plant ion transport
advertisement

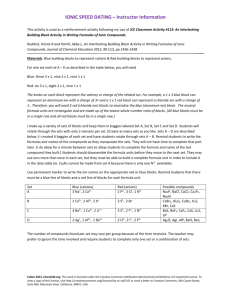
Plant Ion Transport Secondary article Rainer Hedrich, Bayerische Julius-Maximilians-University, Würzburg, Germany M Rob G Roelfsema, Bayerische Julius-Maximilians-University, Würzburg, Germany Article Contents . How to Survive in Pond Water? . Life on Earth Plant ion transport plays a key role in major physiological processes, such as nutrient uptake and redistribution, movement, growth and microbe interaction. Changes in the activity and density of ion-pumps, channels and carriers represent essential membrane-delimited steps in these processes. . Nutrient Uptake . Carbon is Absorbed from the Atmosphere . Long-distance Ion Transport . Growth and Tropisms . Living in a Community . Ion Transport in Progress How to Survive in Pond Water? After eukaryotic organisms were invaded by cyanobacteria, the first primitive unicellular algae were formed. At this point, these creatures became photoautotrophic and took advantage of the CO2-rich atmosphere to generate their own carbohydrates. New habitats could be colonized that were previously unsuitable for growth. The availability of nutrients in these environments differed tremendously from the ‘Jurassic pond’ in which life had started. To sustain growth, nutrients had to be captured from a habitat that could be as poor in ion-content as pond water. Plants developed mechanisms to survive in these hostile environments and learned to concentrate essential nutrients from their surrounding. Most nutrients are taken up by plants as ions, with the exception of carbon. Carbon is taken up as CO2 by most plants, although some water plants take advantage of the high concentration of HCO32 available in their aqueous environment. During photosynthesis, CO2 is converted into low-molecular mass carbohydrates that form the basis of cellular components. The synthesis of proteins and nucleic acids, however, also requires the uptake of inorganic nitrogen, phosphate and sulfate. These ‘macronutrients’ are mainly taken up in their oxidized anionic form, but nitrogen is also taken up as NH41 . Anion uptake is charge-balanced by the uptake of cationic macronutrients, such as K 1 , Ca2 1 and Mg2 1 . Metal ions such as those of Fe, Mn, Cu and Zn are taken up only in limited amounts (micronutrients) and incorporated in the prosthetic groups of enzymes. Life on Earth After colonizing land, higher plants faced problems caused by their immobility. Strategies evolved to overcome limitations in nutrient availability and water supply, yielding plants that are characterized by heterotrophic roots and photoautotrophic shoots. Solvent and solute uptake by the root is separated from CO2 assimilation in the shoot. As a consequence, a complex network for transport of ions, water and photoassimilates had to be formed. The long-distance transport pathway from the roots to the shoot and vice versa comprises the xylem and the phloem. The xylem transports mainly water and ions from the roots to the leaves, while the phloem delivers carbon skeletons and amino acids from photosynthetically active cells (sources) to rapidly growing tissues (sinks). Nutrient Uptake In vascular plants, nutrients are taken up by the root system and face as a first barrier the plasma membrane of root cells. Generally, nutrients are taken up by root hair cells which penetrate the soil via their growing tip. When nutrients are available at high concentrations they also can diffuse through the cell wall continuum (apoplast) of the root. The diffusion in the root, however, is limited by a radial layer of cells (endodermis), with cell walls largely impermeable to ions. Here, ions have to be transported across the plasma membrane and through the cytoplasm towards the root stele. H 1 pumps The uptake of ions into roots, and plant cells in general, is mediated by transporters in the plasma membrane. A large quantity of, ions is subsequently sequestered in the vacuole, the storage organelle of plant cells (Figure 1). Both the plasma membrane and vacuolar membrane are energized by proton (H 1 ) pumps. These membrane proteins translocate H 1 from the cytoplasm to the cell wall space (apoplast) or into the vacuole, by hydrolysing high-energy phosphates. Owing to the positive charge of protons, pump activity can be measured as a positive (outward) electrical current across membranes (Figure 2a). Proton pumps create both a concentration difference for H 1 and an electrical potential difference (membrane potential) that is negative at the cytoplasmic side. Membrane potential changes towards more positive values are referred to as depolarizations, negative going changes ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net 1 Plant Ion Transport ATPases share an