Intership - Generation time in matrix population models (2013)
advertisement
")
École Normale Supérieure
Bachelor Internship
Laboratory Ecology and Evolution (UMR 7625)
Eco-Evolutionary Mathematics unit
Generation Time in
Matrix Population Models
Supervisor:
Intern:
François Bienvenu
Licence de biologie
École Normale Supérieure
Stéphane Legendre
Équipe ‘Éco-Évolution mathématique’
École Normale Supérieure (UMR 7625)
legendre@ens.fr
francois.bienvenu@ens.fr
3 June - 26 July 2013
Contents
Presentation of the laboratory
The Ecology and Evolution laboratory (UMR 7625) . . . . . . . . . . . . . . . . . . . . . .
The Eco-Evolutionary Mathematics unit . . . . . . . . . . . . . . . . . . . . . . . . . . . .
0
0
0
Thanks
0
Introduction
1
Methods
Graph theory . . . . . . . . . . . . . . . . . .
Vocabulary . . . . . . . . . . . . . . . .
Adjacency matrices . . . . . . . . . . . .
Primitivity . . . . . . . . . . . . . . . .
The Perron-Frobenius theorem . . . . .
Matrix population models . . . . . . . . . . .
The model . . . . . . . . . . . . . . . . .
Eigen-elements . . . . . . . . . . . . . .
Sensitivities and elasticities . . . . . . .
Markov chains . . . . . . . . . . . . . . . . .
The model . . . . . . . . . . . . . . . . .
Stationary distribution and return time
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
1
1
1
2
2
2
2
2
3
3
3
3
4
Results
Markovization . . . . . . . . . . . . . . .
Motivation and principle . . . . . .
Interpretations . . . . . . . . . . .
Mathematical expression . . . . . .
Stationary probability distribution
Line graph . . . . . . . . . . . . . . . .
Motivation . . . . . . . . . . . . .
Construction . . . . . . . . . . . .
Properties . . . . . . . . . . . . . .
Generation time . . . . . . . . . . . . .
The A = R + S decomposition . .
e . . . . .
The transformed graph P
Distribution of T . . . . . . . . . .
Mean of T . . . . . . . . . . . . . .
Other results . . . . . . . . . . . .
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
4
4
4
5
5
5
6
6
6
7
9
9
9
10
10
10
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
Discussion
11
Existing measures of the generation time . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
Perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
Conclusion
13
References
14
Appendix
A.1 The age-classified model .
A.2 The standard size-classified
A.3 More on the line graph . .
Notations (cheat-sheet) . . . .
. . . .
model
. . . .
. . . .
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
I
.
I
. II
. IV
. VI
Presentation of the laboratory
The Ecology and Evolution laboratory (UMR 7625)
The aim of the UMR 7625 is to bring together the different levels of study of ecological processes,
which range from the study of the gene to that of the ecosystem. Most of the domains of modern
ecology are represented in the laboratory, with a strong emphasis on evolutionary ecology. The
questions addressed are of theoretical importance, but they are tackled with real-world problems
in mind (global change, conservation, emerging infectious diseases) and the UMR 7625 aims at
developing interdisciplinary work with the relevant fields. Given the variety of the topics, the
laboratory is organized in five research units :
• Evolution of Animal Societies
• Global Change and Adaptive Processes
• Eco-Evolutionary Mathematics
• Evolutionary Physiology
• Populations and Communities Ecology
The Eco-Evolutionary Mathematics unit
The Eco-Evolutionary Mathematics unit develops mathematical tools to better understand ecoevolution. It is composed of mathematicians, physicists and biologists who work together to link
theory and practice. Although not apparent in this report – which focuses on a deterministic, linear
model – a strong focus is put on stochastic and non-linear models, and the interaction between both.
Research subjects include speciation, small population dynamics, bio-ecological oscillations, trophic
networks, epidemiology, genome dynamics and eco-evolutionary dynamics in turbulent fluids. The
question typically encountered are: ‘How did this pattern evolve ?’, ‘What does it tell us about the
current dynamic of the system ?’, ‘Can we use this information to better manage it ?’
In this context, the work of Stéphane Legendre (who incepted and supervised this work) has
focused on population dynamics (small populations), conservation biology, evolutionary dynamics,
trophic networks, the metabolic theory of ecology and the link between these topics. His scientific
activity also consists in writing software aimed at ecologists.
Thanks
First of all I would like to thank Stéphane Legendre, who gladly presented me his work before
offering me an internship; who made sure that I had a desk and proper working conditions; who
accepted to take quite some time to talk about the subject of the intership, even when we had very
different opinions; and finally who made a careful correction of this manuscript.
I would also like to thank Renaud De Rosa, whose Idées et Théories en Biologie lesson gave me
the occasion to re-read the work of Richard Dawkins and who answered a few questions I asked
him (alas, forcing me to face reality).
Finally, I would like to thank all my friends and everyone with whom I even remotedly discussed
this work. I also thank BIOEMCO for the warm and intellectually stimulating atmosphere it
provided during my intership, and I deplore the fact that my successors won’t be able to benefit from
it as the whole laboratory was recently told to leave the École Normale Supérieure on appalingly
short notice.
0
found on arXiv [6]. This version, written mainly by Stéphane
Legendre, is aimed at making the results look as natural as
possible for people working with matrix population models,
whereas the aim of the present report is to be as general and
rigorous as possible when describing the methods used, since
they can be generalized to other models described by weighted
directed graphs.
Introduction
When they want to convert generations into years, molecular
biologists as well as historians use a quantity called the generation time. Intuitively, this is the difference in age between
the generation of an individual and that of his parents (about
20 to 30 years in human populations). Its many applications
and the resulting need for solid estimates justifies theoretical
studies of the generation time, but there are other important motivations: indeed, the generation time is a fundamental descriptor of organisms and populations. For instance,
it has allometric scaling with other biological descriptors [1]
(a mathematical formulation of the fact that smaller organisms tend to have shorter generation times). This makes it
a challenge for the metabolic theory of ecology, which aims
at explaining such relations from the metabolic rate of organisms. The generation time is also the most natural candidate
for a time-scale when describing processes at the level of the
population. Finally, it is arguably a valuable proxy for the
timing of evolution.
Surprisingly for such a basic concept, there is no single definition of the generation time – and the way in which the existing definitions relate is poorly understood. Three definitions
have been commonly used [2], and mathematical expressions
for each of them have been derived for several demographic
models, notably matrix population models [3, 4]. However,
these formula are rather complicated and hard to interpret.
Here, we provide a surprisingly simple and general formula
which we show to encompass the classic and intuitive notion
of the generation time as difference in age between an individual and its parents.
We focused on matrix population models because they are
among the most simple population models and yet provide a
great variety of information. Moreover, they are widely used
in conservation biology. Rather than trying to simplify existing formulas, we started from a new definition which takes
advantage of the cyclic nature of the life cycle to define the
generation time as the time between two reproductive events
in a realization of the life cycle. To perform the mathematical
calculation, we used Markov chains and graph theory. Markov
chains were used to model the life cycle in a way which makes
apparent its cyclic nature – we call this process of building
a Markov matrix out of the classic representation of the life
cycle as a population projection matrix markovization and
it was first introduced by Demetrius [5]. Graph theory was
used to avoid the ambiguous definition of newborn stages by
focusing on reproductive events rather than individuals. This
technique greatly eases the calculation and has to our knowledge never been used in matrix population models.
We also illustrate the interest of our formula by generalizing a result know as Lebreton’s formula and by reflecting on
the link between the generation time and another important
biological descriptor known as population entropy. However,
this topic calls for more work.
Much of the reasoning of this work was made possible by
thinking in terms of graphs rather than matrices. Because
of this, this document was written using the formalism of
graphs. A more classic, ‘matrix’, version of this work can be
Methods
In this section we present the mathematical tools used. Although we mostly rely on elementary linear algebra (the most
elaborate result we use is the Perron-Frobenius theorem),
graph theory, matrix population models, and Markov chains
each have their own vocabulary and conventions, which can
make even the simplest reasoning hard to follow for someone
not acquainted with them.
The following notations will hold for the rest of the document: we denote scalars with lowercase letters, such as p.
Matrices are in uppercase bold letters (e.g. A) and vectors –
row and column – in lowercase bold letters (e.g. v). Finally,
it is implicit that A = (aij ) and v = (vj ). A cheat-sheet
aimed at easing the reading of this document can be found in
the appendix.
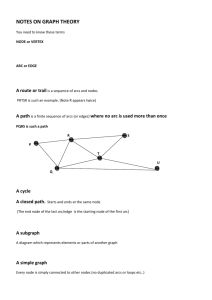
Graph theory
Here we do not aim at providing an introduction to graph
theory but only at defining the vocabulary we use and giving
the main results we will need.
Vocabulary
A graph is a set of vertices, or nodes, linked by edges, or
arcs. Graphs can be directed or not, depending on whether
their edges denote a symetric relation between the vertices
they link (non-directed) or not (directed). Directed graphs
are sometimes called digraphs, but since we work exclusively
with directed graphs we will simply refer to them as ‘graphs’.
To follow what is usually done when working with directed
graphs, we will prefer the terms nodes and arcs over the terms
vertices and edges.
