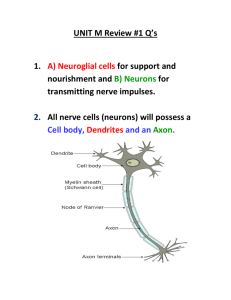
Neurons - University Psychiatry
advertisement

CHAPTER
1
Structure and Function of Neurons
Varieties of neurons
General structure
Structure
of unique neurons
Internal operations
Subcellular
and the functioning
of a neuron
organelles
Protei n synthesis
Neuronal transport:
shipping
and receiving molecules
and organelles throughout
the neuron
Summary
prise tens of billions of neurons, each linked to thousands of other neurons. Thus,
are has
the cells
of chemical
communication
in the
brain.asHuman
brains
combrain
trillions
of specialized
connections
known
synapses.
Neurons
have many sizes, lengths, and shapes, which determine their functions. Localization within
the brain also determines function. When neurons malfunction, behavioral symptoms may
occur. When drugs alter neuronal function, behavioral symptoms may be relieved, worsened, or produced. Thus, this chapter briefly describes the structure and function of normal
neurons as a basis for understanding psychiatric disorders and their treatments.
Neurons
the
Varieties of neurons
General
structure
Although this textbook will often portray neurons with a generic structure (such as that
shown in Figure l-IA and B), the truth is that many neurons have unique structures (see
Figures 1-2 through 1-8). All neurons have a cell body, known as the soma, and are set
up structurally to receive information from other neurons through dendrites, sometimes
via spines on the dendrites, and often through an elaborately branching "tree" of dendrites
(Figure 1-1A and B). Neurons are also set up structurally to send information to other
neurons via an axon, which forms presynaptic terminals as the axon passes by - "en passant"
(Figure l-lA) - or as it ends (in presynaptic axon terminals) (Figure l-IA).
Structure
of unique neurons
Many neurons in the central nervous system have unique structures. For example, each
pyramidal cell has a cell body shaped like a triangular pyramid (Figure 1-2A is a somewhat
Structure and Function of Neurons
I
1
dendrites
dendritic spines
cell body (soma)
dendritic
tree
/'
axon
oo )
presynaptic
axon terminals
en passant
presynaptic axon
terminals
general structure
of the neuron
A
B
another general structure
of the neuron
FIGURE l-IA and B Generic structure of neuron. This is an artist's conception of the generic structure of a
neuron. All neurons have a cell body known as the soma, which is the command center of the nerve and contains
the nucleus of the cell. All neurons are also set up structurally to both send and receive information. Neurons
send information via an axon, which forms presynaptic terminals as it passes by (en passant) or as it ends (A).
Neurons receive information from other neurons through dendrites, sometimes via spines on the dendrites, and
often through an elaborately branching tree of dendrites (B). Although all neurons share these properties, they
can have unique structures that, in turn, dictate specialized functions.
realistic depiction and 1- 2B is an icon of a pyramidal cell); each also has an extensively
branched spiny apical dendrite and shorter basal dendrites (Figure 1-2B) as well as a single axon emerging from the basal pole of the cell body. Pyramidal neurons are discussed
extensively in this textbook because they make up most of the neurons in the functionally
important prefrontal cortex as well as elsewhere in the cerebral cortex. Several other neurons are named for the shape of their dendritic tree. For example, basket cells are so named
because they have widely ramified dendritic trees that look rather like baskets (Figure 1-3A
is a somewhat realistic depiction and 1- 3B is an icon of a basket cell). Basket cells function
as interneurons in the cortex, and the wide horizontal spread of their axons can make many
local inhibitory contacts with the soma of other cortical neurons. Double bouquet cells are
also inhibitory interneurons in the cortex and have a very interesting vertical bitufted appearance, almost like two bouquets of flowers (Figure 1-4A is a somewhat realistic depiction and
1-4B is an icon of a double bouquet cell). Each double bouquet cell has a tight bundle of
axons that is also vertically oriented, with varicose collaterals that innervate the dendrites of
other cortical neurons, including other double bouquet cells, and supply inhibitory input to
those neurons. Spiny neurons, not surprisingly, have spiny-looking dendrites (Figure I-SA
2 I
Essential Psychopharmacology
motor cortex
pyramidal cell
body (soma)
""
basal dendrites
recurrent collateral (axon)
axon /
axon /
_ presynaptic axon
terminal
_ presynaptic axon
terminal
A
realistic pyramidal cell
B
icon of pyramidal cell
FIGURE 1-2A and B Pyramidal cells. Pyramidal cells (depicted somewhat realistically in A and iconically in B)
have a cell body shaped like a triangular pyramid, an extensively branched spiny apical dendrite, shorter basal
dendrites, and a single axon emerging from the basal pole of the cell body. The majority of the neurons in the
cerebral cortex, particularly in the prefrontal cortex, are pyramidal neurons.
