have we forgotten about the role of nerves in asthma?
advertisement

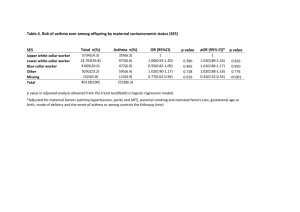
doi:10.1594/eaacinet2007/EO/4-300307 http://www.eaaci.net HAVE WE FORGOTTEN ABOUT THE ROLE OF NERVES IN ASTHMA? Ulrike Raap MD Dept. of Dermatology and Allergology, Hannover Medical University, Germany Correspondence: mail@ulrike-raap.de doi:10.1594/eaacinet2007/EO/4-300307 http://www.eaaci.net ABSTRACT Bronchial asthma is well defined by the clinical characteristics including chronic airway inflammation, recurrent reversible airway obstruction and development of airway hyperresponsiveness as pathogenic components. Several studies have shown that genetic, environmental, immunologic, and pharmacologic factors contribute to the development of this complex disease. However, there is one important component which had long been forgotten but now steps out of the shadow into the spotlight of bronchial asthma: the neuronal network. Our approach to understanding bronchial asthma needs to be broadened to include alterations in the function of this neuronal network with afferent nerves that supply airways and modulate neuroimmune interactions. Changes in the activity of these nerves offer a possible mechanism by which asthmatic subjects are uniquely responsive to a wide range of physiological and chemical stimuli- which may offer novel therapeutic strategies. Key words: Bronchial asthma, eosinophils, neuronal network, neuropeptides, neurotrophins doi:10.1594/eaacinet2007/EO/4-300307 http://www.eaaci.net INTRODUCTION Inflammatory cells such as T-helper 2 cells, eosinophils, and cytokines play a critical role in orchestrating and perpetuating inflammation in bronchial asthma. It is now widely accepted that airway inflammation is one of the key factors underlying the pathogenesis of asthma (1). Even though many promising single agents antagonising specific cytokines or key effector cells in asthma have been investigated for the treatment of asthma, no single drug is able to cure asthma. Therefore, it is tempting to argue that besides cytokines and key effector cells one important compartment deserves more attention within the airways: the neuronal network. Airway nerves not only present the backbone for neuronal interactions but most importantly they represent the hotline for neuroimmune interactions between key target effector cells and nerves in asthma. The airways are supplied with a network of nerves that can cause bronchoconstriction or relaxation. Constriction of airway smooth muscle is controlled by sensory and motor neurons innervating the airways and the lung. In addition to effects on airway smooth muscle tone, the nerves regulate many aspects of airway function including mucus secretion, bronchial microcirculation, vascular permeability and most importantly have a bidirectional interaction with immune cells. The autonomic nerves that regulate many aspects of airway function can be functionally subdivided into cholinergic, adrenergic and nonadrenergic non-cholinergic (NANC) pathways. PARASYMPATHETIC NERVE SYSTEM The parasympathetic nerve system is the dominant neuronal pathway in the control of airway smooth muscle tone. Stimulation of cholinergic nerves causes bronchoconstriction and mucus secretion. Three types of muscarinic receptors (M1-3) have been investigated thus far. Dysfunction of the M2 receptor has been proposed as a mechanism leading to excessive bronchoconstriction and mucus secretion in asthma (2). Indeed, some asthmatics represent a dysfunctional M2 receptor (3). Although some studies have concluded that the cholinergic nerves are not important in allergic asthma (4), others have shown that anticholinergics given intravenously are effective bronchodilators demonstrating vagal nerve blockade (5). In this regard, viral infection has been shown to be clearly doi:10.1594/eaacinet2007/EO/4-300307 http://www.eaaci.net associated with airway hyperresponsiveness that is vagally mediated (6). M2 receptor dysfunction and vagally mediated hyperresponsiveness are absolutely dependent on an inflammatory response. The eosinophil granulocyte has been questioned to contribute to airway hyperreactivity. However, eosinophils have been found in high concentrations in the vicinity of airway nerves (7) and were found to be responsible for the loss of M2 receptor function by the release of major basic protein (MBP) leading to excessive bronchoconstriction (8), underlining their importance for neuroimmune interactions in asthma. THE NON-ADRENERGIC NON-CHOLINERGIC NERVE SYSTEM The only bronchodilator pathways in human airways are presented by the inhibitory non-adrenergic non-cholinergic (i-NANC) nerves containing vasoactive intestinal peptide (VIP) and nitric oxide (NO). Although a dysfunction of