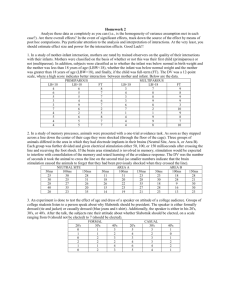
detecting conscious awareness from involuntary autonomic responses
advertisement

Detecting awareness from autonomic responses 1 DETECTING CONSCIOUS AWARENESS FROM INVOLUNTARY AUTONOMIC RESPONSES Ryan B. Scott D.Phil. 1, 4 Ludovico Minati MSc. 2, 5 Zoltan Dienes D.Phil. 1, 4 Hugo D. Critchley D.Phil. 2, 4 Anil K. Seth D.Phil. 3, 4 1 2 School of Psychology, University of Sussex Department of Psychiatry, Brighton & Sussex Medical School (BSMS) 3 4 5 School of Informatics, University of Sussex Sackler Centre for Consciousness Science, University of Sussex Scientific Department, Neuroradiology and Clinical Neurophysiology Units, Fondazione IRCCS Istituto Neurologico Carlo Besta, Milano, Italy. Corresponding author: Dr Ryan Scott Pevensey Building School of Psychology University of Sussex Falmer BN1 9QH Tel: +44 (0)1273 876650 Fax: +44 (0)1273 678058 Email: r.b.scott@sussex.ac.uk Detecting awareness from autonomic responses 2 Abstract Can conscious awareness be ascertained from physiological responses alone? We evaluate a novel learning-based procedure permitting detection of conscious awareness without reliance on language comprehension or behavioural responses. The method exploits a situation whereby only consciously detected violations of an expectation alter skin conductance responses (SCRs). 30 participants listened to sequences of piano notes that, without their being told, predicted a pleasant fanfare or an aversive noise according to an abstract rule. Stimuli were presented without distraction (attended), or while distracted by a visual task to remove awareness of the rule (unattended). A test-phase included occasional violations of the rule. Only participants attending the sounds reported awareness of violations and only they showed significantly greater SCR for noise occurring in violation, versus accordance, with the rule. Our results establish theoretically significant dissociations between conscious and unconscious processing and furnish new opportunities for clinical assessment of residual consciousness in patient populations. Keywords: Consciousness Conscious awareness Disorders of Consciousness Persistent Vegetative State Minimally Conscious State Locked-In Syndrome Unconscious knowledge Detecting awareness from autonomic responses 3 1. Introduction An important challenge for cognitive psychology, neuroscience, and clinical neurology is to determine whether conscious awareness can be detected through nonbehavioural responses. From a theoretical perspective, any method allowing the conscious status of knowledge to be assessed without the confounding influence of explicit subjective reports is of considerable value. More urgently, the assessment of consciousness in brain injured patients is central to the differential diagnosis of vegetative state, minimally conscious state, and locked-in syndrome. Accurate diagnosis is needed to inform prognosis and clinical management (Jennett, 2002). Current clinical practice relies heavily on behavioural evaluation and is therefore limited where volitional motor responses or language comprehension are compromised (Majerus, Gill-Thwaites, Andrews, & Laureys, 2005). These limitations contribute to rates of misdiagnosis, estimated to be 37-43% in patients diagnosed as vegetative state (Andrews, Murphy, Munday, & Littlewood, 1996; Childs, Mercer, & Childs, 1993; Schnakers et al., 2009). A variety of methods have been developed to aid evaluation of behaviourally unresponsive patients. Electrophysiological and nuclear medicine techniques have established power for predicting negative outcome (Carter & Butt, 2005; Daltrozzo, Wioland, Mutschler, & Kotchoubey, 2007). In contrast, detecting the presence of residual consciousness as a means to predict positive outcome, remains a substantial challenge. Cognitive theories and empirical research have motivated a number of candidate methods. One approach draws on an apparent awareness-dependence for trace versus delay conditioning. In trace conditioning the CS ends prior to the US, thus requiring a memory ‘trace’ for an association to be made, while in delay conditioning the CS and US overlap. Several studies have found that trace but not delay conditioning requires awareness of the stimuli and of the associative relationship (Knight, Nguyen, & Bandettini, 2006; Lovibond & Detecting awareness from autonomic responses 4 Shanks, 2002), though others have failed to observe this limitation to unconscious learning (Destrebecqz et al., 2010; Fu, Fu, & Dienes, 2008). Bekinschtein et al., (2009) employed trace conditioning of the eye-blink response evaluating anticipatory electromyographical responses as an indicator of learning. While they observed that the degree of learning was a good indicator of recovery in patients, the method failed to provide a clean separation of conscious and unconscious control subjects; some conscious controls failed to learn and one unconscious control showed marginal learning while under general anaesthetic. A related approach exploits cognitive event-related potentials (ERPs) to detect awareness of violations in temporal regularities. Bekinschtein et al. (2009) devised a paradigm with both local and global violations of an auditory regularity such that detecting the global differences required maintaining the perceptual representation over some seconds, which was held to require consciousness. Consistent with their predictions only the global violations generated a P300 ERP complex previously associated with conscious access (Sergent, Baillet, & Dehaene, 2005), and this effect was only present in participants reporting awareness of the global structure. However, the