is a somewhat realistic depiction and 1-5B is an icon of a spiny neuron). Spiny neurons are
located in the striatum in large numbers and have a highly ramified dendritic arborization
that radiates in all directions and, of course, is densely covered with spines, which receive
input from cortex, thalamus, and substantia nigra. Spiny neurons have long axons that
either leave the striatum or circle back as recurrent collaterals to innervate neighboring spiny
neurons. Finally, Purkinje cells from the cerebellum form a unique dendritic tree that, in
fact, looks very much like a real tree (Figure 1-6). This dendritic tree is extensively branched and fans out from an apical position, with a single axon emerging from the basal pole
of the cell.
At least one type of neuron is named for its unique axonal structure: the chandelier
neuron (Figure 1-7A is a somewhat realistic depiction and 1-7B is an icon of a chandelier
neuron). The axons of this cell look like an old-fashioned chandelier, with odd-appearing
axon terminals shaped like vertically oriented cartridges, each consisting of a series of
axonal swellings linked by thin connecting pieces. Chandelier neurons are yet another type
of inhibitory interneuron in the cortex, where the characteristic "chandelier" endings of their
axons have a specific function and location - namely, to serve as inhibitory contacts close to
the initial segment ofaxons of pyramidal cells. Thus, chandelier neurons terminate in what
Structure and Function of Neurons
I
3
'\/ terminals
presynapticaxon
A
realistic basket cell
B
icon of basket cell
FIGURE 1-3A and B Basket neurons. Basket neurons are named for their widely ramified dendritic trees, which
resemble baskets (depicted somewhat realistically in A and iconically in B). They are cortical interneurons with
axons that spread horizontally to make many inhibitory contacts with the soma of other neurons.
--... presynaptic axon
terminal
......cell body
bouquet shape --...
of dendritic tree
;! --... bouquet shape
of dendritic tree
"
.
presynaptic axon
terminal
A
realistic double bouquet cell
B
icon of double bouquet cell
FIGURE 1-4A and B Double bouquet cells. Double bouquet cells are so called because of their vertical bitufted
appearance, which resembles two bouquets of flowers (depicted somewhat realistically in A and iconically in B).
Like basket neurons, double bouquet cells are inhibitory interneurons in the cortex. They have a tight bundle of
axons that is oriented vertically, with varicose collaterals that innervate the dendrites of other cortical neurons,
including other double bouquet cells.