autoinhibitory domain at their Cterminus. A protein, classified as 14-3-3 binds to this autoinhibitory domain and thereby can stimulate pump activity. Within the vacuolar membrane two different H 1 pumps are present, a V-type ATPase and a pyrophosphatase. The V-type ATPase is a multisubunit H 1 pump, while the pyrophosphatase is encoded by a single gene (AVP) in A. thaliana. The latter pump hydrolyses pyrophosphate (PPi) to transport H 1 . Ion channels Figure 1 Schematic diagram of ion pumps and channels in guard cells superimposed on an A. thaliana stomate. Stomatal movement is associated with ion transport. The following ion channels and pumps have been identified in guard cells: (1) a hyperpolarization-activated calcium conductance, that carries an inward Ca2 1 current; (2) outward rectifying K 1 channel, which releases K 1 after depolarization (see Figure 2c); (3) inward K 1 channel that facilitates K 1 uptake when the membrane potential is hyperpolarized (Figure 2c); (4) the plasma membrane proton pump utilizes ATP to drive H 1 extrusion, its activity hyperpolarizes the membrane potential (Figures 2a and 2b); (5) mechanically activated ion channels with selectivity for either Ca2 1 , K 1 or Cl 2 , which probably represent tension sensors; (6) fast and slow anion channels that upon activation release anions and depolarize the membrane potential (Figures 2b and d); (7) and (8) the V-type H 1 ATPase and pyrophosphatase drive H 1 extrusion into the vacuole; (9) fast vacuolar channels (FV) are active at low cytoplasmic Ca2 1 levels and conduct K 1 ; (10) K 1 -conducting VK channels are active at an intermediate cytoplasmic Ca2 1 level; (11) at a high cytoplasmic Ca2 1 concentration the outward rectifying slow vacuolar (SV) channel is activated (Figure 4a), it conducts K 1 , Ca2 1 and anions to some extent; (12) anion fluxes across the vacuolar membrane are facilitated by a protein kinase-activated anion channel; (13) Ca2 1 -permeable channels activate in a voltage-dependent manner, in response to inositol triphosphate (IP3) or cyclic ADP-ribose (cADPR). as hyperpolarizations. Plasma membrane H 1 ATPases can transport H 1 against a large electrical potential difference; membrane potentials as hyperpolarized as 2 240 mV have been reported. The potential difference across the vacuolar membrane is smaller and most likely ranges from 2 20 to 2 60 mV. Most plant cells maintain a slightly alkaline cytoplasm, with a pH value close to 7.5. The apoplast and vacuole are more acidic, with pH values often ranging from 5 to 6. However, depending on the cell type, these pH differences can be more pronounced. The H 1 pump in the plasma membrane is classified as P type ATPase, a group of transporters widely distributed throughout different phyla (Palmgren and Axelsen, 1998). In Arabidopsis thaliana H 1 ATPases are encoded by at least 10 different AHA genes. Plasma membrane H 1 2 Ion channels are pores, most of which are selective for a particular cation or anion while others discriminate less between ion species. The movement of an ion through such a channel is passive: the interaction between the ion and the channel-protein does not determine its direction of flow. However, ions are electrically charged and the ion current through channels depends on the membrane potential. The negative membrane potential thus allows uptake of cations against their concentration gradient. The relation between ion gradients and the electrical potential for an individual ion species is defined by Nernst’s law; E(I) 5 RT/(zI F) ln([I]out/[I]in), where E(I) is the driving force for ion movement, R is the gas constant, T is the absolute temperature, zI is the charge of the ion, F is the Faraday constant and [I] is the concentration of the ion. According to this law, a membrane potential of 2 240 mV allows cells to maintain a 104 times higher K 1 concentration inside than outside the cell (e.g. 100 mmol L 2 1 K 1 inside root cells and 10 mmol L 2 1 outside). Although open channels can conduct ionic currents irrespective of their direction, the process of opening (gating) is often voltage dependent. Two types of voltagedependent K 1 channels are found in most plant cells, conducting either an inward or an outward current, under physiological conditions. The inward rectifying K 1 channels activate at hyperpolarized membrane potentials, in Figure 2c negative of 2 155 mV. In the same cell, outward rectifying K 1 channels activate at depolarized potentials. K 1 channels represent the best-characterized group of ion transporters, both on the functional and the molecular levels. On the basis of sequence similarities, these channels can be divided into six families (Figure 3a). K 1 -channel families can be grouped according to the number of transmembrane