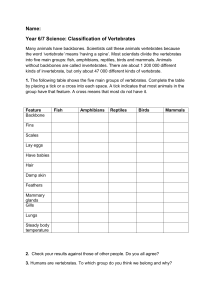
the digestive system of vertebrates
advertisement

THE DIGESTIVE SYSTEM OF VERTEBRATES 1University Esther J. Finegan, PhD1 and C. Edward Stevens, PhD, DVM2 of Guelph, Ontario, N1G 2W1 Canada; 2Department of Molecular Biomedical Sciences, College of Veterinary Medicine, North Carolina State University, Raleigh, NC, 27606 USA. Evolution of the digestive system Introduction: This section summarizes the adaptations of the digestive system to the habitat, diet and other physiological characteristics of vertebrates and speculates on how these may have evolved. Speculations on the digestive system of early vertebrates are based mostly on comparisons of their skeletal remains, habitat, and environment with those of present-day species. Although our discussion has been limited to vertebrates, they represent less than 5% of the species in the Animal Kingdom. As Barnes (1987) pointed out, a taxonomist less biased than Man might have divided these animals into arthropods (over 75% of the species) and non-arthropods. Separation of the Animal kingdom into vertebrates and invertebrates is based on one characteristic of a single subphylum, and many characteristics of the vertebrate digestive system are found among the invertebrates. Therefore, a brief discussion of the invertebrate digestive systems provides some necessary perspectives. Digestive System - Invertebrates: The structural and functional variations in the invertebrate digestive system are discussed by Barnard and Prosser (1973), Barnes (1974; 1987), Wigglesworth (1984), and Vonk and Western (1984). Although it is convenient to use the terms primitive, advanced, lower, higher, and specialized in discussions of phylogenetic relations, this tends to create the erroneous impression that evolution progressed toward an ideal goal. Lower and higher generally refer to the level at which species have stemmed from a main line of evolution. Primitive species are those believed to possess many or the greatest number of characteristics of the ancestral stock within a particular group of animals. Advanced species are those that have changed considerably as a result of different environments or modes of existence. The term specialized refers to characteristics that are especially adapted to a particulate ecological niche. However, as Barnes (1974) points out, the terms advanced and specialized should not be interpreted as more perfect or better, and some species with primitive characteristics are specialized in other respects. The general pattern of invertebrate evolution is illustrated in Figure 12.1. Protozoa (flagellates, sarcodinians, and ciliates), which are believed to have originated as singled-celled organisms in the Archeozoic oceans long before the first fossil records, have no digestive tract. Some presentday species absorb nutrients across their cell membrane, but many protozoa ingest food by phagocytosis at a specific site or various points on the cell membrane (Fig. 12.2). Ingested material is taken up by food vacuoles, which are passed through the cell with digestion of their contents, absorption of nutrients, and the eventual evacuation of waste products. Some of the digestive enzymes found in vertebrates have been isolated from protozoa (Vonk and Western 1984) and intervacuolar digestion can be accompanied by a similar cycle of acidification and alkalization in some species. Figure 12.1. Phylogeny of the Animal Kingdom as reflected by the views of L. Hyman. (From Barnes 1987). Figure 12.2. Formation of food vacuoles and digestion in a ciliated protozoa, such as Tetrahymena. (From Barnes 1987) A number of primitive metazoans, such as the Cnidaderian hydra, have a mouth and a blind gastrovascular cavity, which is lined with phagocytic, secretory, and ciliated, cells (Fig. 12.3). However, most free-living advanced invertebrates have a digestive tract that terminates in an anus. Movement of food and digesta is accomplished by cilia in some species, but this is aided by muscular activity in most of the more advanced invertebrates and cilia are absent in the nematodes and insects. The digestive tract of annelids, mollusks, and arthropods can be divided into a headgut, foregut, and an intestine. The headgut may be designed for a variety of functions, including filter-feeding, chewing, sucking, or piercing. Scorpions and mites regurgitate enzymes into their prey, and some echinoderms evert their stomach to engulf their prey. Figure 12.3. Body form (A) and body wall (B) of a hydra. (From Barnes 1987) The advanced invertebrates show an increasing dependence on extracellular digestion and the replacement of phagocytosis by absorption into the cells (Barrington 1962). Secretion of enzymes, absorption of nutrients, and food storage, which are carried out by multipurpose cells in the lower forms of invertebrates, become the properties of specialized cells. The intestinal ceca of free-living flatworms in Phylum Platyhelminthes, which are considered the most primitive bilateral animals, contain both secretory cells that release mucus and enzymes, and