effect was only reliably observed in participants instructed to attend to the global regularity. The method is consequently dependent on verbal comprehension which is known to be impaired in a significant proportion of stroke and traumatic brain injury patients (Eisenberg et al., 1990; Inatomi et al., 2008). Finally, an alternative approach employs functional neuroimaging to index distinct patterns of regional brain activity associated with the content of intentional visual imagery. This method has provided evidence for conscious awareness in patients otherwise fulfilling the criteria for vegetative state (Monti et al., 2010; Owen et al., 2006). However, in addition to requiring brain-imaging equipment, the method is again reliant on verbal comprehension; patients must change their mental imagery in accordance with verbal instructions. Detecting awareness from autonomic responses 5 Despite these advances, there remains a need for an accurate method to determine conscious awareness, that is not reliant on motor responses or language comprehension, and which preferably can be applied at the bedside. To address this challenge we devised a novel approach which we term the Learned Aversive Contingency (LAC) procedure. The method exploits the skin conductance response (SCR) to index learning of a predictive relationship. Specifically, sequences of piano notes predicted either a pleasant fanfare or aversive white noise. Patients diagnosed as persistent vegetative state (PVS) are known to exhibit significant SCRs in response to white noise (Hildebrandt, Zieger, Engel, Fritz, & Bussmann, 1998; Keller, Hulsdunk, & Muller, 2007), making this a suitable clinical measure. The predictive sequences were designed to minimise the likelihood of unconscious learning. In common with previous attempts, a delay was inserted between the predictive sequence and the aversive or non-aversive stimulus. However, based on evidence that the proportion of unconscious learning reduces with longer delays (e.g. Gustav Kuhn & Dienes, 2005; G. Kuhn & Dienes, 2006) we inserted an extended pause of between 1 and 2 seconds during training and always 2 seconds during testing - four times that commonly used in trace conditioning. In addition, the predictive rule was abstract in nature; the pattern of similarity between the notes predicted the outcome but the pitch of the notes was different in every test-sequence. Implicit learning of this type of consistency, known as repetition structure, can be observed in paradigms such as artificial grammar learning (AGL), where its influence is mediated by feelings of familiarity (Scott & Dienes, 2008, 2010a; but not by fluency, Scott & Dienes, 2010b). However, the learning context in paradigms such as AGL is substantially different to that employed in the current study. Most notably, in AGL all elements of the stimuli are typically presented either simultaneously as a single visual unit, or as an uninterrupted stream. In the present auditory paradigm we aimed to reduce implicit learning by a Detecting awareness from autonomic responses 6 combination of the 2-second delay preceding the final element of each sequence and the use of a unique pitch on each trial. To measure conscious expectation, we exploited differences in SCR in response to the noise when its occurrence was predictable versus unpredictable. SCR magnitudes evoked by an aversive stimulus are greater when the stimulus is unanticipated (Ohman, 1971). Learning was therefore assessed by comparing SCR in test trials where the rule either applied as before or was violated such that the aversive noise was unexpected. Critically, we sought to devise a training sequence that made the rule sufficiently salient that individuals attending the sounds would detect it without instruction, thus avoiding dependence on verbal comprehension. We manipulated attention as the means to simulate patients who are either consciously aware or unaware of the auditory structure, thus permitting us to test the extent to which learning is dependent on awareness. While opinions differ as to whether consciousness and attention are doubly dissociable processes (Bussche, Hughes, Humbeeck, & Reynvoet, 2010; Koch & Tsuchiya, 2007), we merely exploited their uncontroversial possible association, and verified the effectiveness of the attention manipulation on conscious awareness of the rule using subjective reports. We evaluate the LAC procedure by contrasting reported awareness and SCR measurements in participants exposed to the sound sequences without distraction with those of participants engaged in a visual task removing attention from the sounds. Removing awareness in this way has important advantages over alternative approaches, such as the use of anaesthesia (cf. Bekinschtein, Shalom et al., 2009), because unlike those approaches attentional manipulation should not compromise other brain functions possibly preserved in patients. Detecting awareness from autonomic responses 7 2. Material and methods 2.1 Participants Thirty students (11 male, 19 female; age M = 23, SD = 4.1 years) participated in exchange for course credits or £5. All participants were naive to the experimental hypothesis and were randomly assigned to experimental condition (attended or unattended).1 2.2 Materials Sound sequences included 40 training trials and 65 test trials. Each trial consisted of three piano notes followed by a pause (initially 1 s, increasing to 2 s) and either a pleasant fanfare or a burst of white noise (100 dB, 1 s). Two trial types were presented during training: 1) non-aversive trials, consisting of three identical notes followed by the fanfare, and 2) aversive trials, where the third note