domains and K 1 -selective pores (Figure 3b). Initially the Arabidopsis thaliana K 1 channels AKT1 and KAT1 were cloned through complementation of yeast mutants lacking K 1 transporters. Other members belonging to the same families and AKT3 were found by screens for homologous genes. KCO1 and SKOR1 were recognized in an A. thaliana EST database while searching for sequence homology to known K 1 channels. The SKT2 and SKT3 genes of Solanum tuberosum were discovered in ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net Plant Ion Transport Figure 2 Electrical properties of ion transporters in the plasma membrane of guard cells. (a) H 1 pump currents in Vicia faba protoplasts. Pump currents decrease at more negative potentials, when the H 1 is transported against a larger electrical gradient (membrane potential, Vm). (From Lohse and Hedrich R (1992) Planta 188: 206–214.) (b) Spontaneous changes of the free-running membrane potential (Em) of an A. thaliana guard cell. The Nernst potential of K 1 was estimated at 2 80 mV for this cell. Efflux of K 1 therefore will occur when Em is at its most positive value ( 2 60 mV), while K 1 uptake occurs during the periods at which Em is most negative ( 2 180 mV). At times indicated by ., voltage clamp protocols similar to Figure 2c were applied. (From Roelfsema and Prins (1998) Planta 205: 100–112.) (c) Whole-cell K 1 currents of an A. thaliana guard cell. Time-dependent activation of outward and inward rectifying K 1 channels occurs at membrane potentials positive of 2 80 mV and negative of 2 155 mV, respectively. (d) Singlechannel currents of the fast anion channel GCAC1 from V. faba. Opening of anion channels causes stepwise increases of negative (inward) currents, equivalent to anion efflux. The current level with all channels closed is indicated by C, that of open channels by Ox, where x stands for the number of open channels. The open probability depends on the degree of membrane depolarization. (Modified after Schulz-Lessdorf (1996) PhD thesis University of Hannover.) an approach to find proteins interacting with the Cterminus of KST1, using the yeast two-hybrid system. The AKT1 gene encodes an inward K 1 channel that is expressed in roots. Plants with a disrupted AKT1 gene exhibit reduced growth at low K 1 concentrations in the root medium and in the presence of NH41 (Hirsch et al., 1998). Apparently, the channel is involved in K 1 nutrition, but plants have alternative K 1 -uptake systems that enable mutated plants to grow normally in a K 1 -rich medium. As a consequence of the Nernst law, K 1 channels cannot provide an uptake system that operates at K 1 concentrations much lower than 10 mmol L 2 1. Below this concentration, threshold active transport systems (carriers) must secure K 1 uptake. Carriers Owing to the broad specificity of carriers, plant cells are able to take up a wide range of ions. Some carriers utilize the gradient of one ion to energize the uptake of another. In the plant plasma and vacuolar membranes, most carriers mediate proton-coupled transport. For cations such as K 1 , carriers provide a high-affinity uptake system capable of capturing K 1 from extracellular concentrations lower than 10 mmol L 2 1. For Hordeum vulgare and A. thaliana, K 1 -carriers named HAK (Santa-Maria et al., 1997) and AtKUP were found to mediate K 1 uptake, even from concentrations below 1 mmol L 2 1. The transcription of some of these transporters is increased at low K 1 concentrations in the root medium, indicating a role for these proteins in overcoming K 1 starvation. Most of the micronutrients are also taken up by carriers. Specific transporters for nutrients such as Mn, Fe, Cu, Co and Zn have been identified (Fox and Guerinot, 1998). In A. thaliana the transporters for Cu, Fe and NH41 are encoded by COPT, IRT and AtAMT genes, respectively. These plasma membrane transporters most likely mediate cotransport of these cations with H 1 . Carriers are essential for the uptake of anions against their concentration gradient, since the negative membrane potential does not support anion uptake via channels. Similar to that of K 1 , the uptake of anions like NO32 divides into components with low and high affinity. In A. thaliana, three genes encoding NO32 transporters have ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net 3 Plant Ion Transport Figure 3 Clustering of cloned plant K 1 channels based on sequence similarity. (a) Cloned K 1 channels superimposed on fluorescent branching veins of an A. thaliana leaf expressing green fluorescent protein fused to the SUC2-promoter. (Plants provided by N. Sauer, University of Regensburg; confocal laser microscopy by R. Steinmeyer.) At least six families of K 1 channels can be distinguished. The KAT1 gene codes for