4 I
Essential Psychopharmacology
spiny neuron
cell body
-J
/'
spiny dendrites
presynaptic
axon
terminal
/'
presynaptic
axon
terminal
A
realistic spiny neuron
B
icon of spiny neuron
FIGURE I-SA and B Spiny neurons. The dendrites of spiny neurons radiate in all directions and are densely
covered with spines (depicted somewhat realistically in A and iconically in B). Spiny neurons are located in the
striatum in large numbers and receive input from cortex, thalamus, and substantia nigra. The axons of spiny
neurons are long and either leave the striatum or circle back as recurrent collaterals to innervate neighboring
spiny neurons,
FIGURE 1-6 Purkinje cells. Purkinje
cells from the cerebellum have
extensively branched dendritic trees
fanning out from an apical position,
with a single axon emerging from the
basal poll of the cell,
'2
k"
/
unique Pur InJe
/'
dendritic tree
cell body
presynaptic
/axon terminal
Purkinje cell
is called an axoaxonic synapse. Since the initial segment of a pyramidal cell's axon is the
most influential location in determining whether that axon will fire or not, the chandelier
neuron can potentially provide the most powerful inhibitory input to a pyramidal neuron,
possibly even being able to completely shut down a pyramidal cell's firing. Many chandelier
Structure and Function of Neurons
I
5
chandelier
axon
/terminalS
axon
realistic chandelier neuron
A
B
icon of chandelier neuron
FIGURE 1-7A and B Chandelier neurons. The chandelier neuron is named for its unique axonal structure
(depicted somewhat realistically in A and iconically in B). The axons resemble an old-fashioned chandelier with
axon terminals shaped like vertically oriented cartridges, each consisting of a series of axonal swellings linked
together by thin connecting pieces. Like basket neurons and double bouquet cells, chandelier neurons are
inhibitory interneurons in the cortex. The "chandelier" endings of their axons come into close contact with the
initial segments of pyramidal cell axons, forming what is called an axoaxonic synapse. The chandelier neuron can
potentially provide powerful inhibitory input to a pyramidal neuron via this synapse, possibly even completely
shutting down a pyramidal cell's firing. Many chandelier neurons provide input to a given pyramidal cell, and
each chandelier neuron can provide input to several pyramidal cells.
neurons provide input to a given pyramidal cell, and each chandelier neuron can provide
input to several pyramidal cells.
Internal operations and the functioning
Subcellular
of a neuron
organelles
In order to do its duties, the neuron contains various internal working parts that have
specialized functions, from subcellular organelles and protein synthetic machinery to internal superhighways for transport of these materials into dendrites and axons on specialized
molecular "motors." Specific neuronal functions are associated with each anatomical zone of
a neuron (Figure 1-8). For example, the soma and dendrites together form the somatodendritic zone, which has the function of "reception." Neurons receive a wide variety of signals,
sometimes simultaneously and sometimes sequentially, from other neurons, environment,
chemicals, hormones, light, drugs, and so on. In addition to receiving this mass of incoming
information, the somatic zone also serves as a "chemical integrator" of it all. It does this
6 I
Essential Psychopharmacology
General Structure and Function of the Neuron
Structure
Function
somatodendritic
zone
somatic
zone
-zone
elements
synapse
initial segment
~
zone
output
propagation
signalelectrical
encoding
I
~
~
I
presynaptic
presynaptic
presynaptic
o0
signal
signal
I
I
I
n
zone
D
axonal
reception
integration
chemical
encoding
zone
axon hillock
I
FIGURE 1-8 Anatomic zones of neurons. The different anatomic zones of neurons are associated with specific
functions, as shown here. The soma and dendrites form the somatodendritic zone, which has the function of
receiving a wide variety of signals from other neurons. The somatic zone also serves as a chemical integrator of
incoming information: incoming signals from postsynaptic dendrites are decoded by the genome (located in the
cell nucleus in the soma), which then encodes chemical signals destined for either internal or external
cornmunication. The initial segment of the axon, the axon hillock, serves as an electrical integrator, controlling
whether or not the neuron will fire in response to incoming electrical information. The axon propagates these
signals, with electrical signals traveling along the membrane of the axon and chemical signals traveling within its
internal structural matrix. The presynaptic zone at the end of the axon contains unique structures that convert
chemical and electrical signals into signal output.
by first generating cascades of incoming chemical signals from its postsynaptic dendrites,
which speak directly with its genome, located in the cell nucleus in the soma (Figure 1-8).