was substantially different from the previous two and followed by white noise. Training included 15 aversive trials and 25 non-aversive trials arranged to facilitate uninstructed learning of a simple rule: Three identical notes (of any pitch) predicts the pleasant fanfare, whereas a different third note predicts the noise burst. The test phase consisted of eight blocks of eight trials with the first block preceded by one additional aversive trial. This first aversive trial was included solely to reacclimatise participants to the noise burst after the five minutes of silence following training; this trial was therefore not included in the analyses. Each of the eight blocks contained six nonaversive trials, one predictable aversive trial, and one unpredictable aversive trial, in pseudorandom order. In predictable aversive trials, the noise burst was preceded by a sequence in which the third note was different, as in training. In unpredictable aversive trials, the noise burst occurred after three identical notes, violating the rule. Learning would be apparent as 1 Twenty four participants were randomly assigned in equal numbers to the two conditions. To replace those not showing sufficient sympathetic response for learning assessment (see results), an additional six were recruited and randomly assigned in appropriate proportions. Detecting awareness from autonomic responses 8 larger magnitude SCRs for unpredictable compared with predictable trials (Figure 1). The pause between piano notes and fanfare or white noise was 2 s in all test trials. Full details of training and test sequences are given in Tables S1 and S2 of the supplementary material. Non-aversive trial Unpredictable aversive trial NOISE Predictable aversive trial NOISE SCR - attended condition Difference indicative of rule learning SCR - unattended condition Figure 1. Trial types and predicted SCR. In the attended condition, larger magnitude responses are expected for unpredictable as compared to predictable trials, indicating rule learning. In the unattended condition, this difference is expected to be abolished. 2.3 Procedure Participants were seated at a computer and equipped with headphones. Skin conductance was recorded from electrodes attached to the index and middle fingers of their left hand. Training sequences were followed by a five minute rest before the test sequences. The attended condition aimed to simulate conscious patients without motor volition or language comprehension. These participants were therefore asked to imagine they were hospital patients, unable to move or understand language. Care was taken not to provide any verbal or other instruction. The unattended condition aimed to simulate patients lacking conscious awareness of the stimuli. These participants were therefore asked to perform a visual discrimination task as accurately as possible while ignoring the sounds. The visual Detecting awareness from autonomic responses 9 task involved using eye-movements to indicate the position and orientation of a line appearing on screen (see Figure S1 in the supplementary material). 3. Results Skin conductance was measured with a sensitivity of 0-100 µS, the signal sampled at 500 Sa/s 16-bit, low-pass filtered at 5 Hz, and detrended with a fifteen degree polynomial. For each test-trial, the amplitude of SCR evoked by the fanfare or noise burst was determined as the difference between the maximum in the 0-8 s post-stimulus interval and the average in the 1-3 s pre-stimulus interval. Statistical analysis was performed individually for each participant. First, SCRs with amplitude beyond three standard deviations from the mean for each trial type were excluded as outliers; there were 36 outliers in total, no more than two for any participant, and all were for non-aversive trials.2 Second, a two-sample t-test was performed to verify whether SCR amplitude was greater for noise bursts than fanfares; since the absence of this difference would indicate insufficient sympathetic response to use SCR as a measure of learning, such participants were excluded from further assessment. This represents a practical constraint but also ensures that in clinical application compromised autonomic responsivity would not result in a false negative assessment. In the remaining participants, a paired one-tailed t-test assessed whether SCR magnitudes were greater for the eight unpredictable compared with the eight predictable aversive trials. Upon completion of the experiment, each participant in the attended condition reported learning the rule during the training phase and realizing that it was occasionally violated 2 This pattern is to be expected given that there were more non-aversive than aversive trials (48 vs. 16) and that the magnitude of SCRs elicited is smaller for non-aversive than aversive trials. As such, any compromised response, for example resulting from the participant coughing, will have been more likely to occur during a nonaversive trial and would also be more likely to result in an SCR magnitude outside the normal range for nonaversive than aversive trials. Detecting awareness from autonomic responses 10 during the test phase. No participant in the unattended condition reported awareness of the relationship between the piano notes and subsequent sounds; as intended, the visual distraction task eliminated conscious awareness of the auditory structure. Six participants (one attended condition, five unattended condition) failed to show significantly greater SCR magnitudes to noise bursts than fanfares and were hence excluded from the subsequent learning assessment. 