an inward K 1 channel from Arabidopsis thaliana, which is expressed in guard cells and flowers. KST1 is the Solanum tuberosum homologue to KAT1, while KAT2 is structurally related to KAT1. The AKT1 family also encodes inward rectifying channels. AKT1 was cloned from A. thaliana and is involved in K 1 uptake of roots; SKT1 is a related clone from S. tuberosum and ZMK1 originates from maize and is associated with auxin-induced growth of coleoptiles. Members of the AKT3 family are preferentially expressed in vascular tissue and encode K 1 channels that are largely voltage independent. AKT3 was cloned from A. thaliana, ZMK2 from maize, SKT2 from S. tuberosum, VFK1 from Vicia faba and SPICK1 and SPICK2 from Samanea saman. SKOR1 encodes an outward K 1 rectifying channel from A. thaliana, which is expressed in xylem parenchyma cells. SPORK1 is a related S. saman gene. The group of KCO1-related genes encode Ca2 1 dependent outward channels of unknown function. KCO1 and KCO2 originate from A. thaliana and SPOCK1 from S. saman. The KCO3 channel also originates from A. thaliana but differs from KCO1 and KCO2 in the number of transmembrane domains. K 1 channel genes from Hordeum vulgare (HVKCH1) and Plantago media (PMKCH1 and 2) are not grouped into one of these families. (b) K 1 channels consist of a multiplication of two transmembrane domains, indicating a common evolutionary base. Channels with two or six transmembrane domains have only one pore region, while two pore regions are found in channels with four and eight transmembrane domains. Channels with eight membrane-spanning domains have so far only been identified in animals (TWIG) and yeast (YKC1). been identified. The NLT1 gene encodes a low-affinity transporter and is constitutively expressed. The CHL1 transporter may be active in the low-affinity as well as in the high-affinity range, while NRT2 encodes a high-affinity transporter. The expression of CHL1 and NRT2 is induced when roots are exposed to NO32 . In contrast, the transcription of sulfate and phosphate transporters is often enhanced in the absence of the substrate. In A. thaliana, transporters for phosphate and sulfate are encoded by the APT and AST genes. Calcium transport and signalling Owing to the importance of phosphate in cellular processes, cytoplasmic Ca2 1 has to be maintained at micromolar levels, otherwise calcium phosphate would 4 form an insoluble precipitate. Extracellular signals can trigger transient and sustained changes in the cytoplasmic Ca2 1 concentration that serve as an intracellular signal. These signals include chemical, osmotic and mechanical stimuli and are involved in Ca2 1 -dependent responses that control nutrient uptake, movement, growth, differentiation, symbiosis and defence against pathogens (Trewavas, 1999). Calcium ion is extruded from the cytoplasm to the apoplast, the endoplasmic reticulum and the vacuole; in these compartments the Ca2 1 concentration is in the millimolar range. Ca2 1 extrusion is accomplished by Ca2 1 ATPases and possibly by the CAX; encoded Ca2 1 / H 1 antiporter present in the vacuolar membrane, identified in A. thaliana. The Ca2 1 ATPases are either calmodulin-insensitive or calmodulin-stimulated. The calmodulin-insensitive pump is found in the vacuolar ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net Plant Ion Transport membrane only, while calmodulin-stimulated Ca2 1 pumps are present both in the plasma membrane and vacuolar membrane. The latter pump type comprises ACA-encoded Ca2 1 ATPases that have an autoinhibitory domain at their N-terminus that binds calmodulin (Harper et al., 1998). After binding calmodulin, the domain is released and the pump is activated. Ca2 1 stimulates calmodulin binding and thereby regulates the activity of this type of pump. In the search for channels involved in Ca2 1 signalling, Ca2 1 -permeable channels have been described in vacuolar membrane, endoplasmic reticulum and plasma membrane. These channels release Ca2 1 into the cytoplasm upon activation. In the vacuolar membrane, Ca2 1 conductances were reported that activate in response to inositol triphosphate (IP3) and cyclic ADP-ribose (cADPR) (Allen et al., 1995) coexisting with voltage-dependent Ca2 1 channels. The endoplasmatic reticulum, another Ca2 1 store, harbours a voltage and Ca2 1 gradient-sensitive Ca2 1 channel. The plasma membrane contains both voltagedependent (Grabov and Blatt, 1998) and mechanosensitive Ca2 1 channels. Sodium toxicity The selectivity of transporters in the plasma membrane is limited and uptake of toxic ions may occur. This is the case for the micronutrient Na 1 , which is present at large concentrations in the marine environment but at only moderate quantities in most soils. Land plants benefit from low concentrations