The node from which an arc starts is called its tail, and the
node to which it leads its head. We note t and h the functions
which map an arc to its tail and head, respectively. The arcs
coming to a node will be refered to as its in-arcs and those
leaving it as its out-arcs.
We work with weighted graphs, which means that each arc
has a weight, a scalar representing the intensity of the relation
between the head and the tail of the arc. We note WG the
weighting function of graph G, i.e. the function which map
an arc to its weight.
A node i leads to a node j if and only if there is an arc
going from i to j, in which case we note i → j. Similarily
we can say that an arc (i → j) leads to (k → l) if and only
if j = k. A path from i to k is an ordered sequence of arcs
{(i → j1 ), (j1 → j2 ), ..., (jn → k)}. When there exists a path
from i to k, we say that k is reached by i and we note i
k.
A path from a node i to itself is called a cycle (including the
1
case of the self-loop (i → i)). The length of a path / cycle
is simply the number of arcs composing it (therefore, a selfloop has length one). Finally, a graph is said to be strongly
connected if for any pair of nodes (i, j), i
j.
Nothing in those definitions imply that there is only one arc
going from one node to another. However, graphs are often
assumed to have this property. Those that do not are called
multigraphs. Here, we will use ‘graph’ when it does not matter
whether or not a graph is a multigraph, and precise ‘simple’
and ‘multi’ only when we want to emphasize its nature.
Let G be a primitive matrix. There exists an eigenvalue λ
of G so that :
• λ ∈ R, λ > 0
• ∀µ ∈ Sp G,
|λ| > |µ|
• λ is simple, i.e. has multiplicity one.
λ is called the dominant eigenvalue of G. Its right and left
eigenvectors are called the dominant eigenvectors of G, and
they are positive (i.e. all their entries are > 0).
As stated, this formulation of the Perron-Frobenius theorem is quite imprecise, but it is sufficient for our needs. For
a more detailed presentation of the results, see [7].
Adjacency matrices
It is possible to represent a graph with an adjacency matrix.
If G is a simple graph, its adjacency matrix is the matrix G
such that:
WG (i → j) if i → j
gij =
0 else
Matrix population models
Matrix population models were first introduced by Leslie as
age-classified models [8]. They were later generalized to other
type of classes (such as size or developmental stages) by
Lefkovitch [9]. Due to their simplicity and richness, theses
models have been extensively studied and used (especially in
conservation studies). The most complete introduction to the
subject is probably Caswell’s Matrix Population Models [10],
from which the following results were taken.
In that case, G contains all the information about G. But if G
is a multigraph, it cannot be represented unequivocally by a
single matrix. Nevertheless, we can still define its adjacency
matrix. Here, we do so by using the same definition as before
except we adapt it so that gij is the sum of the weights of
every arc going from i to j. The resulting definition coincide
with the classic one for simple graphs.
Of course, defining gij as a function of (i → j) rather than
(j → i) is an arbitrary convention – and unfortunatly this
convention is not respected in matrix population models. One
can deal with this problem either by keeping one definition
of adjacency matrices and working with transposes, or by
adapting the definition of the adjacency matrix when studying
matrix population models. We will use the latter possibility.
The model
The simplest demographic model one can imagine is arguably
the discrete-time malthusian growth model :
n(t + 1) = λ n(t)
Where n is the size of the population and λ is its growth rate.
However, this model supposes that the vital rates of all
individuals are identical, which is obviously false even when
leaving aside individual variability: indeed, in most cases the
population can be divided in stages (age-classes, size-classes,
instars...) with potentially very different fertilities and/or
survival probability. A simple way to take this into account
is to use a matrix population model :
Primitivity
Primitivity is the property we will rely on the most, as
it makes it possible to use the Perron-Frobenius theorem.
Primitivity can be broken down into three properties: nonnegativity, irreducibility and aperiodicity.
The matrix G is non-negative if and only if ∀(i, j), gij ≥ 0.
It is irreducible if and only if the graph it represents when
interpreted as an adjacency matrix is strongly connected. Finally, the period of a graph is the greatest common divisor
(GCD) of the lengths of all its cycle and a graph is said to
be aperiodic if and only if it has period one (therefore, any
graph containing a self-loop is aperiodic). As with irreducibility, the aperiodicity of a matrix is equivalent to that of the
corresponding graph.
There exist purely matricial characterizations of irreducibility (not similar by permutation to a block upper triangular
matrix), aperiodicity (gcd {m | (Gm )ii > 0} = 1) and primitivity (G ≥ 0 and ∃m, Gm > 0) but the graph definitions
are more intuitive and often easier to verify.
n(t + 1) = An(t)
Here, n is a population vector whose entries are the number
of individuals in each class. A is the projection matrix of the
population.
With this model, each class has a different contribution to
the composition of the population at time t + 1. However, all
individuals inside a class are supposed to have identical vital
rates, and there is no density dependence. It can therefore
be said that this model is a natural, simple but powerful,
extension of the malthusian growth model.
An good way to interpret this model is to see it as a discrete
time compartment model and think in term of graph. The
projection matrix is indeed the adjacency matrix (sensu lato,
see adjacency matrices) of a graph whose nodes are the ‘compartments’ containing individuals in each class, and whose
arcs denote the flows between compartments. This graph is
what is usually called the life cycle of the population. Figure 1 is an example of a life cycle and its associated projection
matrix.
The Perron-Frobenius theorem
What is known as the Perron-Frobenius theorem is actually
a set of results proved by Oskar Perron and Georg Frobenius
concerning the eigen-elements of positive and non-negative
square matrices. We will only give the results we need.
2
Sensitivities and elasticities
As the asymptotic growth rate, λ, is a valuable indicator of
the population’s welfare, it is natural to ask how reliable it
is. This is one of the motivation for sensitivity analysis. The
sensitivity of the growth rate to an entry aij of A is :
f3
f2
1
s1
g1
2
s2
g2
3
s1
g1
0
f2
s2
g2
f3
0
s3
sλ (aij ) =
s3
Of course, the sensitivity of any variable to any parameter can
be defined. If not precised, the term ‘sensitivity’ will refer to
the sensibilities of the growth rate, sλ .
The sensitivites to the entries of A are given by :
Figure 1: A typical matrix population model. The life
cycle graph is on the left and the projection matrix of the
population is on the right.
The quantity of information which can be obtained from
such a simple model is impressive but need careful interpretation. The first thing to keep in mind is that matrix population models are usually best not used as predictive models.
Indeed, as we will see, under assumptions which are almost
always verified, the population dynamic converges toward an
exponential growth, ever increasing or decreasing. Rather,
matrix population models must be seen as a way to obtain
information about the current population.
We now provide a basic set of results about matrix population models, without proving them. The proofs, as well as
various examples, can be found in Caswell [10].
sλ (aij ) =
vi w j
vw
Since λ can be viewed as an integrated measure of fitness at
the population level, the sensitivies are of particular interest
when studying evolution. However, they have some shortcomings when studying the life cycle: for instance, non-existent
transitions (null entries in the matrix) can have non-null sensitivities – which might be of great interest if studying how
new transitioned evolved in the life cycle, but is not in many
cases (e.g: if that transition is biologically impossible). This
is why it is sometimes more desirable to work with proportional (rather than absolute) contributions, and this is why
elasticities were introduced. The elasticity of λ to aij is :
Eigen-elements
Projection matrices are non-negative, and they are usually
irreducible, aperiodic, hence primitive. The Perron-Frobenius
theorem therefore applies, and the matrix has a dominant
eigenvalue λ with associated right and left eigenvectors w
and v, respectively. When the matrix is not irreducible, this
is usually due to a post-reproductive class or a source-sink
metapopulation, in which case although v will contain a null
entry, w will still be positive. When the projection matrix is
periodic with period h, there are h dominant eigenvalues and
the population oscillates with period h. However, it is possible
to consider the matrix Ah (which maps the population from
time t to time t + h) to come down to the primitive case.
We therefore assume in the rest of the document that A is
primitive.
For t large enough, we have :
n(t + 1) ∼ λn(t) ∝ w
∂λ
∂aij
eλ (aij ) =
aij ∂λ
λ ∂aij
It is given by :
aij vi wj
(2)
λ vw
The elasticities of λ to the entries of A sum to one, and
the elasticities to null entries are null. They are therefore
interpreted as ‘the relative contribution of a transition to λ’.
As we will see, it is possible to give another interpretation
for them.
eλ (aij ) =
We now give a very brief overview of the last tool used in
this work: finite-state discrete-time markov chains.
(1)
Markov chains
That is, the population vector converges toward the
eigenspace generated by w and then behaves exponentially,
increasing if λ > 1, decreasing if λ < 1 (the case λ = 1, although mathematically negligible, is of biological interest and
correspond to a population at demographic equilibrium).
λ can therefore be interpreted as the asymptotic growth rate
of the population, and w as its stable distribution: assuming
it is scaled so that its entries sums to one, its entries are the
fraction of individuals in each class.
The interpretation of v is less direct, but several independent arguments point to interpreting it as the reproductive
value of the population – that is, its entries indicate the relative contribution of each class to the overall size of the population.