40 4 * Group Means * Participant mean greater than zero p < .05 30 3 + Participant mean greater than zero p < .10 MeanSCR GSRDifference Difference(μS) Mean * * 20 2 * * 10 1 + * * * * * * 00 -10 -1 -20 -2 -30 -3 Attended Condition Unattended Condition (7 female, 5 male) (7 female, 5 male) Figure 2. Mean difference in SCR magnitude for unpredictable minus predictable aversive trials, shown for each participant grouped by condition. Error bars indicate standard error of the mean. For the remaining twenty four participants (12 per condition) the differences in SCR magnitudes for unpredictable minus predictable aversive trials are shown in Figure 2. For the Detecting awareness from autonomic responses 11 attended condition, SCR magnitudes were larger for unpredictable as compared to predictable aversive trials (M = 20.9, SD = 10.8 µS vs. M = 11.2, SD = 7.8 µS). This effect was significant at the individual level for eleven participants (p < .05) and marginal for one (p = .071). In the unattended condition, SCR magnitudes were not larger for unpredictable compared with predictable trials (M = 4.9, SD = 6.2 µS vs. M = 6.6, SD = 8.3 µS); the effect was not significant at the individual level for any participant (all p > .13; 95% CI of the difference -3.4, +1.0 µS, ruling out population differences even remote from the attended mean difference of 10.9 µS). It was possible that the visual task could have influenced SCR magnitudes in some way other than by the intended influence on awareness of the auditory structure. To test for this we compared the magnitude of the SCRs for each type of test trial, between attended and unattended conditions, see Figure 3. The magnitudes did not differ between groups for nonaversive trials, t(22) = .94, p = .356, or predictable aversive trials, t(22) = 1.03, p = .314, but did so for unpredictable aversive trials, t(22) = 4.79, p < .001, where, as predicted based on the violation of expectations, SCR magnitudes were significantly greater for the attended condition. There is hence no evidence that the visual task affected SCRs by a means other than the intended influence on awareness. Detecting awareness from autonomic responses 12 3.0 Unattended Attended 2.5 Mean SCR (μS) 2.0 1.5 1.0 0.5 0.0 Non-aversive Predictable aversive Unpredictable aversive Figure 3. Mean SCR magnitudes for each type of test trial contrasted between attended and unattended conditions. Error bars indicate standard error of the mean. The results demonstrate that only participants who had conscious awareness of the rule showed a significantly greater SCR magnitude when it was violated. Participants in the unattended condition, all of whom reported being unaware of the rule, showed equivalent SCR magnitudes after white noise irrespective of whether it occurred in accordance or violation of the rule. It is not possible to estimate the extent to which the observed absence of implicit learning was due to any specific feature of the paradigm. However, the results are consistent with both the inclusion of an extended pause prior to the predicted stimulus and the use of different pitches on each trial being effective in reducing unconscious learning. Detecting awareness from autonomic responses 13 4. Discussion The results confirm that 1) the LAC procedure enabled reliable learning of an abstract relation in the absence of any instruction, 2) conscious awareness of the relation was important for learning to occur, and 3) learning was detectable from autonomic responses. The procedure thus provides a means to detect conscious awareness based solely on an autonomic response and avoids reliance on language comprehension or motor responses. The technical simplicity of the LAC procedure ensures that it could be easily implemented at the bedside, inviting consideration of its clinical potential. Our results suggest the method would have a high diagnostic specificity; i.e., it would provide strong evidence for the presence of conscious awareness. This is supported by the fact that removing awareness of the contingency by manipulating attention was sufficient to eliminate learning despite cognitive function being otherwise intact. The procedure should also provide higher sensitivity than existing measures because it does not require language comprehension or motor responses. Potential limitations include inadequate cognitive or attentional capacity, which may occur in patients with extensive cortical damage, and compromised autonomic responsivity impairing SCR. While SCRs evoked by white noise are observed in PVS patients (Hildebrandt, Zieger, Engel, Fritz, & Bussmann, 1998; Keller, Hulsdunk, & Muller, 2007) the procedure also embeds a test for these responses, thus allowing identification of patients for whom assessment would be inconclusive. Relevant to both specificity and sensitivity, a recent study found that coma patients have preserved SCR to emotionally salient stimuli (Daltrozzo et al., 2010). The presence of SCR in presumably unconscious patients emphasises the robustness, and hence suitability, of this response, while also highlighting the need to test for learning of an abstract relation to provide specificity. The absence of a reliable SCR to the white noise observed for five participants in the non-attending condition may in part be due to these participants being required to actively Detecting awareness from autonomic responses 14 focus their attention on the visual task. This reduced responsivity would not be expected in conscious patients as it is unlikely they would deliberately focus their attention on something other than the sound sequences. The term consciousness has several meanings (Rosenthal, 2005). For example, there is a distinction between creature consciousness and mental state