of Na 1 , but higher concentrations inhibit the growth of many crops. In irrigated soils the Na 1 content often reaches toxic levels, which complicates agriculture in these areas. Plants have developed different strategies to overcome Na 1 toxicity. Plants sequester Na 1 into their vacuole through a Na 1 /H 1 antiporter in the vacuolar membrane, which in A. thaliana is encoded by AtNHX. Alternatively, plants extrude Na 1 , either from their roots or by specialized salt glands from their leaves. The HKT1 transporter of A. thaliana couples Na 1 and K 1 influx, when expressed in nonplant systems. Based on these results, HKT1 was hypothesized to present a mechanism to overcome Na 1 toxicity (Rubio et al., 1995), although this role has been challenged. Salt tolerance in A. thaliana, however, has been shown to depend on a calcineurin-like protein phosphatase. In the presence of this protein phosphatase, plants were able to accumulate K 1 selectively from media high in Na 1 , in a Ca2 1 -dependent manner (Liu and Zhu, 1998). Carbon is Absorbed from the Atmosphere To prevent excessive loss of water, plants developed a waxy layer, the cuticula, covering the shoot. This polymer is impermeable to both H2O and CO2 and therefore also limits CO2 absorption. To control water loss on one side and enable CO2 assimilation on the other, microscopically small pores in the leaf epidermis evolved, the stomates. Stomates of C3 and C4 plants open in the light when CO2 is required for photosynthesis, but close when CO2 concentrations in the leaf increase. The plant hormone abscisic acid (ABA) mediates the response of stomates to a limited water supply. During drought, the concentration of this hormone increases, forcing stomates to close and reducing transpiration. The hormone indole-3-acetic-acid (IAA) has the opposite effect and stimulates stomatal opening, when IAA-induced growth requires increased CO2-fixation. Stomatal movement is based on ion transport Opening and closure of stomates results from volume changes of the two guard cells that surround the stomatal pore (Figure 1). These guard cells swell or shrink in response to changes in their osmotic content. Modulation of the ionic content in the two motor cells changes the extent to which they are pushed apart and thereby adjusts the stomatal aperture. The changes in osmotic content are largely due to uptake and release of K 1 salts. During stomatal opening, guard cells take up K 1 and Cl 2 and convert starch into the organic anion malate. Guard cells utilize ion channels both for the uptake and for the release of K 1 ions. This is enabled by changes in the membrane potential (Figure 2b). At depolarized membrane potentials, K 1 is extruded via outward channels, while K 1 is taken up via inward channels at hyperpolarized membrane potentials (Figure 2c). The hyperpolarized membrane potentials result from H 1 ATPase activity, while anion channel activity depolarizes the membrane potential. Two types of anion channels are present in the plasma membrane that differ in activation velocity and are referred as rapid (R-type) and slow (S-type) anion channels. Both channels activate upon depolarization; the threshold for the R-type channel is around 2 120 mV (Figure 2d). Anion channels in the plasma membrane of guard cells as well as in other plant and animal cells conduct Cl 2 and NO32 as well as small organic acids. During stomatal closure, these channels facilitate Cl 2 and malate efflux from guard cells (Keller et al., 1989). Changes in the activity of H 1 ATPases and anion channels can alter the direction of the ion fluxes across the plasma membrane, owing to their effects on the membrane potential. Light, CO2 and hormones affect the activity of these two transporters and the activity of K 1 channels. ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net 5 Plant Ion Transport Both red and blue light (Assmann et al., 1985) stimulate the activity of H 1 ATPases and trigger stomatal opening. CO2 has the opposite effect and triggers stomatal closure. This CO2 response may be mediated by malate since apoplastic malate reflects the ambient CO2 concentration. Extracellular malate activates the Ca2 1 -dependent R-type anion channels and triggers stomatal closure. The CO2 response also involves an increase in cytoplasmic Ca2 1 , or triggers Ca2 1 oscillations, in guard cells of Commelina communis (Webb et al., 1996). Comparable increases in cytoplasmic Ca2 1 were evoked by ABA and IAA, although these hormones have opposite effects on stomatal movement. The precise role of the increase in cytoplasmic Ca2 1 in regulating stomatal movement is therefore still under debate. In addition to voltage-dependent channels, mechanically activated ion channels are present in the guard cell