The literature on Markov chains is huge, and once again we
simply recall a few results that we will need. These results
were taken from Amaury Lambert’s lesson LV388 at the École
Normale Supérieure: ‘What a biologist should not ignore’, of
which there is to my knowledge no written version. However,
these are basic results which are to be found in any introductory textbook on Markov chains.
The model
By ‘Markov chains’ we actually mean ‘Finite-stage discrete
time Markov chains’. They model a stochastic system occupying different states numbered {1, ..., n} according to the
following rule: the state of the system at time t + 1, X(t + 1),
depends only on its state at time t, X(t).
3
The formalism of Markov chains is very much like that
of matrix population models, except for a transpose when
writing the system:
both. This has notably been done by Cochran and Ellner,
and by Cushing and Yican [3, 4]. Both paper use the same
technique, which consist in separing the reproductive transitions from the survival ones. A complete presentation of this
technique is given in Caswell’s chapter 5, although he uses
the terms ‘fertilities’ and ‘transitions’ (and therefore different notations). The results derived in this report also depend
on the reproductive-survival decomposition, but for different
purposes than the markovization of the life cycle, which does
not rely on it. Therefore, we discuss it later in paragraph the
A = R + S decomposition.
x(t + 1) = x(t) P
Here, x is the probability distribution of the system among
the different states at time t (i.e. xi = P [X(t) = i]) and
P is the transition matrix, whose entries are the transition
probabilities:
pij = P [X(t + 1) = j | X(t) = i]
Motivation and principle
The term life cycle is rarely questioned. But from an individual’s perspective, it is not justified as life is but a linear
journey from birth to death. I therefore started my reflection
by trying to find what was cyclic about the life cycle. I was
greatly helped in this by the concepts that Richard Dawkins
popularized under the terms replicator and vehicle [11]. But
as it happens, the resulting method does not rely on these
concepts:
Stationary distribution and return time
The transition matrix entries, pij , are probabilities. As a result, P is non-negative. Moreover, {X(t) = i | i ∈ {1, ..., n}}
is a complete system of events, so the rows of P sum to one.
Provided P is irreducible aperiodic, the Perron-Frobenius theorem applies, and we can assert that P has a dominant eigenvalue λ with corresponding right and left eigenvectors e and
π, respectively. Since x is bounded and x(t) ∼ λt x(0), we
have λ ≤ 1. But the fact that the rows of P sum to one
gives that 1 is eigenvalue of P. Therefore, λ = 1, with corresponding right eigenvector e> = (1, ..., 1) (e> denoting the
transpose of e). Finally, π is interpreted as the stationary
probability distribution of X.
The return time of state i is the random variate Ti equal to
the time it takes the system to reach state i, having started
from it:
Ti = min {t ≥ 1 | X(t) = i} ,
0.6
1
X(0) = i
0.15
The mean of T is given by E [Ti ] = π1i . Indeed, in k steps,
we will have visited state i on average πi k times. The average
time between two visits is therefore πki k = π1i . More generally,
the return time to a set of states S is :
1
E [TS ] = P
πs
2
A
0.35
2
0.2
0.7
3
0.75
0.15
4
0.5
B
(3)
s∈S
time
Results
This work is based on the idea (which was suggested to me
by Stéphane Legendre) of defining the generation time as a
return time. To do so, I have made use of two techniques: the
first consists in transforming a population projection matrix
in a Markov chain transition matrix and was first found by
Demetrius [5]; the second consists in transforming the life cycle graph in order to study the properties of its arcs by studying the properties of the nodes of the transformed graph. This
has to my knowledge never been used in matrix population
models.
Figure 2: A, fictive life cycle (with a post-reproductive
class) – B, demographic process compatible with A represented as a branching process. See text for more precisions.
Figure 2.B is an example of a one-sex (or no-sex) demographic process followed at the individual level and represented as a branching process. Oblique lines mark births,
and with the vertical line that follow they correspond to a
given individual. The death of the individual occurs when
the vertical line stops. Different stages correspond to different colors, and Figure 2.A is a life cycle compatible with the
Markovization
Given the facts that: a. Markov chains and matrix population models share a common formalism, and b. the literature
on Markov chains is much more vast than that on matrix population models, it is natural to try to make the link between
4
demographic process depicted in Figure 2.B. This diagram
contains all the information about the demographic parameters of the population.
What we would like is to convert a matrix population model
into a model that would enable us to study the properties of
diagrams such as that of Figure 2.B This might seem impossible, as a matrix population model is built at the population
level whereas the branching diagram necessitates information
at the individual level, but it can actually be done assuming
that the vital rates of an individual depend only on its class –
which is the working hypothesis of matrix population models.
To do this, we work on genealogies, i.e. we trace down
the ancestors of individuals rather than their descendants. In
other words, we inverse the time in Figure 2.B so as to go up
instead of down. As a consequence, the process becomes much
easier to study as every individual comes from one and only
one individual (this is because matrix population models do
not take sex into account: when they are not used for asexual
organisms, only one sex is taken into account. It is then
almost always the female sex as it is often easier to determine
the offspring of a female than that of a male. The model
is then said to be female-based. There exist two-sex models,
but they include frequency-dependence – and therefore nonlinearity – and are much less used). However, by doing this we
also leave out the dead-end branches. This is not a problem
given that the absence of interaction between individuals is a
working hypothesis of matrix population models: as a result,
these post-reproductive individuals do not have any impact
on the population.
To model these genealogies, we use a Markov chain whose
system is an abstract particle following the branches of the
genealogy as it moves back in time, and whose states are the
classes of the population model. Before giving the mathematical expression describing this Markov chain, we give possible
interpretations of this abstract particle.
tation suffers from the same problem as the replicator-based
one when it comes to one-sex models.
Finally, one need not have any opinion on evolution to interpret the results obtained by markovization as they are rigorously identical (though simpler and more complete) to those
obtained when defining the generation time as ‘the mean age
of mothers at birth’, which is a commonly used measure [2].
Indeed, the quantity we compute in this work is directly be
interpreted as ‘the time between two births when tracing a genealogy’ and is therefore exactly the age of mothers at birth.
We will further discuss this point and compare our results to
those existing in the discussion section.
Mathematical expression
We now give the mathematical expression of markovization.
In order to be as independent as possible of any biological
context, we will talk of individuals rather than replicators or
germ-line cells in what follows.
We want to know the probability pij that an individual in class
P i comes from class j. At time t, there are
ni (t) =
k aik nk (t − 1) individuals in class i, aij nj (t − 1)
of which come from class j. Therefore:
pij =
aij nj (t − 1)
ni (t)
Assuming the population is at its stable stage distribution,
n(t) ∼ λn(t − 1) ∝ w (eq. 1). Replacing in the previous
expression,
aij wj
pij =
(4)
λwi
The resulting expression is independent of t, and the matrix
P = (pij ) is the Markov matrix we want (the fact
P that its
rows sum to one is trivial when replacing λwi by j aij wj in
P
P
1
j pij = λwi
j aij wj ) Since it is assumed that w > 0 (see
paragraph eigen-elements), the formula is always valid. Note
that the apparent absence of transposition is due to the fact
that two transpositions have been performed: one to reverse
the time, and the other to switch from the formalism of the
matrix population models to that of the Markov chains.
This formula had been found by Demetrius [5]. Further
exploration of the literature on Markov chains also made me
notice that this formula is very similar to that of what is
known as the ‘reverse Markov chain’ However, this technique
does not seem to have been used to markovize other matrices.
Interpretations
As already stated, I addressed the problem of generation time
with an evolutionary rather than demographic point of view,
and I was initially strongly influenced by the concepts of replicator and vehicle [11]. I wanted to define the generation
time as ‘the time a replicator spends in a vehicle’ and use
Markov chains to model the moves of a replicator from vehicle to vehicle. For a no-sex model, this aim is achieved by the
markovization technique. However, for a one-sex model, the
situation is a bit more complicated: indeed, we only take into
account replicators that stay into the sex of interest. But a
replicator is most likely to shift from vehicle of one sex to the
other. If the time spent in vehicles of both sexes is different,
then the computed quantity will not be ‘the time spent in a
vehicle’. It could still, however, be interpreted as ‘the time a
mitochondrial replicator spends in a vehicle’, given that for
many organisms paternal mitochondrial DNA is usually not
transmitted.
However, one need not embrace the ideas of replicator and
vehicle to interpret markovization: some people such as my
supervisor, Stéphane Legendre, might prefer to interpret it
as the average time spent before germ-line cells (rather than
replicators) experience reproduction. However, this interpre-
Stationary probability distribution
The stationary probability distribution of the previously defined Markov chain (eq. 4) is given by:
πi =
vi w i
vw
(5)
Proof:
pIt is obvious that the entries of π sum to one.
We now
check that π is a left eigenvector of P:
X
j
5
πj pij =
X vi wi aij wj
j
vw λwi
=
vi X
aij wj
λvw j
But since w P
is a right eigenvector of A associated with
eigenvalue λ, j aij wj = wi . Substituting into the previous
expression,
X
vi wi
= πi
πj pij =
vw
j
6
322.38
y
Hence the result.
