1 ANTH 310 Human Origins LABORATORY #3 EARLY HOMININS
advertisement

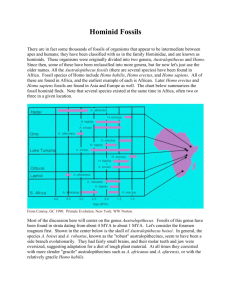
ANTH 310 Human Origins LABORATORY #3 EARLY HOMININS: AUSTRALOPITHECINES AND EARLY HOMO In Preparation for this lab, review the lecture notes, appropriate chapter in the Conroy text, and pages 282-324 I the Photographic Atlas. This lab focuses on some of the earliest hominin fossils. The main objectives will be to observe cranial, dental, and skeletal morphological features in these specimens that will provide background for assessing the evolutionary history of the human family. The lab format consists of a number of lab demonstrations specifically designed to illustrate some of the important cranial, dental, and postcranial variation present in these fossils. Early Hominins The earliest specimens, which are considered hominins primarily because of their obvious adaptations to bipedal locomotion, and because they share dental features (e.g., reduction in the size of anterior teeth) with later hominins, split from the apes approximately 5-10 mya. Several different genera and many new species of these early hominins are now known. This lab will concentrate on some of the most primitive australopithecine species (A. afarensis) as well as later, robust and gracile varieties and the earliest members of the genus, Homo. Leaving aside many of the recently discovered fossils (e.g., Ardipithecus ramidus, Australopithencs anamensis, Australopithencs garhi, A. bahrelghazali, Orrorin tugenensis, Sahelanthropus tchadensis,and Kenyanthropus platyops), which predate earlier discovered hominins, the majority of these fossils are classified in the genus, Australopithecus, and are found at sites located in eastern and southern Africa. Traditionally, this genus has been divided into a robust and a gracile form. Each is associated with savannah habitats like those found in Africa, today. The australopithecines disappear from the fossil record shortly after 1 million years ago. Australopithecus afarensis Beginning in the 1970s, a good number of specimens of A. afarensis were discovered at sites such as Hadar, Ethiopia, and Laetoli, Tanzania. Radiometric dates for these sites range roughly between 3 and 4 mya. A. afarensis exhibits a number of primitive as well as derived characters. The most striking derived character are the adaptations of the pelvis and foot for habitual bipedal locomotion. On the other hand, A. afarensis retained longer, more curved metatarsals and foot phalanges. Others, who have undertaken detailed analyses of the foot remains, have argued that A. afarensis was clearly bipedal and did not use the trees to a great extent (although some have argued that A. afarensis was more adapted to walking rather than running while others believe that A. afarensis was still partially adapted to climbing in trees). Others have argued that the long curved 1 phalanges are a retention from a common ancestor with the apes. Their bones show that they were physically very strong. Females were substantially smaller than males, a condition known as sexual dimorphism. Height varied between about 107 cm (3'6") and 152 cm (5'0"). Body weight estimates for A. afarensis indicate they weighed between 30 kg and 80 kg. A. afarensis had an apelike face with a low forehead, a bony ridge over the eyes, a flat nose, and no chin. They had protruding jaws with large back teeth. Cranial capacity varied from about 375 to 550 cc. The skull is similar to that of a chimpanzee, except for the more humanlike teeth. The teeth of A. afarensis and all the australopithecines were megadont; i.e., their cheek teeth were large relative to body size. The canine teeth of A. afarensis are much smaller than those of modern apes, but larger and more pointed than those of humans, and shape of the jaw is between the rectangular shape of apes and the parabolic shape of humans. Some have argued that the specimens from Laetoli and Hadar represent two species while others maintain that A. afarensis was a highly sexually dimorphic species. South African Australopithecines The first australopithecine was discovered by Raymond Dart in South Africa, the Taung child. Since then, many other specimens have been recovered from South African sites. Traditionally, the specimens have been placed into two species, A. africanus and A. robustus, although the latter has been placed in a separate genus (Paranthropus) by some. Australopithecus africanus specimens are found at fossil sites (e.g., Taung, Sterkfontein) dated from about 2.3 to 3 million years ago while the A. robustus specimens (e.g., Swartkrans) are dated from about 1.5 to 2 mya. Because the cranial and postcranial remains of A. africanus specimens are a bit more delicate than the rugged robust remains of A. robustus, A. africanus is usually referred to as a ‘gracile’ species while A. robustus is generally referred to as a Arobust’ species although both had average body weights of about 50 kg. Australopithecus africanus is similar to A. afarensis, and was also bipedal, but body size was slightly greater. Brain size may also have