plasma membrane that provide sensors for the membrane tension. Three stretch-activated channels have been identified that are selective for either K 1 and Cl 2 or Ca2 1 (Cosgrove and Hedrich, 1991). A number of ion channels in guard cells are regulated through changes in the cytoplasmic Ca2 1 concentration. In the plasma membrane of guard cells, both the R-type and S-type anion channels are Ca2 1 stimulated. In the same cell type, the conductance of the inward K 1 channel is reduced by cytoplasmic Ca2 1 . In the vacuolar membrane, three types of Ca2 1 regulated K 1 -permeable channels have been identified (Figure 1). The FV channels are inhibited by cytoplasmic Ca2 1 , VK channels are active at intermediate Ca2 1 levels and the SV-type channel is Ca2 1 -stimulated (Hedrich and Neher, 1987; Figure 4a). Most of these channels are also sensitive to changes in the cytoplasmic pH. In the plasma membrane of guard cells, cytoplasmic acidification stimulates the inward K 1 channel and inhibits the outward K 1 channel. The inward K 1 channel also is affected by changes in the extracellular pH; acidification stimulates this channel (Figure 4b) through a shift in the voltage dependence. The pH sensor of the KST1 channel is based on two histidine residues located on the extracellular face of the channel (Hoth et al., 1997). A. thaliana mutant analysis has revealed that guard cell responses to ABA involve protein farnesylation and phosphorylation. In guard cells, ABA affects K 1 channels, stimulates S-type anion channels and inhibits H 1 extrusion. These ABA responses were absent in guard cells expressing mutated ABI1 and ABI2 genes, while guard cells lacking a functional farnesyltransferase are ABA hypersensitive (Pei et al., 1998). Long-distance Ion Transport Nutrients taken up by root cells may be transported towards the stele via plasmodesmata that interconnect the cytoplasm of adjacent cells. Finally, the nutrients are extruded into xylem vessels by xylem parenchyma cells. The ionic flow into xylem vessels is followed by an osmotically driven flow of water into the vessels, thereby creating the root pressure. As a result, a unidirectional flow of water and ions through the xylem vessels develops, that mediates long-distance nutrient transport from the root to the shoot. The flow is enhanced by the evaporation of water from stomatal pores in the leaves, that create a tension for water flow through xylem vessels. In xylem parenchyma cells the SKOR1 channel is expressed, encoding an outward K 1 channel. Mutants of A. thaliana lacking this channel are impaired in K 1 transport into the xylem vessels, which results in a lower K 1 concentration of their shoots (Gaymard et al., 1998). During drought, expression of the SKOR1 channel is reduced by ABA, resulting in a lower conductance of outward K 1 channels in xylem parenchyma cells. Sieve tubes mediate a phloem solute flow in the opposite direction, providing roots with carbohydrates and amino acids. In addition, the phloem serves growing parts of the Figure 4 Regulation of ion channels by Ca2 1 ions and protons. (a) The SV channel is regulated by cytoplasmic Ca2 1 . At 80 mV the steady-state current increases upon elevation of cytoplasmic Ca2 1 concentrations. (From Hedrich and Neher, 1987). (b) The inward K 1 channels of A. thaliana guard cells are regulated by extracellular pH. Acidification increases the current carried by the inward K 1 channel at 2 200 mV. (From Brüggemann et al. (1999) Planta 207: 370–376.) 6 ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net Plant Ion Transport shoot such as developing leaves and flowers. The phloem is also important for the redistribution of K 1 . The xylem transports large quantities of K 1 to the leaves that would be concentrated owing to the evaporation of water if the phloem did not transport ions back to the roots. The mechanism sustaining the phloem ion transport is unknown, but VFK1 encoding a K 1 channel with homology to AKT3 is preferentially expressed in sink tissues of the shoot. The AKT3 channel is expressed in phloem companion cells and encodes a weakly voltage-dependent channel that is blocked by extracellular H 1 (Marten et al., 1999). Growth and Tropisms The marine environment, in which life developed, had a high salinity and allowed cells to maintain a large concentration of osmolytes within their cytoplasm. Later, plants encountered more dilute environments and took advantage of a ridged cell wall to prevent uncontrolled osmotic swelling. The pressure by which the plasma membrane pushes against the cell wall is termed turgor. Nonwoody plants gain their mechanical stability from this interaction. Furthermore, the turgor drives the expansion of growing cells. During growth, cell walls become extensible