0.872
30.17
0.750
3.448
0.023
Line graph
0.167
0.125
In this section we describe the second method we use to study
the generation time. This method enables the creation of an
object called the line graph. Although the line graph was already know in graph theory, the fact that it was mostly studied in a different context (non-directed, non-weighted graph)
and for different reasons (namely, per se) made it virtually
impossible to find the needed results, which had to be found
again. Simple though they are, these results and the corresponding proofs are therefore given here. To my knowledge,
this is the first time the line graph has been used in matrix
population models.
1
0.238
0.013
3
0.125
0.007
4
0.245
5
0.038
0.008
0.966
0.010
0.007
0.008
2
Motivation
Now that we have markovized the projection matrix of the
population, it is tempting to define the generation time as the
return time to one (or a subset) of its nodes – those that correspond to newborns. However, there are several cases in which
the newborns are ambiguously defined and are therefore not
an adequate choice: for instance, in certain models (e.g: sizeclasses, instar-classes), the new individuals can spend several
year in the newborn class before growing/maturing into another class (Figure 3.A). In that case, there is a self-loop on
the newborn node which as a survival component. Another
case which can be encountered is that of retrogression in sizeclass models (Figure 3.B). Finally, complex life cycle can also
lead to such situations, as exemplified by Figure 4.
Figure 4: A real-life example of a complex life cycle including survival into ‘newborn’ stages. The organism is
the teasel (Dipsacus sylvestris) and the classes are: 1.
dormant seed year 1, 2. dormant seed year 2, 3. small
rosette, 4. medium rosette, 5. large rosette, 6. flowering
plant. Taken from [10] (corrected from [12])
By contrast, reproductive arcs can be defined unambiguously, although there might be both a reproductive and a survival arc leading from a node to another (see the A = R + S
decomposition). It would therefore be desirable to be able to
work with arcs in the same way as we work with nodes. To
this end, we build a new graph whose nodes correspond to
the arcs of the original graph.
A
1
2
Construction
3
e as follow:
Let G be a graph. We build its line graph, noted G,
e Crucially,
• To each arc of G corresponds one node of G.
if there are several arcs of G going from one node to an
other, each of them will be associated with a distinct
e
node of G.
B
1
2
• A node a of Ge corresponding to arc (i → j) of G leads to
node b of Ge corresponding to arc (k → l) of G if and only
if j = k.
3
• The weight of an arc (a → b) of Ge is the weight of the
arc of G to which corresponds b.
Figure 3: Theoretical examples illustrating how survival
into ‘newborn’ classes might occur. The red arcs are reproductive arcs, the blue ones correspond to survival into the
newborn class. A, standard size-classified model – B, sizeclassified model with retrogression (an example of multigraph).
Figure 5 gives an example of a simple graph and its line
graph.
6
A
b
a
1. Connectedness:
Assuming G contains no isolated nodes,
b
c
d
c
d
G strongly connected ⇐⇒ Ge strongly connected
(9)
Proof:
e with corresponding arcs
pLet a and b be two nodes of G,
B
a
a
a
b
1
c
a
2
in G (i → j) and (k → l), respectively. If G is strongly
connected,
b
b
c
e
d
d
e
b
(i → j) → (m1 → m2 ) → ... → (mn−1 → mn ) → (k → l)
c
d
i.e. a
Conversely, if G is not strongly connected, ∃(j, k), j 6 k.
Since it is assumed that G has no isolated nodes, there exists
an arc containing j and an arc containing k (either as head
e
or tail). Let a and b be the corresponding nodes of G,
respectively. Then a 6 b. Indeed, if we had a
b, we
could explicit a path from a to b and proceed as before to
exhibit a path from j to k. As a result,
Properties
The following properties and their proofs can be written using the formalism of adjacency matrices. Although this might
seem the most natural option to someone used to matrix population models, it is much easier to reason in terms of graphs
– and the corresponding notations are much lighter. Indeed,
if we wanted to work with adjacency matrices and be as rigorous as possible without losing any generality, we would have
to introduce quite heavy notations to deal with the fact that
the graph we are working on can be a multigraph (i.e. can
contain more than one arc going from one node to another),
as will become apparent in paragraph the A = R + S decomposition. We therefore introduce some notations and give a
characterization of matrix products and left eigenvectors ‘in
terms of graphs’:
We note WG the weighting function of graph G, i.e. the
function which maps an arc to its weight. By a slight abuse
of notation, we also define the weighting function so as to map
a node of the line graph Ge to the weight of the corresponding
arc in G. We introduce the function t which maps an arc to
its tail. Finally, we note χ(a) the set of the in-arcs of a. With
these notations, the weighting function of Ge is defined by :
WGe(β) = WG (a)
b. As a result,
G strongly connected =⇒ Ge strongly connected
Figure 5: Constructions of line graphs. The letters indicate the weights of the arcs, and an arc and its corresponding node are the same color. A, a subgraph and its
corresponding subgraph in the line graph – B, a graph and
its line graph. This example shows that a multigraph is
transformed into a simple graph.
∀β ∈ χ(a),
j → m1 → ... → mn → k
It is then clear that
e
e
k, i.e. ∃(m1 , ..., mn ),
j
G not strongly connected =⇒ Ge not strongly connected
Hence the equivalence.
2. Period
G and Ge have the same period
y
(10)
Proof:
that
pTo show this, we show a stronger but obvious result:
e
each cycle in G has a corresponding cycle in G of same
length (and vice versa). This is because a cycle, which can
always be uniquely defined by its arcs, can also be uniquely
defined by its nodes when there is at most one arc going
e Thus, a
from one node to another, as is the case for G.
cycle of G described by its arcs defines a unique cycle of Ge
by considering the corresponding nodes, and vice-versa. As
the number of arcs of a cycle equals its number of nodes,
the cycles all have the same lengths. A fortiori, the GCD
of the lengths are identical.
y
3. Primitivity
(6)
G primitive ⇐⇒ Ge primitive
For any adjacency matrix G representing graph G, a product by a row vector x can be written:
X
(xG)b =
xt(a) WG (a)
(7)
(11)
Proof:
pThis is a direct consequence of (eq. 9) and (eq. 10)
y
4. Sums of the rows
Let Sums denote the set of the sums of the rows of the adjacency matrix of a graph. Then,
a∈χ(b)
Thus, the fact that x is a left eigenvector of G with respect
to the eigenvalue µ translates into:
X
xt(a) WG (a) = µxb
(8)
e
Sums(G) = Sums(G)
a∈χ(b)
7
But since x is a left eigenvector of G associated with respect
to eigenvalue µ, we have from (eq. 8)
X
xt(b) WG (b) = µ xa
Proof:
pThe weights of the out-arcs of node a in Ge are exactly those
of out-arcs of the head of the corresponding arc in G. Theree associated with node a
fore, the entries of the row of G
are those of the row of G associated with the head of a
(completed with zeros for columns associated with nodes to
which a doesn’t lead). Hence the result.
b∈χ(t(a))
Finally,
y
Corollary:
e
eG
x
= µ xa WG (a) = µ x
ea
a
e is a Markov matrix
G is a Markov matrix ⇐⇒ G
Corollary :
Let P be a Markov matrix with stationary probability distrie is a Markov matrix with stationary probability
bution π. P
distribution:
π
ea = πt(a) WG (a)
(14)
Proof:
pAssuming WG ≥ 0, this corresponds to the case where:
Sums(G) = {1, ..., 1}
y
Proof:
e is a Markov matrix from (eq. 12). From (eq. 11), P
e
P
is primitive. Thus, the Perron-Frobenius theorem applies
and the left eigenvector associated with eigenvalue λ = 1
whose entries sum to one can be interpreted as the statione Since π is associated with
ary probability distribution of P.
e from (eq. 13). Thus, all we have
eigenvalue λ = 1, so is π
e sum to one.
to do is check that the entries of π
X
X
X
X
π
ea =
πt(a) WP (a) =
πi
WP (b)
5. Eigen-elements
The question of eigen-elements is slightly more complex and
has not been fully explored during this intership. However,
the only result we need is the stationary distribution of the
line graph of a Markov chain, and this can be found easily.
More general results have been derived, but as they are of no
use for the study of the generation time, they are not given
here. They can, however, be found in appendix A.3.
Let µ be an eigenvalue of G, and x be a left eigenvector of
G associated with eigenvalue µ. Then:
p
a
• µ is an eigenvalue of Ge
e, whose entries are given by:
• row vector x
x
ea = xi WG (a),
where i is the tail of a
is a left eigenvector of Ge associated with µ
y
Which is the desired result.
(12)
a
i
b∈out(i)
Where out(i) is the set of the out-arcs of i. But as P represents a Markov chain,
X
WP (b) = 1
(13)
Proof:
b∈out(i)
pFrom (eq. 7), we have:
Therefore,
X
X
e
eG
x
=
x
et(β) WGe(β)
a
a
β∈χ(a)
As for every arc β pointing to a we have WGe(β) = WG (a)
e a:
(eq. 6), we can substitute both expressions in that of (e
xG)
X
e
eG
x
=
x
eb WG (a)
y
i
But we also have:
• P [X = i] = πi
h
i W (a) if i = t(a)
P
e
• P X=a|X=i =
0 else
b∈χ(t(a))
Substituting x
eb by xt(b) WG , we get
X
e
eG
x
=
xt(b) WG (b) WG (a)
a
πi = 1
i
The proof above isn’t satisfactory as it relies on (eq. 13)
which shows that the stated expression is correct without explaining where it comes from. Here is a less rigorous but more
intuitive justification: π is the stationary probability distribution of the nodes of P. We want the stationary probability
distribution on its arcs. Noting [X = i] event ‘being on node
e = a] event ‘using arc a’, we have:
i’ and [X
h
i X h
i
e =a =
e = a | X = i P [X = i]
P X
P X
b∈χ(t(a))
a
X
because π is a probability distribution.
