at mucosal sites and α4β7 integrin cannot bind to GlyCAM-1 expressed at non-mucosal sites. Thus, lymphocytes activated in MALT only home to MALT o Lymphocytes differentiated in non-mucosal lymphoid tissues (i.e. spleen and LNs) express molecules other than α4β7 integrin and, thus, do not home to mucosal surfaces