been slightly larger, ranging between 420 and 500 cc. This is a little larger than chimp brains (despite a similar body size), but still not advanced in the areas necessary for speech. The back teeth were a little bigger than in A. afarensis. Although the teeth and jaws of A. africanus are much larger than those of humans, they are far more similar to human teeth than to those of apes. The shape of the jaw is now fully parabolic, like that of humans, and the size of the canine teeth is further reduced compared to A. afarensis. Australopithecus robustus had a body similar to that of A. africanus, but a larger and more robust skull and teeth. The massive face is flat or dished, with no forehead and large brow ridges. It has relatively small front teeth, but massive grinding teeth in a large lower jaw. Most specimens have sagittal crests. Its diet would have been mostly coarse, tough food that needed a lot of chewing. The average brain size is about 530 cc. East African Australopithecines 2 There have been no undoubted specimens of A. africanus found in East Africa. However, hyper-robust species, A. boisei, lived in East Africa between 2.5 mya to just about 1 mya. A. boisei has been found at several sites in East Africa.. Australopithecus boisei (formerly Zinjanthropus boisei) is similar to A. robustus of South Africa, but the face and cheek teeth were even more massive, some molars being up to 2 cm across. The brain size is very similar to A. robustus, about 530 cc. The A. robustus/boisei dentition represents the culmination of dental trends first observed in the Miocene ramamorphs: a reduction of the anterior dentition, enlargement of the cheek teeth with thick enamel, and thickening of the jaws. The premolars of A. boisei/robustus are referred to as molariform premolars because of the molar-like form and function of these teeth. The large, thick-enameled cheek teeth probably provided the surface area for grinding small, tough food which need little slicing and cutting by the anterior teeth, which are small. Recently, a robust australopithecine was found in Kenya with a very large face similar to A. boisei. This specimen, referred to as the ‘black skull’, has been dated to 2.6 mya. It may be an ancestor of A. robustus/boisei, but it has a baffling mixture of primitive and advanced traits. The brain size is very small, at 410 cc, and parts of the skull, particularly the posterior portions, are very primitive, most resembling A. afarensis. Other characteristics, like the massiveness of the face, jaws and single tooth found, and the largest sagittal crest (a sagittal crest is a bony ridge on top of the skull to which chewing muscles attach.) in any known hominid, are more reminiscent of A. boisei. Because it unusual features and early date, this specimen has been placed in a new species, A. aethiopicus. Australopithecus aethiopicus, robustus and boisei are all referred to as robust australopithecines, because their skulls in particular are more heavily built than the gracile australopithecines. Estimates of the cranial capacities of the australopithecines range from just under 400 cubic centimeters (cc.) to more than 500 cc. When body size is taken into consideration, these cranial capacities are not much more than those of the extant apes. Brain expansion, so characteristic of out lineage, began with the first appearance of our genus. Homo habilis/rudolfensis Living contemporaneously in East Africa (and South Africa) was another hominid species, Homo habilis. H. habilis, the "handy man", was so called because of evidence of tools found with its remains. H. habilis existed between 2.4 and 1.5 million years ago. It is very similar to australopithecines in many ways. The face is still primitive, but it projects less than in A. africanus. The back teeth are smaller, but still considerably larger than in modern humans. The average brain size, at 650 cc, is considerably larger than in australopithecines. Brain size varies between 500 and 800 cc, overlapping the australopithecines at the low end and H. erectus at the high end. The brain shape is also more humanlike. The bulge of Broca's 3 area, essential for speech, is visible in one habilis brain cast, and indicates it was possibly capable of rudimentary speech. H. habilis is thought to have been about 127 cm (5'0") tall, and about 45 kg (100 lb) in weight, although females may have been smaller. Recently, it has been prposed that two species of Homo coexisted 2 million years ago, Homo, Homo habilis and Homo rudolfensis. Homo habilis was a smaller-brained creature with archaic postcranial skeleton and H. rudolfensis was larger-brained with a more modem postcranium. Bipedalism Because of the changes in the ilium, the center of gravity is different in the pelvic region between quadrupeds and bipeds. The area of articulation of the sacrum and ilium (auricular surface) in humans has moved with the ilium until it is closer to the acetabulum, which bring the weight from the spinal column closer to the femur head. This is a more stable arrangement for a biped. Abductors and adductors of the