These incoming volleys of chemical information are then decoded and read by the genome,
after which the genome adds its own reaction to this information by encoding chemical
Structure and Function of Neurons
I 7
nucleus
~
anterograde
motor
Golgi apparatus
retrograde
motor
RNA
polysomes
~
mitochondrion
~
~
rough endoplasmic reticulum
microtubule
peptide/
'@
secretory granule
protein
neurotransmitter
'@
synaptic vesicle
Iysosomes
-- -
-
neurofilaments
cytoskeleton
pre-/postsynaptic density
retrograde
vesicle
FIGURE 1-9 Neuronal components. Depicted here are many neuronal components manufactured by the cell
nucleus, which contains the neuron's DNA. These components are located in specific locations within the neuron
and have specific functions.
signals destined either for internal communication within its own neuronal boundaries or
for external communication via its neuronal connections.
Another anatomical zone is that of the axon hillock, also called the axon's initial segment
(Figure 1-8). Its job is to serve as an "electrical integrator" of all the incoming electrical
information and decide whether or not to "fire" the neuron. Directly connected to the
axon hillock is the axon itself, which propagates electrical signals along its membrane and
chemical signals within its internal structural matrix. At the end of the axon is a specialized
zone with unique structures that allow it to convert the chemical and electrical signals
arriving there into signal output to the next neuron.
How does all of this happen? It is done by orchestrating many specialized neuronal
instruments to work together in amazing functional harmony - at least when things are
working normally. Many components of a functioning neuron are shown in Figure 1-9.
A representation of where these components are localized within the neuron is shown in
Figure 1-10. These specialized neuronal instruments are put into action in the remaining
figures of this chapter (Figures 1-11 through 1-20). The specific roles thatthese specialized
neuronal instruments play in neuronal functioning as shown in these figures are eXplained
briefly here.
As already mentioned, the cell nucleus, containing the neuron's DNA, is located in the
neuron's soma and is responsible for manufacturing essentially all the components shown
8
I
Essential Psychopharmacology
Localization
of Subcellular
Organelles
synaptic vesicles
presynaptic
density
FIGURE 1-10 Localization of neuronal components. The function of each neuronal component is unique; in
addition, each component is distributed differently throughout the neuron, as shown here. Thus, different parts of
the neuron are associated with different functions. For example, DNA transcription occurs only in the soma, while
protein synthesis, which involves polysomes and endoplasmic reticulum, occurs both in the soma and in
dendrites.
Structure and Function of Neurons
9
Synthesis
of a Cytoplasmic/Peripheral
Protein: Ready for Transport
".
free
A./ polysomes
Udd
cP,
\' <
'Ji{fT
W
nUcl~eus> ...
~~n~s
~R~~
;1•
~
p~ripheral
protem
FIGURE 1-11 Protein synthesis. Most of the structural and regulatory molecules of a neuron are proteins. When
DNA is transcribed into RNA, it is read by one of two types of ribosomes: free polysomes, which are not
membrane bound, or rough endoplasmic reticula, which are membrane bound. Proteins are then synthesized
on/within the ribosomes. Peripheral proteins, which are soluble and live in the cytoplasm, are synthesized on free
polysomes and transported directly into the dendrites and axons.
in Figures 1-9 and 1-10. As can be seen from Figure 1-10, these components have specific
locations within the neuron's specialized structure; therefore some functions occur in one
part of the neuron but not another. For example, all the nuclear DNA is transcribed in
the soma but all protein synthesis does not occur there, because the synthetic machinery
of polysomes and endoplasmic reticulum exists in dendrites as well as the soma but not
to any great extent in axons (Figure 1-10). The vital function of transport occurs in both
axons and dendrites, but there are more microtubules for transport in dendrites and more
neurofilaments for transport in axons (Figure 1-10). Cytoskeletal support proteins exist
along the membranes of the entire neuron, but postsynaptic density proteins exist only in
dendrites and soma membranes and at the beginning and end ofaxons, whereas presynaptic
density proteins exist only in axon terminals (Figure 1-10).
10 I
Essential Psychopharmacology
FIGURE 1-12 Peptide synthesis.
