BS3050 Physiology of Sport and Exercise
advertisement

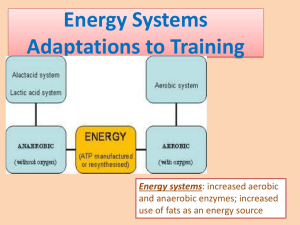
BS3050 Physiology of Sport and Exercise Muscle Metabolism Aims: to explain how ATP is generated for muscular contraction and the factors which limit and regulate the availability of ATP in the muscle cell Learning Outcomes: An understanding of the role of ATP in muscle contraction Knowledge of the sources and use of ATP in muscle contraction: Alactic and lactic anaerobic sources and aerobic sources. An understanding of the roles of glycolysis, citric acid cycle, oxidative phosphorylation and -oxidation in the provision of energy Molecular Mechanism of Skeletal Muscle Contraction The major proteins involved in muscle contraction are actin and myosin which are assembled into filaments that undergo transient interactions and slide past each other to bring about the contraction. This process requires ATP, which is consumed in the process with the concomitant formation of ADP and inorganic phosphate (Pi). I will not go into the detailed mechanism but just explain how ATP is consumed during the contractile process and what effect this has on the metabolism within the muscle. Some Basic facts about muscle contraction: The signal from the nerve triggers an action potential in the muscle cell plasma membrane. The signal is relayed to the sarcoplasmic reticulum (sr), a sheath of anastomosing vesicles forming a network which surrounds each myofibril Muscle contraction is initiated by a sudden but transient (less than 30 millisec) rise in cytosolic Ca2+ which is released from the sr Ca2+ changes are detected by a protein called troponin C which interacts with the actin filaments and initiates the contraction In the resting state the interaction between actin and myosin is inhibited but when cytosolic Ca2+ increases then ATP can bind to the myosin which has ATPase activity (ATP ADP + Pi) The ATP binding causes a change in conformation of the myosin head causing it to move along the actin filament but the hydrolysis of ATP causes the myosin to regain its original conformation generating the power stroke. Thus a molecule of ATP is consumed for every binding event and conformational change at multiple points of contact between myosin and actin moves the filaments with respect to one another and therefore the muscle contracts. Each cycle the consumption of 1 molecule of ATP generates about 3 – 4 pNewtons (1 Newton = Kg.m.s-2 ) of force and moves the filaments about 5 – 10 nm relative to one another. Phosphoryl Transfer Potential of ATP The standard free energy of hydrolysis of ATP and other metabolites, which is a convenient method of measuring the ‘energy’ associated with different metabolites (see Table 14.1), reveals that ATP is not the intermediate with the highest phosphoryl transfer potential but its properties allow it to function efficiently as a carrier capable of transferring energy from one molecule to another. ATP is therefore an immediate donor of energy which is very rapidly turned over. The total ATP in the body at any one time is only about 100g yet a resting individual consumes about 280 g ATP per minute rising to about 500 g per minute on strenuous effort. In a marathon 60 kg of ATP may be consumed! Muscle energy ( i.e. ATP) is derived from 3 main sources: Alactic Energy Source (it is called alactic because lactic acid does not accumulate in the muscle) Anaerobic source (for up to about 30 sec depending on the intensity of effort) This source is mainly used for explosive sports such as sprinting, javelin, discus, weightlifting, golf swing, tennis serve, kicking a football, diving etc. The capacity is about 23 - 36 kJ with maximum power output of 4 - 12 kW for about 8 sec. The ATP source for this phase is via the cytoplasmic ATP present in the muscle cytoplasm and that generated from creatine phosphate in an anaerobic step catalysed by creatine kinase: Creatine Phosphate + ADP Creatine + ATP G0 = -8kJ/mol i.e. the equilibrium lies well to the right and this maintains the muscle adenylate (ATP, ADP and AMP) pool in the ATP form. In the resting muscle the relative concentrations are: ATP (4mM), ADP (0.13mM), Creatine phosphate (25mM) and creatine (13mM) and When explosive muscle contraction occurs it is initially the ATP which is utilised closely followed by the creatine phosphate. In a 100 meter sprint, the ATP in muscle is sufficient for less than 1 sec whilst the regeneration of ATP by creatine phosphate is sufficient for about 4 sec of effort and the remainder of the race is dependent on anaerobic glycolysis. NMR studies on contracting muscle shows that creatine phosphate content is depleted by about 90% while the ATP content is maintained. Because of the varied roles of ATP in cells it would not be desirable for the ATP to become too depleted. Creatine phosphate is resynthesised by the creatine phosphate shuttle which exchanges creatine phosphate synthesised in the mitochondria for the creatine in the cytoplasm. A second reaction can provide ATP catalysed by myokinase: 2ADP ATP + AMP This reaction is at equilibrium and is therefore driven to the right by ATP consumption. The reaction can even be driven further to the right by the deamidation of AMP to IMP but this depletes the adenylate pool. After a rest of a few minutes the ATP and creatine phosphate can be restored to their resting levels by aerobic metabolism. Lactate-generating Anaerobic source of ATP This source is used for short spells of strenuous effort lasting less than 3 minutes as in gymnastics, 200 – 800 metre races in athletics, 100 – 200 metre races in swimming events. The capacity of the muscles for such effort is about 95 – 120 kJ and the maximum power output is approx. 