Since (by construction) Ge is not a multigraph, for every arc
β of Ge pointing to node a, there is a unique node b of G so
that β = (b → a). And since the set of theses nodes for
every β pointing to a corresponds exactly to the set of the
arcs of G pointing to the tail of a,
X
X
function of t(β) =
function of b
β∈χ(a)
π
ea =
h
i
e = a = πt(a) WP (a) if i = t(a). This is how
As a result, P X
the expression of (eq. 13) was initially found.
b∈χ(t(a))
8
Generation time
instance, one must decide (depending on the particular population studied) whether to include vegetative reproduction
in the reproductive arcs.
This decomposition has been frequently used in matrix population models, because R is a convergent matrix which can
be interpreted as describing the moves of individuals from
class to class (by adding a ‘dead’ class, R can be transformed
into a transient Markov matrix which describes the fate of an
individual before being absorbed into the ‘dead’ state). The
fact that R is convergent (i.e. limk→∞ Rk = (0)) makes it
possible to compute expressions such the geometric series of
R and its derivatives, making this technique a powerful tool.
See Cochran and Ellner [3] or Cushing and Yican [4] for detailed example, or Caswell [10] for a more general discussion
of the technique.
However, we do not use this decomposition to this end.
We only use it to partition the nodes of the line graph into
‘reproductive’ and ‘survival’. This done, the generation time
can be defined as:
We now have all the mathematical tools we need to study
the generation time. All we have to do is to apply them to
the projection matrix of the population. The procedure is as
follow :
1. Identify the reproductive arcs
2. Markovize the life cycle
3. Build the line graph of the Markov chain
4. The generation time is then defined as the return time to
the nodes of the line graph corresponding to reproductive
arcs.
Interestingly enough, 2. and 3. need not be done in that
order as the markovized line graph of the life cycle is the
same as the line graph of the markovized life cycle (a proof of
this in the diagonalizable case can be found in appendix A.3).
By contrast, it is important to identify the reproductive arcs
before building the line graph because a reproductive arc and
a survival arc having the same tail and head will correspond
to disctinct nodes in the line graph.
At this point, the reader may wonder how to markovize a
multigraph, as (eq. 4) was given in terms of the adjacency
matrix of the graph. The general formula for markovization
is :
WA (a)wt(a)
(15)
WP (α) =
λwh(a)
T = min {t ≥ 1 | X(t) ∈ R}
(16)
Where X is the random variate of the Markov chain associated with the graph. In other word, the A = R + S decomposition, the markovization and the construction of the line
graph have enabled us to translate the initial definition of the
generation time into a mathematical definition which we can
compute. We now perform the calculation.
e
The transformed graph P
The definition of the transformed graph can be easy to apply
formally. Two examples of constructions can be found in the
appendix: that for the age-classified model (A.1) and that
for the standard size-classified model (A.2). Even when a
formal expression is hard to obtain, it is always possible to
get a numerical expression, as the numerical calculation of
the transformed graph is easy to program. About half of
my internship was spend writing a program performing this
calculation (as well as various others). The resulting code can
be found at [13].
e to find its stationHowever, one need not even compute P
ary probability distribution. Indeed, from (eq. 5), the stationary probability distribution π of the markovized graph P
i wi
. From (eq. 14), the stationary distribuis given by πi = vvw
tion of the associated line graph is given by π
ea = πt(a) WP (a).
Substituting WP (a) from (eq. 15),
Where α is the arc of markovized graph P corresponding to
arc a of A; h and t are the functions mapping an arc to its
head and tail, respectively; and w is the right (or left, if not
using the formalism of matrix population models) eigenvector
of A with respect to the dominant eigenvalue λ. This formula
reduces to (eq. 4) when A is not a multigraph. We did not
give it in the first place so as to avoid introducing to many
notations at once. We hope that after having familiarized
with reasoning on graphs rather than matrices in section Line
graph, the reader will find it obvious.
We now detail the steps described above.
The A = R + S decomposition
The arcs of the graph representing the life-cycle can be partitioned into two sets: R, which contains arcs associated with
the creation of new individuals, and S, whose elements correspond to the survival of individuals either in the same stage
or in a different one (growth, migration in multi-site models...). Even if there were several arcs of one type going from
one node to another, it would still be possible to aggregate
them into a single arc whose weight is the sum of the weights
of aggregated arcs, because we are only interested in whether
the arcs are reproductive or not.
Therefore, it is possible to decompose the adjacency matrix
A as A = R + S, where R and S are the partial adjacency
matrices associated with the sets of arcs R and S, respectively. Note that this does not correspond to decomposing
the graph into two subgraphs, as the nodes are shared.
Of course, the decomposition is dependent on the biological context and the definition of ‘reproduction’ adopted. For
π
ea =
=
vt(a) wt(a) WA (a)wh(a)
vw
λwt(a)
vt(a) wh(a) WA (a)
λvw
(17)
Stéphane Legendre noticed that from (eq. 2), this can also be
written:
π
ea = eλ (a)
(18)
It is very surprising that nobody has found this result before,
as it has important consequences for the interpretation of the
elasticities. Indeed, a direct interpretation of (eq. 18) is that
the elasticity of λ to aij is the frequency of traversal of the
arc (j → i).
9
Distribution of T
Homo sapiens
0.6
0.4
0.2
(19)
0.20
0.0
e SS
P
0.10
e SR
P
!
0.00
e RS
P
Density
e=
P
e RR
P
10
eRR contains the weights of the arcs
Where the submatrix P
going from R to R (similar notations hold for the other submatrices). Similarly, the stationary probability distribution
can be written:
50
4
6
8
10
12
Astrocaryum mexicanum
0.012
0.030
0.006
(20)
e R can be scaled so that it sums to one. We note the reπ
sulting vector ω. It is interpreted as a stationary probability
distribution on R, i.e. ω a = P [X = a | X ∈ R]
0
50 100
200
300
0
Generation time (years)
100
300
500
Generation time (years)
Figure 6: Distributions of T for various models. The
model for H. sapiens being age-classified with a maximum
cycle length of 10, its generation time has a finite support.
This is not the case for the other models. O. orca and
A. mexicanum are examples of standard size-class models, which typically exhibit a ‘negative binomial-shaped’
distribution. The model for D. sylvestris is less classic
and show an example of a generation time with infinite
support but which in practice takes only a few values. The
data used for these calculation can be found in [10]
The distribution of T is given by:
e RR e,
ωR P
t−2 P [T = t] =
ω
e
e RR
e SR e,
P
P
R PRS
2
0.000
40
0.015
eS
π
30
0.000
eR
π
20
Orcinus orca
Density
e=
π
Dipsacus sylvestris
0.30
e can be written:
The adjacency matrix of P
t=1
t≥2
(21)
where e is a column vector of the same length as ω full of
ones (it only sums the entries of the row vector by which it
is multiplied)
Mean of T
The distribution of T given in (eq. 21) is probably the most
complete information one can have about the generation time.
But it is hard to interpret and therefore of little interest in
itself (it can be used to compute other quantities though, see
section other results).
However, it is possible to compute the mean of T . Indeed,
combining (eq. 3) and (eq. 18), we have:
Proof:
pThe rigorous demonstration of (eq. 21) is simple but tedious. Therefore, we only give the main idea without detailing the calculus:
• T = 1 means that we have gone from a reproductive node to another. The probability of this is the
sum (hence e) of the probabilies of leaving a particular
node in R (hence ω R ) times the probability of using an
arc going from that node to another node of R (hence
e RR )
P
T = E [T ] = P
1
eλ (a)
(22)
a∈R
This is, with (eq. 18), the main result of this work. However,
this expression is only valid in the simple graph case. But
one can also use (eq. 17) to compute T. By noting that ∀a ∈
R, WA = rt(a)h(a) (where r comes from A = R + S), we have:
• T = t ≥ 2 means that we have left R (one projection interval), then spent t − 2 projection intervals in S
before returning to R (one projection interval) the vectors and matrices interpretation is as straightforward
as that of the case T = 1.
T=
λvw
vRw
This expression in valid in the general case.
Other results
The implications of the previous results have not yet been
fully explored. In this section, we give two results which might
benefit from further investigation.
1. Lebreton’s formula
This paragraph is due to Stéphane Legendre as I had no
y
The formula given in (eq. 21) makes it possible to compute
the distribution of T numerically. Some examples are given
in Figure 6.
10
Existing measures of the generation time
knowledge of Lebreton’s formula for age-classified models.