consciousness. Our method bears only on the latter. That is, our aim is to answer the question: Does the person being assessed have any conscious mental states? By virtue of the empirical reliance that we have demonstrated, namely that under the conditions of our test conscious knowledge is required for learning to be apparent, learning on our test can be taken as evidence for the existence of conscious knowledge. A negative result allows no conclusion as to whether or not the person has any conscious mental states. During development of our procedure a variety of experimental factors were found to influence the reliability with which attending participants acquired the rule. Multiple revisions of the training sequences were necessary to overcome an apparent contingency blindness, whereby seemingly obvious relations were missed in the absence of direct instruction. This phenomenon has parallels with other surprising examples of cognitive opacity including change blindness and inattentional blindness (Simons & Chabris, 1999; Simons & Rensink, 2005) and is it itself worthy of further research. For example, such research could investigate more generally how dissociations between conscious and unconscious processing depend on task instructions and participant expectations, as well as on the details of the stimuli. A summary of all factors observed to influence learning is provided in the supplementary material available online. Detecting awareness from autonomic responses 15 5. Conclusion We have demonstrated how learning of an aversive contingency, assessed by an autonomic response, can be employed to reliably identify conscious awareness independently of language comprehension or motor volition. The LAC procedure advances our understanding of dissociations between conscious and unconscious processing in the absence of explicit instruction. It also has the potential to substantially enhance the clinical assessment of conscious awareness in brain injured patients. 6. Acknowledgements The authors are grateful to Paolo Cortellazzi, MD, for insightful feedback on an earlier version of the manuscript. RS and ZD were supported by an ESRC Project Grant. LM was supported by a Wellcome Trust Programme Grant to HDC. Participant funding was provided by the Dr Mortimer and Theresa Sackler Foundation. Detecting awareness from autonomic responses 16 7. References Andrews, K., Murphy, L., Munday, R., & Littlewood, C. (1996). Misdiagnosis of the vegetative state: Retrospective study in a rehabilitation unit. British Medical Journal, 313(7048), 13-16. Bekinschtein, T. A., Dehaene, S., Rohaut, B., Tadel, F. O., Cohen, L., & Naccache, L. (2009). Neural signature of the conscious processing of auditory regularities. Proceedings of the National Academy of Sciences of the United States of America, 106(5), 1672-1677. Bekinschtein, T. A., Shalom, D. E., Forcato, C., Herrera, M., Coleman, M. R., Manes, F. F., et al. (2009). Classical conditioning in the vegetative and minimally conscious state. Nature Neuroscience, 12(10), 1343-U1176. Bussche, E. V. d., Hughes, G., Humbeeck, N. V., & Reynvoet, B. (2010). The relation between consciousness and attention: An empirical study using the priming paradigm. Consciousness and Cognition, 19(1), 86-97. Carter, B. G., & Butt, W. (2005). Are somatosensory evoked potentials the best predictor of outcome after severe brain injury? A systematic review. Intensive Care Medicine, 31(6), 765-775. Childs, N. L., Mercer, W. N., & Childs, H. W. (1993). Accuracy of Diagnosis of Persistent Vegetative State. Neurology, 43(8), 1465-1467. Daltrozzo, J., Wioland, N., Mutschler, V., & Kotchoubey, B. (2007). Predicting coma and other low responsive patients outcome using event-related brain potentials: A metaanalysis. Clinical Neurophysiology, 118(3), 606-614. Daltrozzo, J., Wioland, N., Mutschler, V., Lutun, P., Calon, B., Meyer, A., et al. (2010). Emotional electrodermal response in coma and other low-responsive patients. Neuroscience Letters, 475(1), 44-47. Detecting awareness from autonomic responses 17 Destrebecqz, A., Perruchet, P., Cleeremans, A., Laureys, S., Maquet, P., & Peigneux, P. (2010). The influence of temporal factors on automatic priming and conscious expectancy in a simple reaction time task. Quarterly Journal of Experimental Psychology, 63(2), 291-309. Eisenberg, H. M., Gary, H. E., Aldrich, E. F., Saydjari, C., Turner, B., Foulkes, M. A., et al. (1990). Initial CT Findings in 753 Patients with Severe Head-Injury - A Report from the NIH Traumatic Coma Data-Bank. Journal of Neurosurgery, 73(5), 688-698. Fu, Q. F., Fu, X. L., & Dienes, Z. (2008). Implicit sequence learning and conscious awareness. Consciousness and Cognition, 17(1), 185-202. Hildebrandt, H., Zieger, A., Engel, A., Fritz, K. W., & Bussmann, B. (1998). Differentiation of autonomic nervous activity in different stages of coma displayed by power spectrum analysis of heart rate variability. European Archives of Psychiatry and Clinical Neuroscience, 248(1), 46-52. Inatomi, Y., Yonehara, T., Omiya, S., Hashimoto, Y., Hirano, T., & Uchino, M. (2008). Aphasia during the acute phase in ischemic stroke. Cerebrovascular Diseases, 25(4), 316-323. Jennett, B. (2002). The vegetative state: medical aspects, ethical and legal dilemmas. Cambridge, England: Cambridge University Press. Keller, I., Hulsdunk, A., & Muller, F. (2007). The influence of acoustic and tactile stimulation on vegetative parameters vegetative state and EEG in persistent. Functional Neurology, 22(3), 159-163. Knight, D. C., Nguyen, H. T., & Bandettini, P. A. (2006). The role of awareness in delay and trace fear conditioning in humans. Cognitive, Affective, & Behavioral Neuroscience, 6(2), 