i-NANC nerves has been proposed in asthma, thus far, no difference in the i-NANC responses have been found between asthmatics and healthy subjects (4). This is in contrast to the excitatory-NANC (e-NANC) nerves containing tachykinins including substance p (SP) and neurokinin-A (NKA) which contribute to bronchoconstriction, mucus secretion, vascular hyperpermeability, cough and vasodilatation (9,10). In guinea pigs SP induces airway hyperreactivity (11). Whereas in a mice model for allergic airway inflammation it has been clearly shown that NK1 receptor- to which SP binds with high affinity- knock-out animals promote feature of remodelling such as goblet cell hyperplasia (12). In addition, tachykinins stimulate the proliferation of fibroblasts, epithelial, endothelial and airway smooth muscle cells. Further, SP stimulates the chemotaxis of lymphocytes, monocytes, neutrophils, eosinophils and is able to induce the release of eosinophilic cationic protein (ECP) and reactive oxygen species in human eosinophils (13,14). Most strikingly, eosinophils are a source of SP itself, underlining their participation in neuronal reflex circuits being able to stimulate nerves (15). Interestingly, increased baseline levels of SP have been found in upper and lower airways of asthmatics compared with healthy subjects which further increased after allergen challenge (16), suggesting that SP may also account for nasobronchial interaction mechanisms, which represents an important feature for bronchial asthma (17). doi:10.1594/eaacinet2007/EO/4-300307 http://www.eaaci.net NEUROMEDIATORS IN BRONCHIAL ASTHMA In addition to tachykinins, neurotrophins originally defined as neuronal growth factors, have been identified as important modulators for the functional plasticity of sensory and motor neurons and inflammatory signals regarding the bronchial nervous system. Animal data demonstrate that a number of pathomechanisms controlling allergic diseases are directly related to neurotrophin function, including the development of airway hyperresponsiveness (18,19). Increased levels of neurotrophins such as nerve growth factor (NGF) but also brain-derived neurotrophic factor (BDNF) have been found in the peripheral blood, bronchoalveolar lavage and nasal lavage fluid (NGF) of patients with allergic airway inflammation (20,21). However, it still remains unclear whether or not neurotrophins also contribute to nasobronchial interactions which represent an important feature for neuronal reflex circuits in bronchial asthma. Interestingly, neurotrophins not only directly modulate the functional activity of nerves- termed neuronal plasticity, but also have a direct functional effect on peripheral immune cells- counting for immunological plasticity. Again, this has been best investigated for the eosinophil granulocyte, which is not only a source of NGF and BDNF (23,24) but also target of BDNF and NGF as shown in functional studies of human eosinophils in vitro (23,25). CONCLUSION Patients with asthma feature both airway inflammation and airway hyperresponsiveness to non-specific stimuli which is partly neuronally controlled. Neurotrophins have been shown to be exceptional in supporting survival of neurons in the central and peripheral nervous system. In addition, tachykinins and neurotrophins modulate the functional activity of key effector cells of the inflammatory infiltrate, supporting the immunological plasticity. Further, inflammatory key effector cells such as eosinophil granulocytes, which are in close vicinity of airway nerves produce tachykinins and neurotrophins orchestrating neuronal feedback mechanisms. This bidirectional interaction of neuromediators between nerves and key effector cells bridges the gap between neuronal and immunological plasticity (Fig. 1). Therefore, bronchial asthma presents a complex disease in which nerves and their mediators are of major doi:10.1594/eaacinet2007/EO/4-300307 http://www.eaaci.net importance closing the loop for neuroimmune interactions, presenting promising therapeutic targets for the future. Figure 1 Concept for the role of neuromediators including tachykinins and neurotrophins bridging the gap between neuronal and immunological plasticity in bronchial asthma. NANC= Non-adrenergic noncholingeric, SP= Substance P; NKA= Neurokinin-A; BDNF= Brain-derived neurotrophic factor, NGF= Nerve growth factor, NT-3= Neurotrophin-3. References (1) (2) (3) (4) Walsh GM. Targeting airway inflammation: novel therapies for the treatment of asthma. Curr Med Chem 2006; 13(25):3105-11. Barnes PJ. Overview of neural mechanisms in asthma. Pulm Pharmacol 1995; 8(4-5):151-9. Minette PA, Lammers JW, Dixon CM, McCusker MT, Barnes PJ. A muscarinic agonist inhibits reflex bronchoconstriction in normal but not in