Here we generalize this formula to any model described by
a simple graph: Let c be a parameter multiplying the weights
of all reproductive arcs. Then:
eλ (c) =
The generation time is intuitively thought of as ‘the time
between two generations’. However, this definition might be
interpreted in more that one way. The demographist Coale is
generally credited for clarifying this definition [2]. He defined
three measures of the generation time:
1
T
Similarly, if d is a parameter multiplying the non-reproductive
arcs:
1
eλ (d) = 1 −
T
Proof:
• The difference in age between mothers and
daughters in the population. When the population
is at its stable age distribution, this is the age of mothers
at birth. Of course, it is also possible to define another
measure as ‘the mean difference in age between fathers
and sons’. But, as most models are female-based, this is
of less interest.
pSince c multiplies the weights of all reproductive arcs, we
have aij = c rij + sij . The fact that A is a not a multigraph
translates into rij 6= 0 =⇒ sij = 0 and rij 6= 0 =⇒ sij =
0 Hence,
(
∂λ ∂aij
∂λ ∂aij
if rij 6= 0
aij c
=
∂aij ∂c
0
else
• The age of mothers at birth in a given cohort.
This definition is very similar to the previous one, but
the population need not be at its stable age distribution.
• The time it takes for the population to grow by a
factor of its net reproductive rate. The net reproductive rate is the mean of the number of offspring an
individual is expected to produce during its life. Thus,
given the net reproductive rate, it is possible to map the
population from one ‘generation’ to another, rather than
from one time interval to another. The time scale of this
process is obtained by taking the ratio of the logarithm
of the growth rate by that of the net reproductive rate.
This can be interpreted as a measure of the generation
time.
As a result,
X
c X ∂λ ∂aij
c X ∂λ aij
c ∂λ
=
=
=
eλ (a)
λ ∂c
λ i,j ∂aij ∂c
λ
∂aij c
rij 6=0
a∈R
From (eq. 22),
1
T
The corresponding identity for eλ (d) can be derived by using the fact that the elasticities sum to one (this classic
result can be easily derived from the fact that as λ is a homogenous function ofP
degree one of the aij ’s, it follows from
∂λ
= λ)
Euler’s formula that
aij ∂a
ij
eλ (c) =
As of today, these are still the three common measures of the
generation time. The first and the second ones can be viewed
as formalizing the concept of ‘distance between generations’,
whereas the third is more similar to a ‘renewal rate’ of the
population.
For matrix population models, mathematical expressions
are available for each of these definitions. The mean age of
mothers at birth as been known for age-classified models since
Leslie [16]:
m
X
T=
iφi λ−i
y
2. Variance, entropy...
(eq. 21) makes it possible to perform numerical computation
of other values characterizing T , such as its variance, etc.
Although in some cases it is possible to get a formal explicit
expression of some of these quantities (see, for instance A.1
and A.2), the resulting expressions are rather obscure and
hard to interpret (but this might simply be due to a failure
to simplify them).
One of these quantities is the Shannon Entropy [14] of T ,
defined as:
X
S (T ) = −
P [T = t] log (P [T = t])
(23)
i=1
Where m is the number of age-classes of the model, and φi is
the net fertility of age-class i, i.e. the product of the survival
to class i times the fertility of class i.
However, a formula for the general case was unknown before
1992, until Cochran and Eller’s paper [3]. Their expression of
the mean age of mothers at birth is given by:
t
This quantity is intriguing because it as the same interpretation as the population entropy defined by Demetrius [15],
but is different from it for non age-classified models. We will
discuss this further in the discussion.
Pm
yi wi γi
Pi=1
m
i=1 wi γi
Discussion
Where m is the number of classes of the model, w is the stable
stage distribution, y is the distribution of ages in classes and
γ is the fecundity in ‘newborn equivalents’ of the classes. y
and γ are given by:
In this section, we first compare our expression of the generation time to the existing ones. We then discuss ways in which
this work could be extended.
11
Pm
(I − λ−1 S)−2
((I −
λ−1 S)−1 )
b
ij j
ij bj
(Rw)j
, where bj = Pm
i=1 (Rw)i
Comparison of the three measures of
generation time
300
yi =
j=1
Pm
j=1
and
250
●
●
100
(v, R and S having the same meaning as before).
150
200
(vR)i
, where vref is a newborn stage of reference
vref
Years
γi =
mu
tau
T
50
The interpretation of this quantity being the same as that
we computed, we expect both expressions to be identical.
And indeed, although this hasn’t been shown analytically,
numerical computations show that it is the case. As a result, we have found a much simpler expression of an already
known quantity. Our derivation is very different from that of
Cochran and Ellner. It is also more general at it envisions
the generation time as a random variate and provide its full
distribution. Moreover, our method is independent of any biological context and might be applied to other models than
matrix population models, as we will discuss in the next section. However, Cochran and Ellner’s method enabled them to
derive an expression for the mean age of mothers at birth in a
given cohort, as well as the time it take the population to grow
by a factor of its net reproductive rate (although with respect
to this quantity, both Cushin and Yican’s expression [4] and
its derivation are more simple and intuitive).
●
●
0
●
a
O.
●
●
orc
A.
xic
me
ii
s
is
ule
um
siz
ien
str
ca
an
as
ve
ap
l
a
g
s
y
.
a
.
s
C
H
G.
D.
Figure 7: Comparison of the three measures of the generation time for six organisms. mu is the age of ‘mothers’
at birth in a newborn, tau is the time it takes for the
population to grow by a factor of its net reproductive rate,
and T is the quantity which we calculated and showed to
equal the mean age of mothers at birth in the stable-stage
population. This illustrates the fact that the three values
can be quite different and that Coale’s relation does not
hold in the general case.
At this point, one might wonder how these three quantities
relate to one another, or whether one of them is superior in
anyway to the others, or might be better suited to a particular
use.
Perspectives
Equation 18 and the new interpretation of elasticities it leads
to are very intriguing. Although this is only valid in the
simple graph case, this is not a real limitation as far as matrix population models are concerned since multigraphs are
exceptional in this field (but this might in part be due to a
bias in the construction of projection matrices, as it is easy to
imagine situations best described by multigraphs). It is therefore very surprising that this result had never been found, as
elasticities and their interpretation have been a major subject of study [17,18]. As a result, it remains to be determined
whether this new interpretation of elasticities as the frequency
of use of arcs (rather than the classic, more indirect ‘relative
contribution to the growth rate’) can lead to novel interpretations of existing results.
Another point which will have to be studied further is the
question of the entropy of T . Demetrius [1, 15] defined a
quantity he calls population entropy as
Z ∞
S=−
p(x) log p(x) dx
As of how the quantities relate to one another, this remains
to be explored. However, Coale says that in humans, they are
very similar, and that the time it takes for the population to
grow by a factor of its reproductive rate is approximately the
mean age of mothers of a given cohort at birth and the average age of mothers at birth at the stable stage distribution [2].
Caswell backs up this assertion [10], but we found this assertion to be erroneous in the general case: the discrepancies
between the values can be huge, and the alleged relation does
not hold, as show by Figure 7.
It is harder to say that one of these quantity is better suited
for a particular use than the others. However, the mean age
of mothers at birth in a given cohort has the advantage that
it does not rely on the population being at its stable stage
distribution, a very strong and often questionable hypothesis
– But it is also more challenging to interpret. As of the mean
age of mother at birth in the population and the time it takes
for the population to grow by a factor of its net reproductive
rate, although we have no definitive argument nor experimental support for this, the way they were derived suggests that
the former might be more adapted when studying evolution,
and the latter better suited from a purely demographical perspective.
0
where p is the probability distribution of the age of mothers
at birth – that is, of the random variate we have called T . As
a result, it seems natural to adapt this definition to matrix
population models by using the Shannon entropy (the discrete
analogue of the differential entropy) of T and define S as
12
we did in (eq. 23). However, for matrix population models,
population entropy has been defined as
X
S = −T
πi pij log pij
be used to test for allometric relations in ecosystems in an
effort to scale-up the metabolic theory of ecology to the level
of the ecosystem.
i,j
The S = log(T) relation
(P being the previously defined Markov chain and π its stationary probability distribution).
Although it is easy to show that both definitions coincide
for age-classified models (see appendix A.1), numerical computations show that they are different for other models. It is
therefore interesting to compare both definitions, but this is
outside the scope of this document: population entropy relies
on far more complex mathematical concepts than those used
here, and there is a vast literature about its biological significance [1, 15, 19]. However, it is worth noting a few points:
150
100
0
• Many studies restricted themselves to the age-classified
case – as a consequence, many result about population
entropy apply to the Shannon entropy of the generation
time.
●
●
50
Population entropy: S = HT
●
● ●●●
●● ●
●
●●●
●● ●
●●●●
●●
● ●●
●●
● ●●● ●●
●
●
●
● ● ●
●●
●
●●
●
●
● ●
●
● ●
●
●
3
4
● ●
● ●
●●
●
●●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
1
• A 2009 paper [1] gave theoretical as well as experimental
support for the relation S = a log T + b. But this relation
is best observed when using the Shannon entropy of the
generation time, as shown by Figure 8.