157-162. Detecting awareness from autonomic responses 18 Koch, C., & Tsuchiya, N. (2007). Attention and consciousness: two distinct brain processes. Trends in Cognitive Sciences, 11(1), 16-22. Kuhn, G., & Dienes, Z. (2005). Implicit Learning of Nonlocal Musical Rules: Implicitly Learning More Than Chunks. Journal of Experimental Psychology: Learning, Memory, and Cognition, 31(6), 1417-1432. Kuhn, G., & Dienes, Z. (2006). Differences in the types of musical regularity learnt in incidental- and intentional-learning conditions. Quarterly Journal of Experimental Psychology, 59(10), 1725-1744. Lovibond, P. F., & Shanks, D. R. (2002). The role of awareness in Pavlovian conditioning: Empirical evidence and theoretical implications. Journal of Experimental Psychology: Animal Behavior Processes, 28(1), 3-26. Majerus, S., Gill-Thwaites, H., Andrews, K., & Laureys, S. (2005). Behavioral evaluation of consciousness in severe brain damage. Boundaries of Consciousness: Neurobiology and Neuropathology, 150, 397-413. Monti, M. M., Vanhaudenhuyse, A., Coleman, M. R., Boly, M., Pickard, J. D., Tshibanda, L., et al. (2010). Willful Modulation of Brain Activity in Disorders of Consciousness. New England Journal of Medicine, 10.1056/NEJMoa0905370. . Ohman, A. (1971). Interaction between instruction-induced expectancy and strength of unconditioned stimulus in GSR conditioning. Journal of Experimental Psychology, 88(3), 384-390. Owen, A. M., Coleman, M. R., Boly, M., Davis, M. H., Laureys, S., & Pickard, J. D. (2006). Detecting awareness in the vegetative state. Science, 313(5792), 1402-1402. Rosenthal, D. M. (2005). Consciousness and Mind. Oxford: Clarendon Press. Detecting awareness from autonomic responses 19 Schnakers, C., Vanhaudenhuyse, A., Giacino, J., Ventura, M., Boly, M., Majerus, S., et al. (2009). Diagnostic accuracy of the vegetative and minimally conscious state: Clinical consensus versus standardized neurobehavioral assessment. BMC Neurology, 9. Scott, R. B., & Dienes, Z. (2008). The conscious, the unconscious, and familiarity. Journal of Experimental Psychology-Learning Memory and Cognition, 34(5), 1264-1288. Scott, R. B., & Dienes, Z. (2010a). Knowledge applied to new domains: The unconscious succeeds where the conscious fails. Consciousness and Cognition, 19, 391-398. Scott, R. B., & Dienes, Z. (2010b). Fluency does not express implicit knowledge of artificial grammars. Cognition, 114(3), 372-388. Sergent, C., Baillet, S., & Dehaene, S. (2005). Timing of the brain events underlying access to consciousness during the attentional blink. Nature Neuroscience, 8(10), 1391-1400. Simons, D. J., & Chabris, C. F. (1999). Gorillas in our midst: Sustained inattentional blindness for dynamic events. Perception, 28(9), 1059-1074. Simons, D. J., & Rensink, R. A. (2005). Change blindness: Past, present, and future. Trends in Cognitive Sciences, 9(1), 16-20. Detecting awareness from autonomic responses 20 SUPPLEMENTAL MATERIALS Detecting awareness from autonomic responses 21 Table S1. Training trial sequences. Inter-trial Trial No. Note 1 Pause Note 2 Pause Note 3 Pause Stimulus pause 1 C3 100ms C3 100ms C3 1000ms fanfare 2500ms 2 C3 100ms C3 100ms C3 1000ms fanfare 2500ms 3 C3 100ms C3 100ms C3 1000ms fanfare 2500ms 4 C3 100ms C3 100ms D#1 1000ms white noise 2500ms 5 C3 100ms C3 100ms C3 1000ms fanfare 2500ms 6 C3 100ms C3 100ms D#1 1000ms white noise 2500ms 7 C3 100ms C3 100ms C3 1000ms fanfare 2500ms 8 C3 100ms C3 100ms C3 1000ms fanfare 2500ms 9 C3 100ms C3 100ms D#1 1000ms white noise 2500ms 10 C3 100ms C3 100ms C3 1000ms fanfare 10000ms 11 D4 100ms D4 100ms D4 1000ms fanfare 2500ms 12 D4 100ms D4 100ms D4 1000ms fanfare 2500ms 13 D4 100ms D4 100ms F#1 1000ms white noise 2500ms 14 D4 100ms D4 100ms D4 1000ms fanfare 2500ms 15 D4 100ms D4 100ms F#1 1000ms white noise 10000ms 16 F#3 100ms F#3 100ms F#3 1000ms fanfare 2500ms 17 F#3 100ms F#3 100ms G2 1000ms white noise 2500ms 18 F#3 100ms F#3 100ms F#3 1000ms fanfare 2500ms 19 F#3 100ms F#3 100ms G2 1000ms white noise 2500ms Detecting awareness from autonomic responses 22 20 F#3 100ms F#3 100ms F#3 1000ms fanfare 10000ms 21 B3 100ms B3 100ms G#2 1050ms white noise 2675ms 22 B3 100ms B3 100ms B3 1100ms fanfare 2850ms 23 B3 100ms B3 100ms G#2 1150ms white noise 3025ms 24 B3 100ms B3 100ms G#2 1200ms white noise 3200ms 25 B3 100ms B3 100ms B3 1250ms fanfare 10000ms 26 G5 100ms G5 100ms G5 1300ms fanfare 3550ms 27 G5 100ms G5 100ms C#3 1350ms white noise 3725ms 28 G5 100ms G5 100ms C#3 1400ms white noise 3900ms 29 G5 100ms G5 100ms G5 1450ms fanfare 4075ms 30 G5 100ms G5 100ms G5 1500ms fanfare 10000ms 31 G2 100ms G2 100ms G2 1550ms fanfare 4425ms 32 F5 100ms F5 100ms F5 1600ms fanfare 4600ms 33 C2 100ms C2 100ms C2 1650ms fanfare 4775ms 34 C#3 100ms C#3 100ms D1 1700ms white noise 4950ms 35 F#1 100ms F#1 100ms F#1 1750ms fanfare 10000ms 36 A3 100ms A3 100ms A3 1800ms fanfare 5300ms 37 E5 100ms E5 100ms E5 1850ms fanfare 5475ms 38 E5 100ms E5 100ms C2 1900ms white noise 5650ms 39 B4 100ms B4 100ms B4 1950ms fanfare 5825ms 40 D#2 100ms D#2 100ms C1 2000ms white noise - Detecting awareness from autonomic responses 23 Note. The piano notes are written with the octave number. For example, D#2 refers to the note ‘D sharp’ played in the second octave. The duration of the training phase was approximately 5 minutes. The piano notes and the stimuli (fanfare or white noise) were played to different ears; in the first 20 trials the piano notes were played to the left ear and the stimuli to the right ear, for the remaining 20 trials this was reversed. Detecting awareness from autonomic responses 24 Table S2. Test trial sequences. InterBlock Trial Note Note Pause No. No. 1 Note Pause 2 Type of Pause