Synthesis of Integral Membrane and
Secretory Proteins and Peptides:
Packaged for Transport
Integral or secretory proteins, or
peptides, are proteins that are inserted
into a membrane. They are produced
when mRNA is read by the rough
endoplasmic reticulum, which
"'J
synthesizes these proteins and packages
them into vesicles to be sent to the
nucleus
Golgi apparatus. The proteins are then
""""
genes
I,.!~ 666
t(fDNA/
'-rv
mRNA'"
Ir-J~~
\
modified within the Golgi apparatus and
packaged into secretory vesicles ready
for transport.
endoplasmic
reticulum
rough
.
3
~'2
(;6
@\
~
@
Protein synthesis
Few neuronal functions are more important than the synthesis of proteins, which are produced as the result of gene activation. Because most of the important structural and regulatory molecules of a neuron are proteins, they functionally carry out orders from the
genome. For example, proteins become the building blocks when the genome orders a new
synapse to be made; proteins are the receptors and enzymes of the neuron; proteins can
activate messengers or synthesize anything the neuron needs. Thus, it is no surprise that the
neuron is organized so that high priority can be given to making and transporting various
proteins.
Proteins are synthesized on a subcellular organelle known as a ribosome. When DNA
is transcribed into RNA, the RNA can be read by either of two types of ribosomes in order
for proteins to be synthesized. One type are called free polysomes, because they are not
membrane-bound. The other type are membrane bound and are called rough endoplasmic
reticulum, or "Nissl substance." Protein synthesis occurs predominantly in the soma (Figures
1-11 and 1-12). Proteins that are soluble, and thus live in the cytoplasm, are synthesized on
Structure and Function of Neurons
I
11
secretory
protein
FIGURE 1-13 Dendrite protein synthesis. Mostproteinsynthesisoccurs in the soma; however,some protein
synthesisoccurs in dendrites.mRNAis somehow made accessible, perhapsvia microtubules,to free polysomes
and roughendoplasmicreticulalocatednear dendriticspines, whichthen synthesizeproteinslocally.
free polysomes and then transported directly into dendrites and axons, wherever they are
needed (Figure 1-11). These are called peripheral proteins. Proteins that are destined for
insertion into a membrane, called integral or secretory proteins or peptides, are synthesized
within the rough endoplasmic reticulum, packaged there into vesicles, and shipped to the
Colgi apparatus, which modifies and molecularly "decorates" these proteins; finally, they
exit the Colgi apparatus in secretory vesicles, ready for transport (Figure 1-12).
Some protein synthesis occurs in dendrites (Figure 1-13). Presumably these proteins are
necessary for implementing those specialized functions unique to dendrites, such as receiving
information, forming postsynaptic signal reception and signal transduction machinery, and
the like. Polysomes are located in dendrites, often close to dendritic spines. RNA formed
in the soma is somehow accessible to these polysomes in the dendrites, so that proteins can
be synthesized locally where they would be ready for action immediately upon synthesis, as
they would not need transport into the dendrite.
Neuronal
throughout
transport:
shipping and receiving molecules
and organelles
the neuron
Much of the neuron functions like a busy depot. Following the manufacture of protein and
organelles, these components must be packaged and shipped. Some must be dispatched
with the speed of an "overnight" delivery system (fast transport), whereas others are sent
with the deliberation of "snail mail" (slow transport). The various transport systems up and
down axons and dendrites form a type of neuronal infrastructure of roads and bridges to get
every component where it must go and when it must get there. For example, cytoplasmic
proteins are sent into both axons and dendrites by a slow transport system (Figure 1-14).
This system is really slow, moving only about 2 mm a day, or 50 to 100 !Jm an hour. Slow
transport motors (indicated by a tortoise carrying the soluble proteins in Figure 1-14) crawl
12
i
EssentialPsychopharmacology
Slow Transport of Cytoplasmic
Proteins
cytoplasmic
protein
slow transport
motor
cytoskeleton
FIGURE 1-14 Slow transport of proteins. Once proteins and organelles have been made, they must be
transported to their ultimate destination. This can occur via one of two delivery systems: slow transport or fast
transport. Cytoplasmic proteins are sent via slow transport motors (depicted here as tortoises) that crawl along
the cytoskeleton at a rate of 2 mm per day, or 50 to 100 ILm an hour.
along the cytoskeleton and slowly yet inexorably deliver these proteins to both axonal and
cytoplasmic destinations. Interestingly, the infrastructure system itself is also transported
via this slow transport system (Figure 1-15). Thus, microtubules are transported slowly into
dendrites and axons and neurofilaments are transported into axons (Figure 1-15) to form
the very highways upon which other components are rapidly transported through the fast
transport systems, which are shown in Figures 1-16 though 1-20.