3 – 4 kW but this cannot be sustained for more than a minute or so. After this effort it can take 1 to 2 hrs to recover depending on the fitness of the individual. This source of energy depends on the glycogen degradation and the rate of glycolysis in the muscle, which generates ATP from ADP, and results in the accumulation of lactate and H+ ions which accumulate in the cytoplasm. Lactate cannot be utilised quickly enough by the muscle mitochondria and hence it accumulates but it is the fall in pH in the muscle cytoplasm (rather than lactate accumulation per se) which decreases the efficiency of contraction of muscle. This process is self-limiting because the acidosis builds up before the glycogen has been completely utilised. The accumulation of H+ ions appears to inhibit various enzymes involved in the provision of ATP rather than have a direct effect on the contractile mechanism although an accumulation of Pi does have a direct effect. H+ ions are removed via the blood stream where they are ‘mopped up’ by the Bicarbonate/Carbonic Acid buffering system. The lactate can be converted back into glucose via a process known as gluconeogenesis but this can only occur in the liver and hence lactate accumulates in the blood circulation after exercise. It should be emphasised that lactate itself is not toxic it is a by-product of anaerobic glycolysis, which is rich in energy and indeed is a vital intermediate which can be used by many tissues but not used very efficiently by skeletal muscle in the short term. The lactate concentration in the blood is sometimes used as a marker to measure muscle function but it is a very indirect and inaccurate way of measuring muscle acidosis. There is a concept of an ‘anaerobic threshold’ of 4mM lactate where the muscle has to switch to aerobic metabolism to supply ATP but in reality this has little or no validity. There is a limited store of glycogen in muscle –it makes up about 1.5% of the wet weight of the muscle in a rested individual i.e. about 420g in 28 Kg of muscle found in a standard 70 Kg man. An additional glycogen store of about 500 g is found in the liver: this is used to maintain blood glucose in the short term. Glycogen is metabolised by the enzyme phosphorylase Glycogen(n) + Pi Glycogen(n-1) + Glucose-1-phosphate. Glucose-1-phosphate can be readily converted to Glucose-6-phosphate, which can be readily converted to lactate by anaerobic glycolysis. An alternative source of Glucose6-phosphate is the uptake of glucose from the blood via a transporter (Glut4 – insulindependent) in the muscle plasma membrane and its phosphorylation by hexokinase. This is a relatively slow process and requires the expenditure of ATP. Phosphorylase is present in the cytoplasm but anchored to the sarcoplasmic reticulum and becomes activated very rapidly in response to the increase in cytoplasmic Ca2+ which is the signal for muscle contraction. Hence muscle contraction is very closely coupled to the supply of glycolytic intermediates which can be used immediately to generate ATP. McArdles disease, an inherited deficiency of muscle phosphorylase, results in inability to perform strenuous efforts results in painful muscular cramps. No accumulation in H+ or lactate but NMR studies indicate that there is an increase in ADP on exercise which correlates with the onset of cramps. Anaerobic Glycolysis Conversion of glycogen or glucose to pyruvate and lactate (Fig 16.3) Occurs in 3 stages : (a) Conversion of Glucose or glycogen to fructose 1,6-bisphosphate. This requires an input of energy in the form of ATP (either 2 ATP in the case of glucose or 1ATP for glycogen) (b) Cleavage of the 6-carbon sugar to give two molecules of the high energy intermediate 1,3 bisphosphoglycerate (c) The generation of ATP by phosphoglycerate kinase and puruvate kinase Overall 2 molecules of ATP are consumed and 4 molecules of ATP produced ( i.e. net 2 mol ATP) during anaerobic glycolysis of glucose whilst glycolysis from stored glycogen yields 3 mol of ATP. Therefore in the short term, glycogen produces more energy than glucose. Why does lactate accumulate? Lactate accumulates in the anaerobic muscle because of the reductant, NADH, which is produced at an earlier step in glycolysis, reacts with pyruvate to form lactate which accumulates at high concentrations until it can be removed. This reaction is catalysed by lactate dehydrogenase: Pyruvate + NADH Lactate + NAD+ Lactate can be removed by conversion to pyruvate and the metabolism of the latter by the Citric acid cycle by muscle mitochondria but before this can happen the accumulated NADH must also be removed by the mitochondria. A second mechanism for the removal of lactate (which does not depend on the removal of muscle NADH) involves the Cori Cycle in which lactate is transported to the liver where it can be reutilized to make glucose by the process of gluconeogenesis. Aerobic Mitochondrial Generation of ATP For longer periods of exercise lasting more than 3 minutes: middle and long distance running, football and hockey matches etc the main source of energy is from aerobic sources with perhaps bursts from anaerobic sources. The capacity of the muscles for such effort is almost unlimited a but the maximal power is only of the order of 1 - 2 kW, and rather limited over the longer time period. The recovery time after such exercise may be 24 - 48 hr. Aerobic power depends on the ability of an individual to (a), absorb, (b), transport and (c), utilize oxygen. In turn (a) and (b) are a function of the efficiency of the lungs and cardiovascular system whilst (c) is dependent on the number and volume of the muscle mitochondria. Aerobic endurance ultimately depends on the ability of the mitochondria to utilize oxygen in the oxidation of cellular substrates to CO2 and H2O The carbohydrates represent 50 – 60% of the aerobic substrates mainly in the form of glucose (4 –5 mM, fasting level) circulating in the blood and glycogen, the polymeric form of glucose which is stored in an inert form in the muscle cytoplasm (1.5 - 2% of total weight in the resting muscle). The total energy available in the circulating glucose is 170 – 250 kJ while the muscle glycogen stores represent about 2500 - 5000 kJ. Most other carbohydrates in the diet need to be transformed into glucose, and then to glycogen, before they can be used by muscle The lipids represent about 25 – 30 % of the anaerobic energy consumption. Storage fat is in the form of triacylglycerol which is an ester made of glycerol (a glycolytic intermediate) and fatty acids. These fatty acids may be absorbed in food or synthesised in the liver or adipose tissue and stored as triacylglycerol. This reserve is large (420,000kJ in adipose tissue) in most people although it clearly varies from person to person – even the supremely fit have 5 - 20 % of body weight. In order to be utilised these fatty acids have to be mobilised and used by mitochondria by a process of -oxidation to generate ATP. This oxidation requires more oxygen than the metabolism of carbohydrate but generates much more ATP (see later) and thus more power. The link between glycolysis and the citric acid cycle is irreversible The citric acid cycle is the final common pathway for the aerobic oxidation of fuel molecules. Entry of substrates into the cycle is via Acetyl CoA which is formed from pyruvate in the mitochondria by an enzyme complex known as pyruvate dehydrogenase: Pyruvate + Coenzyme A + NAD+ AcetylCoA + CO2 + NADH This is an irreversible reaction in humans (indeed in all animals); once acetyl CoA is formed it can only be used for two main purposes: generation of energy via the citric acid cycle or conversion to fat which can be laid down in adipose tissue until required. The Citric Acid Cycle and Oxidative Phosphorylation For the complete oxidation of the glycolytic product, pyruvate to CO2 and H2O requires the function of mitochondrial metabolic cycle - the citric acid cycle (Fig 17.15). The fuel substrate is acetyl CoA which is converted to CO2 . The citric acid cycle requires a supply of the intermediate oxaloacetate to combine with Acetyl CoA to give citrate, in a series of oxidation reactions which yield GTP, NADH and FADH2. The latter two are also energy-rich molecules because they contain a pair of electrons having a high transfer potential (i.e. they are strong reductants). These can be used to generate ATP by the process of oxidative phosphorylation as the result of the transfer of these electrons to O2 via a series of electron carriers. This generates a gradient of H+ across the inner mitochondrial membrane which is used to drive ATP synthesis from ADP and Pi. The rate of oxidative phosphorylation is determined by the need for ATP as indicated by the accumulation of ADP and the availability of oxygen to the mitochondria. Table 18.4 shows the ATP yield from the complete oxidation of glucose. For every molecule of glucose converted to CO2 : 2 mols of ATP are generated by glycolysis, 2 mol of ATP by the operation of the citric acid cycle and the remaining 26 mol ATP by oxidative phosphorylation. Generation of energy from fat As the immediate sources of energy ATP, carnitine phosphate and glycogen become depleted the muscle turns to another source of energy triacylglycerol, which becomes very important in endurance events. Blood glucose remains available but this is necessary to maintain the function of all the other tissues in the body e.g. the brain, and so can only be used