●
●●
●
2
Shannon entropy of T
• The interpretation as the uncertainty over the genealogies of individuals also holds for the Shannon entropy of
the generation time, and is arguably more straightforward. Moreover, the Shannon entropy of the generation
time seems a more natural translation of the population
entropy expressed as the differential entropy of the age
of mothers at birth.
5
6
●
●● ●
●
● ●●
●
0
● ●
●
● ●
●
●
1
2
3
4
5
log(T)
Given the importance of population entropy [19], determining how both definitions relate is going to be the main
extension of this work.
Figure 8: The relation ‘S = a log T + b’ using the two
definitions of the entropy. The relation is best observed
with the Shannon entropy of the generation time, for
which a = 1.15 and b = 0.052, with a correlation coefficient superior to 0.98. The data are from 56 population
projection matrices randomly chosen in the literature.
The time is expressed in projection intervals rather than
years, since the entropy is independent of the actual
duration of the projection interval. However, this does
not make much difference since here, only 7 models have
projection intervals different from one year.
Conclusion
We greatly simplified the mathematical expression for the
mean age of mothers at birth in matrix population models.
But beyond its mathematical simplicity, our expression leads
to a new interpretation of the generation time as well as the
classic quantities known as the elasticities. As a consequence,
our results call for more work on the subject.
This study was motivated by a biological question. Thus,
each step of the reasoning that lead to our results was strongly
influenced by the biological context. However, retrospectively, the underlying biology can be seen as a guide rather
than a set of constraints: indeed, our results hold for any
model described by a directed weighted graph. It is therefore natural to wonder whether there are other areas where
they could be applied. Stéphane Legendre pointed out that
trophic networks are a good candidate: if one had enough
data about the flows of matter between the populations of an
ecosystem and its environment to model the trophic network
with a weighted graph, it would be possible to use our formula to define a generation time for ecosystems. This could
13
[18] H. de Kroon, A. Plaisier, J. van Groenendael, and
H. Caswell. Elasticity: the relative contribution of demographic parameters to population growth rate. Ecology,
67, 1986.
References
[1] L. Demetrius, S. Legendre, and P. Harremoës. Evolutionary entropy: A predictor of body size, metabolic rate and
maximal life span. Bulletin of Mathematical Biology, 76,
2009.
[19] L. Demetrius and S. Legendre. Evolutionary entropy predicts the outcome of selection: Competition for resources
that vary in abundance and diversity. Theoretical Population Biology, 83, 2013.
[2] A.J. Coale. The Growth and Structure of Human Populations. Princeton University Press, 1972.
[3] M.E. Cochran and S. Ellner. Simple methods for calculating age-based life history parameters for stagestructured populations. Ecological Monographs, 62(3),
1992.
[4] J.M. Cushing and Z. Yican. The net reproductive value
and stability in matrix population models. Natural Resources Modeling, 8(4), 1994.
[5] L. Demetrius. Demographic parameters and natural selection. Proceedings of the National Academy of Science,
71, 1974.
[6] F. Bienvenu, L. Demetrius, and S. Legendre. A general
formula for the generation time. arXiv:1307.6692, 2013.
[7] C.D. Meyer. Matrix Analysis and Applied Linear Algebra, chapter 8. SIAM, 2000.
[8] P.H. Leslie. On the use of matrices in certain population
mathematics. Biometrika, 33(3), 1945.
[9] L.P. Lefkovitch. The study of population growth in organisms grouped by stages. Biometrics, 21(1), 1965.
[10] H. Caswell. Matrix Population Models: construction,
analysis, and interpretation. Sinauer, 2nd edition, 2001.
[11] R. Dawkins. The Selfish Gene. Oxford University Press,
1976.
[12] P.A. Werner and H. Caswell. Population growth rate and
age versus stage-distributions models for teasel. Ecology,
58.
[13] Link to the software used:
http://francois.bienvenu.free.fr/generation_
time.
[14] C.E Shannon. A mathematical theory of communication.
Bell System Technical Journal, 27(3), 1948.
[15] L. Demetrius. Statistical mechanics and population biology. Journal of Statistical physics, 30(3), 1983.
[16] P.H. Leslie. Some further notes on the use of matrices in
population mathematics. Biometrika, 35(3-4), 1948.
[17] H. Caswell. Optimal life histories and age-specific costs
of reproduction: two extensions. Journal of Theoretical
Biology, 107, 1984.
14
Appendix
A.1 The age-classified model
Prerequisites
In age-classified models, individuals can either survive – in which case they get one projection interval older – or
die. Moreover, we consider the age of individuals to be bounded – that is, we know that individuals cannot get
older than a certain age, either because we have never observed this or because we have physiological reasons to
think so. Finally, in the more general case, each age-class has a specific fertility. As a result, the age-classified
model is:
f1 f2 · · · fn−1 fn
s1 0 · · ·
0
0
0 s2 · · ·
0
0
A=
..
.. . .
..
..
.
.
.
.
.
0
···
0
sn−1
0
Here, there is only one, well-defined newborn stage. Thus, there is no need to use the line-graph, since the
return to a reproductive transition is equivalent to the return to the newborn stage.
The characteristic equation, det(A − λI) = 0, can be made explicit by recursively developing the determinant.
After rearranging the terms, one obtains:
X
φi λ−i = 1, where φi = s1 ... si−1 fi is the age-specific net fertility
i
The left and right eigenvectors can be obtained either by solving the corresponding systems directly or by first
making use of the z-transform to simplify the life-cycle. They are given by:
vi =
n
X
j−1
Y
j=i
k=i
!
sk
i−(j+1)
fj λ
and wi =
i−1
Y
!
sk
λi−1
k=1
Markovization, generation time and entropy
Applying (eq. 4) to markovize A gives:
φ1 λ−1
1
P = 0
..
.
0
φ2 λ−2
0
1
..
.
···
···
···
..
.
φn−1 λ−(n−1)
0
0
..
.
0
···
1
φn λ−n
0
0
..
.
0
The fact that P is a Markov chain is matrix is made apparent by the characteristic equation,
P
i
φi λ−i = 1.
Here, the distribution of the return time to the newborn stage is easily obtained without even bothering to
find the stationary probability distribution of P because the only probabilistic event in this Markov chain is the
‘departure’ from the newborn stage: being in stage 1, the chain goes to stage i with the probability φi λ−i . Then,
it comes back to stage 1 deterministically in (i − 1) steps, and the return time associated with this cycle is i. As
a result, the probability distribution of the return time is given by:
P [T = i] = φi λ−i
Computing the mean of T gives the classic result about the mean age of mothers at birth:
X
T=
iφi λ−i
i
I
Interestingly, for these models, the Shannon entropy coincide with the population entropy. Indeed, it is given
by:
X
S=
φi λ−i log φi λ−i
i
which is Demetrius’ formula.
A.2 The standard size-classified model
Here, unlike in the age-classified model, individuals have more than one possible fate when they survive: they
can either grow or not. Some models also include retrogression in smaller stages, or even ‘rapid growth’ from
one stage to a non-immediately-larger one, but we do not take them into account and we limit ourselves to what
we call the ‘standard size-classified model’:
f1 + s1
g1
A=
0
.
..
···
···
..
.
f2
s2
g2
..
.
0
fn−1
0
0
..
.
..
.
···
0
gn−1
fn
0
0
0
sn
This model makes it interesting to use the line graph to work with reproductive transitions rather than
newborn stages since, although unique, the newborn stage is somewhat ambiguous: indeed, an individual in the
newborn stage can either come from a reproductive event (i.e. has just been born) or not (i.e. has survived
without growing).
Here, we explicit all the intermediate results as an example of construction, although they are not needed to
compute the mean generation time since one can directly use T = vRw
λvw
The A = R + S decomposition gives:
f1
0
R=
0
.
..
f2
0
0
0
···
···
..
.
0
..
.
..
.
···
fn
0
0
0
0
fn−1
0
0
..
.
0
s1
g1
and S
0
.
..
0
s2
g2
..
.
0
0
···
···
..
.
..
.
···
0
0
0
..
.
gn−1
0
0
0
0
sn
The right eigenvectors of A is given by:
i−1
Q
wi =
gk
k=1
i
Q
(λ − sk )
k=2
Thus, the markovization of A gives:
σ1
γ1
P=
0
.
..
0
0
σ2
γ2
..
.
0
···
···
..
.
..
.
···
0
0
0
..
.
γn−1
0
φ1
0
0
0
+0
.
0 ..
σn
0
φ2
0
0
..
.
0
···
···
..
.
..
.
···
φn−1
0
0
..
.
0
φn
0
0
0
0
si
σi = λ
γi = 1 − σi+1
where
(g1 ...gi−1 )fi
φi = λ(λ−s
2 )...(λ−si )
e RR , P
e SR ,
We now build the corresponding line-graph. To do so, it is much easier to build the submatrices P
e
e
PRS and PSS :
II
The only reproductive arc leading another reproductive arc is
(including itself). As a result,
φ1 φ2 · · ·
0
0 ···
e RR =
P
..
..
..
.
.
.
0
0 ···
the arc φ1 . It leads to every reproductive arc
φn
0
..
.
0
Likewise, the only to survival arcs leading to reproductive arcs
φ1 φ2 · · ·
φ1 φ2 · · ·
0 ···
e SR =
P
0
..
..
..
.
.