Structure and Function of Neurons
./
I 13
FIGURE 1-15 Slow
Slow Transport of Microtubules
and Cytoskeleton
transport of microtubules
and neurofilaments. Slow
transport is also the
deliverysystem used for
movingthe organelles
involvedin fast transport.
Thus, microtubulesare
deliveredto dendritesand
axons and neurofilaments
are deliveredto axonsvia
slowtransport.
slowtransportmotor
fast transportmotor
cytoplasmic
protein
cytoplasmic
enzyme
microtubule
cytoskeleton
neurofilament
Many neuronal materials are passengers that ride on fast transport systems with fast
transport motors, which are shown as hares in Figure 1-17. Such passengers include mitochondria, synaptic vesicles containing neurotransmitters, secretory vesicles containing secretory proteins, and all sorts of other proteins, from receptors to enzymes to ion channels
to transport pumps and many more. Transport of these materials allows supplies depleted
during the normal conduct of neuronal business by dendrites and axons to be replenished. A fast transport system (indicated by "hares" in Figures 1-17 through 1-20) carries
membrane-bound secretory vesicles full of secretory proteins at about 200 mm per day, but
only from the soma to the axon terminal, a direction known as "anterograde" and designated
in Figure 1-17 as "southbound lanes." There is also transport in the opposite direction,
known as "retrograde" and designated in Figure 1-18 as "northbound lanes." However,
14
EssentialPsychopharmacology
Types of Materials for Fast Transport
mitochondria
receptors
synaptic vesicles
enzymes
.........
growth factor
..
anterograde
secretory
vesicles
retrograde
secretory
vesicles
::::~
ion channels
..
<q
o
neurotransmitters
~neurotransmitter
peptide
~
oldgrowth
mitochondrion
factor
reuptake pumps
(neurotransmitter
transporters)
peptides
FIGURE 1-16 Materials for fast transport. Passengersof fast transport systems includemitochondria,synaptic
vesiclescontainingneurotransmitters,secretoryvesiclescontainingsecretoryproteins,receptors,enzymes,ion
channels, reuptake pumps, and other proteins.
retrograde transport is about half as fast and includes the return of used and discarded
proteins and organelles from the axon terminal, which are shipped up to the soma for
destruction in lysosomes. Also, the retrograde system takes up growth factors and viruses
from the synapse and sends them up to the soma, where they can signal the genome chemically (Figure 1-18).
Another fast transport system carries the machinery for synthesizing, metabolizing, and
utilizing neurotransmitters. In the case oflow-molecular-weight
neurotransmitters such as
monoamines, this includes all of their synthetic machinery, since these neurotransmitters
are not only manufactured in the soma and shipped to the axon terminal but are also made
locally in the axon terminal from synthetic enzymes shipped there (Figure 1-19). This is
important, because the rate of utilization of these neurotransmitters can be greater than
the rate at which they can be shipped all the way from the soma, even on a "fast" transport
system. Neurotransmitter is thus packaged and stored in the presynaptic neuron in vesicles,
like a loaded gun, ready to fire. Since a reuptake pump (monoamine transporter), which can
recapture released monoamines, is present on the presynaptic neuron, monoamines used in
one neurotransmission can be captured for reuse in a subsequent neurotransmission. This
is in contrast to the way in which neuropeptides function in neurotransmission (Figure
1-20). That is, higher-molecular-weight
peptides are synthesized only in the soma and are
not taken back up into the presynaptic neuron by a reuptake pump. Fortunately, peptide
Structureand Functionof Neurons I 15
,!