sparingly in muscles. Endurance training increases the individual’s capacity to mobilise, deliver and oxidise fatty acid derived from triacylglycerol stored in the adipose tissue. Mobilization of triacylglycerol involves hydrolysis with the release of fatty acids (and glycerol). Fatty acids are long chain hydrocarbon acids which are transported in the blood stream as a complex with albumin to the sites where they can be metabolised, including muscle, by a process of -oxidation which yields Acetyl CoA, which can enter the citric acid cycle and produce energy as described. The utilization of Acetyl-CoA derived from fat does require some metabolism of carbohydrate as well to replenish the pool of oxaloacetate which is essential for the operation of the citric acid cycle. In addition -oxidation also yields NADH and FADH2 which can also be harnessed to make ATP via oxidative phosphorylation. The complete oxidation of a single molecule of palmitic acid yields 106 molecules of ATP (c.f. 30 molecules for glucose) and hence fat is a much more efficient storage form of energy although it is not immediately accessible for muscular work. The percentage of energy supplied by fatty acids varies from about 30 to 80 % according to the nutritional state, the intensity of training and the intensity of physical training and becomes the primary source in endurance events. Furthermore exposure to low carbohydrate diets accentuates the dependence on fatty acids because the muscle adapts to using this source to generate ATP. -oxidation of fatty acids by the liver can also generate ketone bodies (e.g. acetoacetate) which can also be used by skeletal muscle to generate Acetyl CoA and therefore ATP. Cardiac muscle, in particular, is dependent on a continuous supply of ketone bodies as a fuel source, which clearly needs a reliable and constant source of energy! Generation of energy from proteins Proteins also play a role as energy yielding substrates during endurance training and heavy physical work. Proteins, mainly in muscle and liver can be mobilized in the form of aminoacids, which then undergo deamination to convert them into products which can be metabolised via the glycolysis and citric acid cycle. Some amino acids are converted to pyruvate, others to acetyl CoA whilst others enter the citric acid cycle directly. Skeletal Muscle Fibre Types Voluntary skeletal muscle is under conscious control. Each fibre is an enormous, multi-nucleate cell, formed by fusing hundreds of myoblasts end-to-end. They show a striated pattern, reflecting the regular arrangement of sarcomeres within each cell. Type I fibres : depend on oxidative metabolism, slow twitch, relativly low ATPase. The contraction cycle procedes at a relatively slow pace. The fibres have high aerobic capacity but low anaerobic capacity. Used in endurance exercise and maintaining posture – resistant to fatigue. Dark red in colour because of the presence of myoglobin, and the highly vascularized tissue, with large mitochondria. Type II fibres: glycolytic, fast twitch, high anaerobic capacity, recruited for higher intensity exercise in addition to type I Type IIa fast twitch, react more quickly than Type I, ATP hydrolysed more quickly by ATPase in myosin. They have both aerobic and anaerobic metabolisms well developed and are moderately fatigue-resistant. Their level of stored glycogen is high, hence capable of anaerobic glycolysis. The fibres are active in walking and sprinting. Type IIb very fast twitch, hydrolyse ATP very rapidly, almost devoid of mitochondria, blood vessels and myoglobin (hence white). High glycogen content - The fibres have a high anaerobic capacity, recruited for very intense efforts (weight lifting, throwing etc.) and fibres fatigue very quickly. Training can increase the volume of these fibres via the synthesis of new muscle proteins. Relative proportion of different types of fibre is genetically determined and helps to account for difference between individuals athletic performances but characteristics of fibres can be changed to some extent by training. Chickens use their legs (red meat) for walking and standing for long periods of time, whilst their wings (white meat) are used for brief bursts of activity during flying. This perfectly illustrates the differences between fast and slow-twitch fibres. In mammals the different types of fibre coexist in each muscle. References: Exercise Physiology 5th ed. (2001) McArdle, Katch and Katch Chapters 5, 6 and & 7 612.044 MAC ( Bedford Library Quarto Holdings) Physiology of sport and exercise (1999) Wilmore, Jack H. 612.044 WIL( Bedford Library Quarto Holdings) Biochemistry 5th Ed (2002) Berg, Tymockzo and Stryer Chapters 14, 16, 18 and 22. (any basic Biochemistry textbook published in the last 10 years will suffice) British Medical Bulletin Vol 48 (3) Sports Medicine p 477 – 95, 569-91 Basic and Applied Sciences for Sports Medicine ( Bedford Library Journal Section) Human Anatomy and Physiology (2003) Marieb PowerPoint slides on muscle contraction http://www.aw-bc.com/ppt/marieb_hap/chap09c.ppt