Stimulus trial 3 aversive trial pause - 1 D#2 100ms D#2 100ms C1 2000ms white noise 14000ms Block 1 2 D#4 100ms D#4 100ms D#4 2000ms fanfare 14000ms 3 D#5 100ms D#5 100ms D#5 2000ms fanfare 14000ms 4 G#3 100ms G#3 100ms G#3 2000ms fanfare 14000ms 5 A#4 100ms A#4 100ms D2 2000ms white noise 14000ms 6 C2 100ms C2 100ms C2 2000ms fanfare 14000ms 7 B5 100ms B5 100ms B5 2000ms fanfare 14000ms 8 D#1 100ms D#1 100ms D#1 2000ms white noise 14000ms 9 B3 100ms B3 100ms B3 2000ms fanfare 14000ms 10 F4 100ms F4 100ms F4 2000ms fanfare 14000ms 11 D#5 100ms D#5 100ms G3 2000ms white noise 14000ms 12 F#5 100ms F#5 100ms F#5 2000ms fanfare 14000ms 13 A#2 100ms A#2 100ms A#2 2000ms fanfare 14000ms 14 F#2 100ms F#2 100ms F#2 2000ms fanfare 14000ms 15 F#4 100ms F#4 100ms F#4 2000ms white noise 14000ms 16 C#4 100ms C#4 100ms C#4 2000ms fanfare 14000ms 17 G3 100ms G3 100ms G3 2000ms fanfare 14000ms 18 D#2 100ms D#2 100ms D#2 2000ms fanfare 14000ms 19 G1 100ms G1 100ms G1 2000ms fanfare 14000ms Block 2 Block 3 Predictable Predictable Unpredictable Predictable Unpredictable Detecting awareness from autonomic responses 25 Block 4 Block 5 20 F5 100ms F5 100ms F5 2000ms fanfare 14000ms 21 E1 100ms E1 100ms E1 2000ms white noise 14000ms 22 C#1 100ms C#1 100ms C#1 2000ms fanfare 14000ms 23 A3 100ms A3 100ms A3 2000ms fanfare 14000ms 24 F3 100ms F3 100ms F3 2000ms fanfare 14000ms 25 F4 100ms F4 100ms B2 2000ms white noise 14000ms 26 C#2 100ms C#2 100ms C#2 2000ms fanfare 14000ms 27 C#3 100ms C#3 100ms C#3 2000ms fanfare 14000ms 28 A4 100ms A4 100ms A4 2000ms fanfare 14000ms 29 B1 100ms B1 100ms B1 2000ms fanfare 14000ms 30 A3 100ms A3 100ms G1 2000ms white noise 14000ms 31 E3 100ms E3 100ms E3 2000ms fanfare 14000ms 32 D3 100ms D3 100ms D3 2000ms fanfare 14000ms 33 B4 100ms B4 100ms B4 2000ms white noise 14000ms 34 A1 100ms A1 100ms A1 2000ms fanfare 14000ms 35 G#5 100ms G#5 100ms G#5 2000ms fanfare 14000ms 36 D2 100ms D2 100ms D2 2000ms fanfare 14000ms 37 G#4 100ms G#4 100ms G#4 2000ms white noise 14000ms 38 A2 100ms A2 100ms A2 2000ms fanfare 14000ms 39 C#5 100ms C#5 100ms C#5 2000ms fanfare 14000ms 40 G#2 100ms G#2 100ms E1 2000ms white noise 14000ms 41 C5 100ms C5 100ms C5 2000ms fanfare 14000ms Unpredictable Predictable Predictable Unpredictable Unpredictable Predictable Detecting awareness from autonomic responses 26 Block 6 Block 7 Block 8 42 C1 100ms C1 100ms C1 2000ms fanfare 14000ms 43 G4 100ms G4 100ms G4 2000ms fanfare 14000ms 44 E2 100ms E2 100ms E2 2000ms fanfare 14000ms 45 A#4 100ms A#4 100ms A#4 2000ms white noise 14000ms 46 G#1 100ms G#1 100ms G#1 2000ms fanfare 14000ms 47 G#2 100ms G#2 100ms G#2 2000ms fanfare 14000ms 48 B2 100ms B2 100ms B2 2000ms fanfare 14000ms 49 E4 100ms E4 100ms A1 2000ms white noise 14000ms 50 A5 100ms A5 100ms A5 2000ms fanfare 14000ms 51 D1 100ms D1 100ms D1 2000ms fanfare 14000ms 52 A#1 100ms A#1 100ms A#1 2000ms white noise 14000ms 53 F1 100ms F1 100ms F1 2000ms fanfare 14000ms 54 A#3 100ms A#3 100ms A#3 2000ms fanfare 14000ms 55 D#3 100ms D#3 100ms D#3 2000ms fanfare 14000ms 56 G4 100ms G4 100ms A#2 2000ms white noise 14000ms 57 D5 100ms D5 100ms D5 2000ms fanfare 14000ms 58 F#1 100ms F#1 100ms F#1 2000ms fanfare 14000ms 59 B2 100ms B2 100ms C#1 2000ms white noise 14000ms 60 C4 100ms C4 100ms C4 2000ms fanfare 14000ms 61 A#5 100ms A#5 100ms A#5 2000ms fanfare 14000ms 62 E4 100ms E4 100ms E4 2000ms fanfare 14000ms 63 F2 100ms F2 100ms F2 2000ms white noise 14000ms Unpredictable Predictable Unpredictable Predictable Predictable Unpredictable Detecting awareness from autonomic responses 27 64 G2 100ms G2 100ms G2 2000ms fanfare 14000ms 65 E5 100ms E5 100ms E5 2000ms fanfare 14000ms Note. The piano notes are written with the octave number, for example D#2 refers to the note ‘D sharp’ played in the second octave. The duration of the test phase was approximately 20 minutes, starting 5 minutes after the training phase had completed. The delay after training was included to reduce participants’ habituation to the aversive noise and the consequent reduction in their SSR. The first trial – a predictable aversive trial – is included to reacclimatise participants to the white noise after the delay; it is not included in the analyses. The piano notes and the stimuli (fanfare or white noise) were played to different ears. In blocks 1-4 the piano notes were played to the left ear and the stimuli to the right ear, in the remaining blocks this was reversed. Detecting awareness from autonomic responses 28 Visual Discrimination Task The visual task was employed for the purpose of distraction in the unattended condition. The task required participants to discriminate between four potential stimuli occurring in random order at a rate of 1 Hz. The stimuli consisted of a 1 cm line oriented either vertically or horizontally and positioned at a vertically central position either towards the left or right side of the screen. In order to remove the potentially confounding effect of movement artefacts on skin conductance measurements, responses were given using eye movements. A sham eye-tracking camera was positioned on top of the screen, and participants were told that this would record their eye-movements. They were instructed that if a vertical line appeared on the right they were to briefly fixate the cross located at the top of the screen, and if a horizontal line appeared on the left they were to briefly fixate the cross located at the bottom of the screen, see Figure 1 below. For the two remaining alternatives, a vertical line on the left or a horizontal line on the right, participants were instructed to continue looking at the centre of the screen. Right and Vertical + + Left and Horizontal Figure S1. Example of the