200
tr~
Limit
'" "
mitochondrion
synaptic vesicle
anterograde
receptor
enzyme
slow
transport vesicle
motor
tast
transport
motor
microtubule
~
.c.", •••••
~
~
'000""'"' "'~ I
0
•
bm
"
0
H
I
ion channel
~
cytoskeleton
FIGURE 1-17 Fast anterograde transport. Shown here is delivery of various neuronal components to their
axonal destinations via fast transport. Membrane-bound secretory vesicles full of secretory proteins are
transported at a rate of 200 mm per day from the soma to the axon terminal in a direction known as anterograde
(depicted here as southbound lanes).
16
I
Essential Psychopharmacology
w
fast transport motor
@
lysosome
old mitochondrion
~
old synaptic vesicle
Northbound Lanes
Speed
Limit
100
mm/day
o
~:-:~.
retrograde vesicle
growth factor
microtubule
cytoskeleton
FIGURE 1-18 Fast retrograde transport.
Fast transport also occurs in the opposite direction at 100 mm per day;
this is known as retrograde transport (designated as northbound lanes here). With retrograde transport, used and
discarded proteins and organelles are brought from the axon terminal to the soma, where they are destroyed by
Iysosomes. In addition, growth factors and viruses from the synapse are sent to the soma, where they can signal
the genome chemically.
Structure and Function of Neurons
17
•
serotonin
amino acid
LIMIT
0
100
SPEED
mm/day
CD
(~I~
m~
monoamine
vesicle
oxidasesynaptic
decarboxylase
last transport motor
W
I
L1
fi
hydroxylase
tryptophan
~
FIGURE 1-19 Fast transpDrt: low-molecular-weight neurotransmitter machinery. Another fast transport system
carries the machinery for synthesizing, metabolizing, and utilizing neurotransmitters.
Because the synthetic
enzymes involved in manufacturing low-molecular-weight neurotransmitters such as monoamines are transported
to the axon terminal, these neurotransmitters can be made both in the soma and locally in the axon terminal. In
addition, reuptake pumps can recapture released neurotransmitters for reuse in subsequent neurotransmission.
This is important because the rate of utilization of these neurotransmitters can be greater than the rate at which
they can be shipped from the soma.
18 I
Essential Psychopharmacology
D
.D~Ad-,mRN\A,,{;f
0)\
P1"" ~':-f-<'?[
@primary
(
peptide
pmpm-
gene
mRNA
., !~
inactive
peptide
prepropeptide
propeptide
endoplasmic
catabolic
converting
metabolite
core
vesicle
reticulum
enzyme
peptidase
fast transport motor
large dense,
~
~~ a:::::>
~
100
FIGURE 1-20 Fast transport:
'It'.,.
prepropeptide /
larger neuropeptide
0
/
"
c:::::::>
i!:.
I
machinery. Unlike low-molecular-weight
neurotransmitters,
larger neuropeptides are synthesized only in the soma and are not taken back up into the presynaptic neuron by a
reuptake pump. However, peptide neurotransmitters are generally released more slowly, allowing transport of
these neurotransmitters from the soma in larger dense-core vesicles to keep up with demand.
Structure and Function of Neurons
I
19
neurotransmitters are generally released more slowly, so that transport of these neurotransmitters from the soma in larger dense-core vesicles can keep up with demand (Figure 1-20).
Summary
This chapter has described the structure and function of various types of neurons. Although
all neurons share some structural similarities, there are many unique aspects to some neurons, including the shapes of their somas, dendritic trees, and axons. This chapter has also
reviewed how the various components of a neuron work together to carry out specialized
functions, such as synthesis of important neuronal proteins and transport of proteins and
other vital supplies throughout the neuron. An understanding of the structure and function
of normal neurons can provide a good background for grasping what goes wrong with neurons in various psychiatric disorders and how drugs affect neurons to treat various psychiatric
disorders.
20
Essential Psychopharmacology