femur maintain lateral balance by shifting body weight more over the sagittal line (midline) of the body. The lateral extension of the ilium and the lengthening of the femoral neck add power by increasing the lever arm for those muscle. Comparison of the pelvic (innominate) bones of various Early hominins, modern H. sapiens, and a living great apes indicates that many of the anatomical features they possess are related to bipedalism or the lack of bipedalism. The following is a partial list of some of the important morphological characteristics which are found in the innominate bone of modern hominins: 1. short broad ilium (extends attachment of gluteal muscles) 2. backward extension of iliac crest (moves gluteus maximus) 3. low position of sacral articulation relative to the acetabulum (increases stability in weight transmission) 4. orientation of sacral articulation relative to whole hip bone 5. sharp angle of sciatic notch produced by bending of ilium onto ischium 6. prominent ischial spine (for ligaments supporting weight) 7. strongly developed anterior inf. iliac spine (for rectus femoris and ilio psoas, hip flexors) 8. shortened length of the ischium 9. well-marked groove for ilio psoas muscle (this muscle acts to flex and rotate the hip and supports the hip in extension) 10. orientation and position of the ischial tuberosity in relation to the acetabulum The attached diagram will help to locate some of the features mentioned. The innominate bone of a pongid (e.g., chimp) will possess many features which contrast those present in hominins. 1. elongated narrow ilium 2. limited iliac crest 3. high position of sacral articulation away from acetabulum 4. less angulated sacrum relative to rest of hip 5. shallow sciatic notch 6. less prominent ischial spine and ant. inf. iliac spine 7. longer ischium (for separate attachment of gluteus maximus) etc. 4 Exercise No. 1 Cranial morphology of Early hominins. Using the specimens provided, compare the following cranial features. Feature A. afarensis A. africanus Cranial size/shape Face (dimensions, prognathism, etc.) Muscle attachments (sagittal, nuchal crests etc.) Forehead Brow ridge development/ supraorbital tori Mandible (size, chin, etc.) 5 A. robustus Early Homo Exercise No. 2 Dental morphology of Early hominins. Using the specimens provided, compare the following dental features. Feature A. afarensis A. africanus Tooth size Relative size of the posterior teeth (premolars and molars) to anterior teeth Canine size/shape Diastema (location, size) Lower first premolar (P3) Tooth row shape Molar (appearance and cusp patterns) Other 6 A. robustus Early Homo Ape Exercise 3. Postcranial Comparisons. Using specimens of the pelvis, femur, and foot bones of a modern ape, australopithecine, and modern human, make detailed comparisons of the following traits. Feature/trait Ape Shape of ilium & iliac crest Greater sciatic notch Sacro-iliac articulation (extent & placement reative to acetabulum) Acetabulum (size & shape) Ischial spine Ant. inf. iliac spine Ischium length Femur Foot bones 7 Australopithecine Modern Human Exercise 4. Based on your observation in Exercise 3, were the Austalopithecines capable of facultative or obligatory bipedalism. Explain. Exercise 5. List some of the major dental and cranial differences between Australopithecus africanus and Australopithecus robustus. Australopithecus robustus Australopithecus africanus Exercise 6. Make a list of salient differences between the crania of A. robustus with A boisei Australopithecus robustus Australopithecus boisei Exercise 7. What differences are there between KNMER 1470 and KNMER 1813 crania? Do the differences warrant a separate (or same) species designation? Explain. 8 Comparative Anatomy of the Pelvis Typical Quadruped (e.g., horse) Typical Biped (e.g., human) Typical Brachiator (e.g., gibbon) Ilium Long & narrow, greater sciatic notch is very shallow, forming an obtuse angle Short and splayed, greater sciatic notch is deeper, less obtuse Intermediate Gluteus muscles Gluteus maximus, medius, and minimus are all abductors (draw leg away from midline of the body) Gluteus maximus is an extensor (see Fig. below) 9 10 11 Early Hominids and Their Immediate Ancestors SPECIES pre-hominid apes or ape-hominid transitional species Sahelanthropus tchadensis Orrorin tugenensis 7-6? 6.0 Ardipithecus ramidus 5.8 - 4.4 Australopithecus anamensis 4.2 - 3.9 Australopithecus afarensis 4.0 - 2.9 gracile Kenyanthropus platyops species Australopithecus bahrelghazalia early hominids TIME RANGE (millions of years ago) Australopithecus africanus Australopithecus garhi Australopithecus aethiopicus robust species Australopithecus robustus Australopithecus boisei 3.5 - 3.2 3.4 - 3.0 3.3 - 2.3 2.5 2.5 2.0 ? - 1.5 2.0 - 1.2 The species listed above in red are controversial because of relatively limited discoveries of their fossils and disagreement as to their classification. Those listed in black have been found at many sites and now are widely accepted. Some researchers refer to the robust australopithecines as paranthropoids (genus Paranthropus). 12