and cells swell through the uptake of ions and water, followed by de novo synthesis of membranes and cell wall. The cell wall extensibility is regulated by expansins, acid-stimulated proteins. Stimuli like light and IAA can induce acidification of the cell wall and trigger cell growth. Acid growth of cells is accompanied by activation of H 1 ATPases and several ion channels. The growth response of maize coleoptiles, triggered by IAA, involves transcriptional regulation of the K 1 channel ZMK1. Expression of ZMK1, a K 1 -uptake channel gene homologous to AKT1 (Figure 3), correlates with coleoptile elongation (Philippar et al., 1999). Upon gravistimulation, auxin redistributes between the upper and lower half of the coleoptile. An increase of ZMK1 expression in the lower half and a decrease in the upper precedes the upward bending of this organ. Apparently, the auxin response involves an increase of K 1 channel density. Hormonal control of ion channel transcription was also described for SKOR1 (see above). A growth pattern different from cell elongation is found in Fucus rhizoids, root-hair and pollen-tube cells, which display tip-growth. In Fucus rhizoids, mechanically activated channels can increase Ca2 1 concentrations in the rhizoid tip. Growth in all three cell types is reflected by a Ca2 1 gradient from the growing tip towards the base of the cell. Changes in growth direction can be evoked by mechanical and chemical stimuli and are accompanied by a change in the Ca2 1 gradient. These Ca2 1 gradients enable root hairs to grow away from the root and around objects in their way, while pollen tubes are guided towards the oocytes by a largely unsolved chemical messenger mechanism. Living in a Community Within their natural habitats, plants interact with many different organisms. Often these interactions are disadvantageous to the plant, as they are affected by animals, fungi or bacteria. Plants have mechanisms to defend themselves against pathogens. Some pathogens are recognized by plant cells through messengers, called elicitors, extruded by the pathogen. These elicitors were found to activate Ca2 1 channels in the plasma membrane of tomato and parsley cells (Zimmermann et al., 1997). Subsequently, the elicitorevoked rise in cytoplasmic Ca2 1 mediates a defence response to the pathogen. Some interactions with microorganisms, however, are beneficial and stimulate plant growth under certain conditions. Most common are symbiotic interactions with soil fungi, called mycorrhiza. Mycorrhiza penetrate the soil over large distances and provide the plant root with nutrients that are scarce in the soil. The fungi benefits from carbohydrates released by the roots. Plants have developed mechanisms to recognize nitrogen-fixing bacteria that enable them to grow on soils poor in fixed nitrogen. Their roots associate with bacteria such as Azoarcus or Azospirrilum that produce NH41 using the N2 that is readily available. In tomato, the LeAMT2 transporter is induced by NH41 and nitrogen-fixing bacteria, providing an uptake mechanism for NH41 . A more intense interaction occurs between Rhizobia and roots of certain plant species. Here, the plant root forms a structure, called nodule, to host the bacteria within the root. The Rhizobia bacteria are surrounded by the peribacteroid membrane in which specialized transport proteins are present. In this membrane an NH41 -permeable channel was identified (Tyerman et al., 1995) that transports the NH41 produced by the bacteria towards the sites of amino acid synthesis in the root. Endosymbiosis represents a tight interaction between microorganisms and cells. Chloroplasts most likely originate from an endosymbiotic interaction. Following the invasion of plant cells by cyanobacteria, this green symbiont evolved into the photosynthetic organelle. This hypothesis is supported by the finding that protein transport proteins and K 1 channels of chloroplasts are closely related to that of the cyanobacteria Synechocystis. Ion Transport in Progress The molecular structure of Ca2 1 -permeable channels expressed in the plasma membrane and vacuolar mem- ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net 7 Plant Ion Transport branes remains as yet unknown. These channels will probably be identified either by homology to an L-type like channel of yeast or by functional complementation of the respective yeast mutant. Chloride channels have been identified in an approach as outlined above, but these channels so far lack functional expression in heterologous systems. So far, most of the ion channels cloned have been voltage-dependent, with the exception of the AKT3 channel family, which is blocked by protons but largely voltage-insensitive. Genes encoding channels activated by ligands, which