screen layout used for the visual discrimination task employed in the unattended condition. Detecting awareness from autonomic responses 29 Experimental factors influencing rule acquisition During development of the training sequence a variety of experimental factors were found to influence the reliability with which participants acquired the rule. While these factors were not systematically explored, the following provides a summary of those identified to have a substantial influence together with the insights acquired from interviews with participants of the pilot studies. Richness of the stimuli. Piano notes were more effective than pure tones. Participants’ comments indicated that this was based on the greater ease with which the pitch of piano notes could be distinguished. Delay between notes. A short delay (100 ms) between notes was found to be preferable to longer delays (300+ ms). Participants’ reports suggest that the shorter delay inclined them towards processing the pattern of three notes as a single predictive stimulus rather than as separate notes. Delay between the note pattern and subsequent stimulus. While there is a two second delay between the piano notes and the subsequent stimulus during testing, a delay of this length inhibited initial learning of the rule. Participants’ comments suggest that this effect was due to a longer delay reducing awareness that the notes and subsequent stimulus formed a single trial. This was addressed by having an initial delay of one second that incrementally increased to two seconds by the end of the training phase. Delay between trials. During testing the delay between trials was 14 seconds in order to provide a suitable window for monitoring the sympathetic skin responses. However, delays of this length during training reduced learning. An initially short delay of two and a half seconds was ultimately found to be effective. Participants’ comments indicated that the Detecting awareness from autonomic responses 30 longer delay reduced the likelihood that they would contrast one trial with the next and consequently notice the predictive pattern. Note repetition. Initial pilots included just two notes, with the second note either the same or different from the first. However, learning was found to be more reliable if there were three notes, with the final note being either the same or different. Participants indicated that the repetition caused more anticipation of the final note thus making any difference more salient. Elimination of irrelevant variations. Variations in the stimuli that were not relevant to the rule, particularly early in training, were found to substantially reduce learning. This most obviously applied to note pitch. The predictive rule is independent of pitch i.e. three notes the same irrespective of pitch predicts the fanfare, while two notes the same followed by one of a different pitch predicts the noise burst. However, reliable learning of this rule was only achieved when the training sequence was structured in stages. It was found to be necessary for initial blocks of training trials to differ only in whether the last note was the same or different, and not in pitch. This was repeated in subsequent blocks with each block employing a different pitch. Only at the end of the training phase was a different pitch used on each trial. When variations in pitch were introduced too early in training, participants were found to generate incorrect inferences about those changes and fail to identify the intended difference relating to the last of the three notes. A similar difficulty was identified with the ‘different’ third note. In early pilots the different note could be either higher or lower than the previous two notes. However, this was again found to reduce learning of the rule, with participants being inclined to hypothesise about consecutive trials showing the same or different direction of pitch change. In the final sequence this is avoided by the ‘different’ third note always being lower than the previous two. It is always lower by greater than one full octave to ensure the direction of the difference is clear. Detecting awareness from autonomic responses 31 A fixed trial format. Initial pilots included a noise burst on aversive trials, where the third note was different, and an equivalent period of silence on the non-aversive trials, where the three notes were the same. However, it was found to be beneficial to include a ‘positive’ sound on non-aversive trials in place of the silence. Participants indicated that this made it clearer that there was a fixed trial format, namely three notes followed by either a positive or negative sound. Various positive sounds were piloted, including applause, a fanfare, and a positive ‘ting’. The ‘fanfare’ was found to be most readily recognised as indicating a positive outcome. Delays between blocks of training trials. It was found to be beneficial to include periodic pauses between blocks of training trials. These were inserted between blocks employing the same note frequencies. Participants indicated that these pauses inclined them to refocus attention on the task. If they were yet to identify any rules in the pattern the pauses provided time for reflection and encouraged them to start afresh on the next block. Interaural separation. It was apparent that despite the cues present in the stimuli sequence, a small proportion of participants in the pilot studies failed to recognise that each trial consisted of a predictive pattern (three notes) followed by a predicted event (noise burst or fanfare). It was found that playing the two parts to separate ears increased the salience of this conceptual separation.