.
0
0
···
e SR .
are γ1 and σ1 , hence P
φn
φn
0
..
.
0
e RS . The arc φi leads to the node i, whose out-arcs are σi and γi−1 . If we choose to place
We now turn to P
e RS , we have:
the arcs in the order σ1 , γ1 , σ2 , γ2 , ... when writing P
e RS
P
σ1
0
=0
..
.
0
γ1
0
0
σ2
0
···
...
···
0
0
0
..
.
0
0
0
..
.
0
0
···
0
γn−1
σn
That is, (n − 1) 1x2 blocks and a single element in upper left corner.
e SS comes from the fact that σi and γi+1
Finally, P
are 2x2 blocks, except for the upper left and bottom
σ1 0
0
σ1 0
0
0 γ1 σ2
0 γ1 σ2
e SS =
P
0 0
0
0 0
0
..
.
0
0
lead to i, whose out-arcs are σi and γi . So this time, there
right corners:
0 0 ···
0
0
0 0 ···
0
0
0 0 ···
0
0
0 0 ···
0
0
γ2 σ3 · · ·
0
0
γ2 σ3 · · ·
0
0
..
..
.
.
··· 0 0
γn−1 σn
e SS is particularly interesting. Indeed, from (eq. 21), if we manage to compute its powers, we
The matrix P
have a closed expression for the full distribution of T . Unfortunately, I didn’t manage to do it. Nevertheless, it is
possible to find a closed expression for the moments of T . Indeed, nth moment of T can be expressed as a linear
P∞ e t
combination of the E T i , i ∈ {1, ... , n}. But from (eq. 21), given t=1 t P
SS , there is a closed formula for
i
e
E T . And since PSS is convergent (as a submatrix of an irreducible Markov matrix), we have:
∞ t
−2
X
e SS = P
e SS I − P
e SS
t P
t=1
Example: the variance of T is given by:
V (T ) =
ωR
e RR + P
e RS
P
∞
X
t
2
t=2
III
e SS
P
t−2
!
!
e SR
P
e
−
2
E [T ]
where
∞
X
t−2 −3
−2 −1
e SS
e SS I − P
e SS
e SS
e SS
t2 P
= I+P
+2 I−P
+ I−P
t=2
Since we have
e SS
I−P
−1
1 + ρ1
ρ1
ρ1
= ρ1
..
.
ρ1
ρ1
0
1
1
1
..
.
0
0
1 + ρ2
ρ2
..
.
0
0
0
1
..
.
···
···
···
···
0
0
0
0
..
.
1
1
ρ2
ρ2
1
1
···
···
1
1
0
0
0
0
..
.
0
ρn
where
ρi =
σi
1 − σi
, the formula for the variance of T given above is a closed expression. However, expanding it does not seem to
simplify the expression and as a result it is little informative.
A.3 More on the line graph
Eigenvalues
Let G be the adjacency matrix of a connected graph G. If G is diagonalizable, then
e ∪ {0}
• Sp G = Sp G
e are the same
• The multiplicities of the non-zero eigenvalues of G and G
e is diagonalizable
• G
Proof:
p1.
e the multiplicities of the eigenvalues of G
e being superior or equal to that of
From (eq. 13), Sp G ⊂ Sp G,
the corresponding eigenvalues of G.
e Then G
e has exactly n different rows. Indeed, let a
2. Let n be the number of rows of G and ñ that of G.
and b be two arcs of G such that h(a) = h(b). Then, the corresponding nodes of Ge have the same out-arcs and
e are identical. As a result, there are at most n different rows
therefore the rows to which they correspond in G
e
e corresponding them
in G. But if h(a) 6= h(b), then since h(a) and h(b) have different out-arcs, the rows of G
are different. Since there are n nodes in G, there are n possible heads for the arcs of G. And because G is
e Combining
connected, each of them is ‘used’ at least one. As a result, there are at least n different rows in G.
both inequalities gives the desired result.
e ≤ n, i.e. 0 is an eigenvalue of G
e of multiplicity k ≥ ñ − n.
3. The direct consequence of 2. is that rank G
4. We now use the fact that G is diagonalizable: This means that the sum of the multiplicities of its eigenvalues
e we have found so far is n + k. Since we must have
is n. Thus, the sum of multiplicites of the eigenvalues of G
e is exactly ñ. As a result,
n + k ≤ ñ and since k ≥ ñ − n, the sum of the multiplicities of G
e and their multiplicity, i.e. Sp G = Sp G∪{0},
e
• We have found all the eigenvalues of G
and the multiplicities
e
of the non-zero eigenvalues of G and G are the same.
e is diagonalizable.
• G
y
e have the same dominant eigenvalue. Although this property was
A consequence of this result is that G and G
shown in the diagonalizable case, empirical testing suggests that it is always true.
IV
Commutative property
e M and L commute for primitive, diagonalizable matrices.
Let M : G 7−→ P and L : G 7−→ G.
Proof:
pThis proof is elementary, but it is complicated by the difference of convention between matrix population
models and Markov chains.
e = (L ◦ M) (A). From (eq. 15) and from the definition of
Let A be a primitive, diagonalizable matrix and P
the line-graph,
WA (b) wt(b)
if a → b in P, i.e. b → a in A
λ wh(b)
p̃ab =
0
else
Now let Q = (M ◦ L) (A). We have:
ãab =
WA (a) if b → a in A
0
else
Thus, a direct application of (eq. 4) gives:
qab =
WA (a) w̃b
λ̃ w̃a
0
if b → a in A
else
But we have shown in the previous section that, since A is primitive and diagonalizable, λ̃ = λ. Moreover,
(eq. 13) shows that w̃a = wt(a) WA (a). As a result,
wt(b) WA (b)
WA (a) wt(b) WA (b)
WA (a) w̃b
=
, since t(a) = h(b).
=
λ wt(a) WA (a)
λ wh(b)
λ̃ w̃a
y
Therefore, p̃ab = qab . Hence the result.
V
Notations cheat-sheet
P
7 : (xG)b =
General notations
xt(a) WG (a) (product by a row vector
a∈inarcs(b)
A : life cycle graph
A : population projection matrix (adjacency matrix of A)
n : population vector
λ : dominant eigenvalue of A (growth rate)
v : left eigenvector of A (reproductive value)
w : right eigenvector of A (stable stage distribution)
P / P : markovized graph / matrix
π : left eigenvector of P (stationary proba. distri.)
e : line graph of P / corresponding adjacency matrix
e/P
P
expressed with the formalism of graphs)
8:
P
(definition of eigenvectors with the formalism of graphs)
9 : G strongly connected ⇐⇒ Ge strongly connected
10 : G and Ge have the same period
11 : G primitive ⇐⇒ Ge primitive
e : left eigenvector of P (stationary proba. distri.)
π
e
ω R : stat. proba. distri. on the reproductive nodes of P
eλ (a) : elasticity of λ to arc a
T : generation time (random variate)
T : mean generation time (E [T ])
e is a Markov matrix
12 : G is a Markov matrix ⇐⇒ G
e
13 : x̃a = xt(a) WG (a) (expression of the eigenvectors of G)
14 : π
ea = πt(a) WG (a)
e
(stationary probability distribution of P)
Graph notations
WG (a) : weight of arc a in graph G
h(a) : head of arc a
t(a) : tail of arc a
i → j : i leads to j, i.e. there is an arc going from i to j
i
j : i is reaches j, i.e. there is a path going from i to j
W (a)w
A
t(a)
(general formula for the
15 : WP (α) =
λwh(a)
markovization – valid in the multigraph case)
16 : T = min {t ≥ 1 | X(t) ∈ R} , X(0) = i
(definition of the generation time)
Other notations
17 : π
ea = eλ (a)
(application of equation 15 to population projection matrix)
x ∝ y : x is proportional to y
GCD : greatest common divisor
Sp A : spectrum of A (multiset of its eigenvalues)
18 : π̃a = eλ (a) (the frequency of traversal an arc is the
elasticity of λ to this arc)
Equations
e=
19 : P
1 : n(t + 1) ∼ λn(t) ∝ w
(convergence of the population stage distribution).
2 : eλ (aij ) =
3 : E [TS ] =
e=
20 : π
aij vi wj
λ vw
P1
xt(a) WG (a) = µxb
a∈inarcs(b)
e RR
P
e RS
P
e SR
P
e SS
P
eR
π
eS
π
!
e RR e,
ωR P
t−2 21 : P [T = t] =
ω
e
e RR
e SR e,
P
P
R PRS
πs
s∈S
(expression of the mean return time to the set of nodes S).
t=1
t≥2
(closed expression for the probability distribution of T )
4 : pij =
aij wj
λwi
(markovization – simple graph case).
22 : T = E [T ] =
P 1
eλ (a)
(expression of the mean genera-
a∈R
v w
i j
5 : πi = vw
(stationary probability distribution of the markovized matrix)
λvw
tion time in the simple graph case) and T = vRw
(expression
of the mean generation time in the general case)
6 : ∀β ∈ inarcs(a), WGe(β) = WG (a)
(weights of the arcs of the line graph)
P
23 : S (T ) = − t P [T = t] log (P [T = t])
(Shannon entropy of the generation time)
VI