are commonly found in animal cells, have so far not been isolated in plants. Recently, however, sequences with similarity to the channels binding glutamate and cyclic nucleotides have been found for plants. These newly identified genes offer intriguing new areas for research in ion transport and signalling processes of plants. Their animal counterparts have functions in the brain or sensory cells of the retina; tracking the function of these genes in plants may therefore answer questions about how plant cells communicate. References Allen GJ, Muir SR and Sanders D (1995) Release of Ca2 1 from individual plant vacuoles by both insP3 and cyclic ADP-ribose. Science 268: 735–737. Assmann SM, Simoncini L and Schroeder JI (1985) Blue light activates electrogenic ion pumping in guard cell protoplasts of Vicia faba. Nature 318: 285–287. Brüggemann L, Dietrich P, Dreyer I and Hedrich R (1999) Pronounced differences between the native K+ channels KAT1 and KST1 asubunit hormones of guard cells. Planta 207: 370–376. Cosgrove DJ and Hedrich R (1991) Stretch-activated chloride, potassium and calcium channels coexisting in plasma membranes of guard cells of Vicia faba L. Planta 186: 143–153. Fox TC and Guerinot ML (1998) Molecular biology of cation transport in plants. Annual Review of Plant Physiology and Plant Molecular Biology 49: 669–696. Gaymard F, Pilot G, Lacombe B et al. (1998) Identification and disruption of a plant shaker-like channel involved in K 1 release into the xylem sap. Cell 94: 647–655. Grabov A and Blatt MR (1998) Membrane voltage initiates Ca2 1 waves and potentiates Ca2 1 increases with abscisic acid in stomatal guard cells. Proceedings of the National Academy of Sciences of the USA 95: 4778–4783. Harper JF, Hong B, Hwang I et al. (1998) A novel calmodulin-regulated Ca2 1 -ATPase (ACA2) from Arabidopsis with an N-terminal autoinhibitory domain. Journal of Biological Chemistry 273: 1099–1106. 8 Hedrich R and Neher E (1987) Cytoplasmic calcium regulates voltagedependent ion channels in plant vacuoles. Nature 329: 833–835. Hirsch RE, Lewis BD, Spalding EP and Sussman MR (1998) A role for the AKT1 potassium channel in plant nutrition. Science 280: 918–921. Hoth S, Dreyer I, Dietrich P, Becker D, Müller-Röber B and Hedrich R (1997) Molecular basis of plant-specific acid activation of K 1 -uptake channels. Proceedings of the National Academy of Sciences of the USA 94: 4806–4810. Keller B, Hedrich and Raschke K (1989) Voltage-dependent anion channels in the plasma membrane of guard cells. Nature 341: 450–453. Lohse G and Hedrich R (1992) Characterization of the plasma membrane H+-ATPase from Vicia faba guard cells. Planta 188: 206–214. Liu J and Zhu J-K (1998) A calcium sensor homolog required for plant salt tolerance. Science 280: 1943–1945. Marten I, Hoth S, Deeken R, Ache P, Ketchum KA, Hoshi T and Hedrich R (1999) AKT3, a phloem-localized K 1 channel, is blocked by protons. Proceedings of the National Academy of Sciences of the USA 96: 7581–7586. Palmgren MG and Axelsen KB (1998) Evolution of P-type ATPases. Biochimica et Biophysica Acta 1365: 37–45. Pei ZM, Ghassemian M, Kwak CM, McCourt P and Schroeder JI (1998) Role of farnesyltransferase in ABA regulation of guard cell anion channels and plant water loss. Science 282: 287–290. Philippar K, Fuchs I, Lüthen H et al. (1999) Auxin-induced K+ channel expression represents an essential step in coleoptile growth and gravitropism. Proceedings of the National Academy of Sciences of the USA, in press. Roelfsema MRG and Prins HBA (1998) The membrane potential of Arabidopsis thaliana guard cells; depolarizations induced by apoplastic acidification. Planta 205: 100–112. Rubio F, Gassmann W and Schroeder JI (1995) Sodium-driven potassium uptake by the plant potassium transporter HKT1 and mutations conferring salt tolerance. Science 270: 1660–1663. Santa-Maria GE, Rubio F, Dubcovsky J and Rodrı́guez-Navarro A (1997) The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter. The Plant Cell 9: 2281– 2289. Tyerman SD, Whitehead LF and Day DA (1995) A channel-like transporter for NH41 on the symbiotic interface of N2-fixing plants. Nature 378: 629–632. Trewavas AJ (1999) Le calcium, c’est la vie: Calcium makes waves. Plant Physiology 120: 1–6. Webb AAR, McAinsh MR, Mansfield TA and Hetherington AM (1996) Carbon dioxide induces increases in guard cell cytosolic free calcium. The Plant Journal 9: 297–304. Zimmermann S, Nürnberger T, Frachisse J-M et al. (1997) Receptormediated activation of a plant Ca2 1 -permeable ion channel involved in pathogen defense. Proceedings of the National Academy of Sciences of the USA 94: 2751–2755. ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net Plant Ion Transport ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net 9