asthmatic subjects. J Appl Physiol 1989; 67(6):2461-5. van dV, V, Hulsmann AR. Autonomic innervation of human airways: structure, function, and pathophysiology in asthma. Neuroimmunomodulation 1999; 6(3):145-59. doi:10.1594/eaacinet2007/EO/4-300307 (5) (6) (7) (8) (9) (10) (11) (12) (13) (14) (15) (16) (17) (18) (19) (20) (21) (22) (23) (24) (25) http://www.eaaci.net Sheppard D, Epstein J, Holtzman MJ, Nadel JA, Boushey HA. Effect of route of atropine delivery on bronchospasm from cold air and methacholine. J Appl Physiol 1983; 54(1):130-3. Aquilina AT, Hall WJ, Douglas RG, Jr., Utell MJ. Airway reactivity in subjects with viral upper respiratory tract infections: the effects of exercise and cold air. Am Rev Respir Dis 1980; 122(1):3-10. Costello RW, Schofield BH, Kephart GM, Gleich GJ, Jacoby DB, Fryer AD. Localization of eosinophils to airway nerves and effect on neuronal M2 muscarinic receptor function. Am J Physiol 1997; 273(1 Pt 1):L93-103. Fryer AD, Stein LH, Nie Z, Curtis DE, Evans CM, Hodgson ST et al. Neuronal eotaxin and the effects of CCR3 antagonist on airway hyperreactivity and M2 receptor dysfunction. J Clin Invest 2006; 116(1):228-36. Martling CR, Theodorsson-Norheim E, Lundberg JM. Occurrence and effects of multiple tachykinins; substance P, neurokinin A and neuropeptide K in human lower airways. Life Sci 1987; 40(16):1633-43. Joos GF, De Swert KO, Schelfhout V, Pauwels RA. The role of neural inflammation in asthma and chronic obstructive pulmonary disease. Ann N Y Acad Sci 2003; 992:218-30. de Vries A, Engels F, Henricks PA, Leusink-Muis T, McGregor GP, Braun A et al. Airway hyperresponsiveness in allergic asthma in guinea-pigs is mediated by nerve growth factor via the induction of substance P: a potential role for trkA. Clin Exp Allergy 2006; 36(9):1192-200. De Swert KO, Tournoy KG, Joos GF, Pauwels RA. The role of the tachykinin NK1 receptor in airway changes in a mouse model of allergic asthma. J Allergy Clin Immunol 2004; 113(6):1093-9. Wiedermann FJ, Kahler CM, Reinisch N, Wiedermann CJ. Induction of normal human eosinophil migration in vitro by substance P. Acta Haematol 1993; 89(4):213-5. Iwamoto I, Nakagawa N, Yamazaki H, Kimura A, Tomioka H, Yoshida S. Mechanism for substance P-induced activation of human neutrophils and eosinophils. Regul Pept 1993; 46(1-2):228-30. Metwali A, Blum AM, Ferraris L, Klein JS, Fiocchi C, Weinstock JV. Eosinophils within the healthy or inflamed human intestine produce substance P and vasoactive intestinal peptide. J Neuroimmunol 1994; 52(1):69-78. Nieber K, Baumgarten CR, Rathsack R, Furkert J, Oehme P, Kunkel G. Substance P and betaendorphin-like immunoreactivity in lavage fluids of subjects with and without allergic asthma. J Allergy Clin Immunol 1992; 90(4 Pt 1):646-52.. Braunstahl GJ. The unified immune system: Respiratory tract-nasobronchial interaction mechanisms in allergic airway disease. J Allergy Clin Immunol 2005; 115(1):142-8. Path G, Braun A, Meents N, Kerzel S, Quarcoo D, Raap U et al. Augmentation of allergic early-phase reaction by nerve growth factor. Am J Respir Crit Care Med 2002; 166(6):81826. Kerzel S, Path G, Nockher WA, Quarcoo D, Raap U, Groneberg DA et al. Pan-neurotrophin receptor p75 contributes to neuronal hyperreactivity and airway inflammation in a murine model of experimental asthma. Am J Respir Cell Mol Biol 2003; 28(2):170-8. Virchow JC, Julius P, Lommatzsch M, Luttmann W, Renz H, Braun A. Neurotrophins are increased in bronchoalveolar lavage fluid after segmental allergen provocation. Am J Respir Crit Care Med 1998; 158(6):2002-5. Sanico AM, Stanisz AM, Gleeson TD, Bora S, Proud D, Bienenstock J et al. Nerve growth factor expression and release in allergic inflammatory disease of the upper airways. Am J Respir Crit Care Med 2000; 161(5):1631-5. Bonini S, Lambiase A, Bonini S, Angelucci F, Magrini L, Manni L et al. Circulating nerve growth factor levels are increased in humans with allergic diseases and asthma. Proc Natl Acad Sci U S A 1996; 93(20):10955-60. Raap U, Goltz C, Deneka N, Bruder M, Renz H, Kapp A et al. Brain-derived neurotrophic factor is increased in atopic dermatitis and modulates eosinophil functions compared with that seen in nonatopic subjects. J Allergy Clin Immunol 2005; 115(6):1268-75. Solomon A, Aloe L, Pe'er J, Frucht-Pery J, Bonini S, Bonini S et al. Nerve growth factor is preformed in and activates human peripheral blood eosinophils. J Allergy Clin Immunol 1998; 102(3):454-60. Nassenstein C, Braun A, Erpenbeck VJ, Lommatzsch M, Schmidt S, Krug N et al. The neurotrophins nerve growth factor, brain-derived neurotrophic factor, neurotrophin-3, and neurotrophin-4 are survival and activation factors for eosinophils in patients with allergic bronchial asthma. J Exp Med 2003; 198(3):455-67.