absorptive cells that lack food vacuoles and have microvilli on their lumen-facing membranes. Phylum Annelida is comprised of 8700 species of segmented worms, including the familiar earthworms. The digestive tract of lumbricid earthworms consists of a headgut (mouth, buccal cavity, and muscular pharynx), foregut (esophagus, crop, and gizzard), and intestine (Fig. 12.4). The anterior half of the intestine is the principal site of secretion and digestion, and the posterior half is the principal site of absorption. The intestinal absorptive cells of Arenicola marina, a burrowing annelid that ingests much sand along with organic matter, phagocytize food particles and transfer them to wandering amoebocytes in a manner similar to that of primitive metazoa (Barrington 1962). However, the midgut absorptive cells of most annelids lack food vacuoles and demonstrate microvilli or a brush border on their lumen-facing membranes. Figure 12.4. Anterior internal structures of the earthworm Lumbricus. (From Barnes 1987) Phylum Mollusca contains the clams, oysters, snails, slugs, squid, and octopods, and the largest number of species (80,000) of any phylum other than Arthropoda. The mollusks include carnivores, omnivores, scavengers, and parasites that inhabit marine, freshwater, and terrestrial environments. Digestion is at least partly extracellular in all mollusks, and enzymes may be secreted by salivary glands, esophageal pouches, portions of the stomach, intestinal glands, or a combination of these. A chitinous radula in the headgut of many mollusks grinds food into smaller particles (Fig. 12.5). Cilia in the stomach or style sac of some mollusks rotate its contents into a mucous mass (protostyle) and direct finer particles of food into digestive glands for digestion and absorption. The remainder is directed into the intestine for absorption of water and excretion of waste material. Figure 12.5. Lateral view of internal structures a generalized mollusk. (From Barnes 1987) Diverticula in the stomach of some mollusks contain phagocytic vacuolated cells (Barrington 1962). However, the absorptive cells in the digestive glands of others have microvilli and the digestive glands of the snail Helix secrete a variety of digestive enzymes but have little absorptive capacity. The digestive glands of some mollusks contain cells that can function for absorption, intracellular digestion, food storage, secretion, or excretion, and are often referred to as the hepatopancreas or the pancreas and liver in squid (Fig. 12.6). The squid and octopods have well developed salivary and digestive glands. The pancreas of squid produces aminopeptidase, dipeptidase, and lipase. The liver produces aminopeptidase, dipeptidase. and carboxypeptidase. Although cell vacuoles of the pancreas contain food particles, nutrients must be absorbed into the circulatory system to reach the liver cells. The pancreatic and liver ducts combine to form a common duct that can direct its flow into the stomach or the cecum, and a sphincter at the terminus of the liver duct prevents the backflow of lumen contents. Figure 12.6. Digestive tract of the squid Loglaga and octopus Octopus vulgaris. (From Barnes 1987) Phylum Arthropoda (horseshoe crabs, crustaceans, arachnids, and insects) includes over 75% of the animal species. Most crustaceans are filter feeders. Their foregut is often enlarged and lined with chitinous ridges to provide a triturating stomach. Their midgut contains glandular ceca that are modified into large digestive glands in some species and provide the principal source of extracellular digestive enzymes. Digestion in arthropods is principally extracellular. The "hepatic ceca" of horseshoe crabs consists of two large glands that function for both absorption and secretion of digestive enzymes. The "hepatopancreas" of the crustacean midgut consists of a pair of ceca and secretory ducts, which serve for absorption, secretion of digestive enzymes, and storage of glycogen, fat, and calcium (Fig. 12.7). Many advanced invertebrates produce emulsifying agents that serve functions similar to those of the vertebrate bile salts (Haslewood 1967). Figure 12.7. Digestive system of a crustacean crayfish. (From Barnes 1987) Van Weel (1974) concluded that the terms hepatopancreas, pancreas, and liver are inappropriate when applied to mollusks or crustaceans and they should be referred to simply as digestive glands. Bidder (1976) agreed and proposed the substitution of "digestive glands" and "digestive gland appendages" for the "liver" and "pancreas" of the cephalopods. However, Gibson and Barker (1979) concluded that the digestive glands of decapod crustaceans "are rightly and properly named the hepatopancreas”. The largest group of arthropods is the insects, which contain more than 750,000 species that have adapted to all types of habitats and are the most successful of all terrestrial animals. The digestive tract of most insects consists of a headgut, foregut (esophagus, crop, proventriculus, and gastric ceca), midgut, and hindgut (Fig. 12.8 and 12.9). Their headgut is designed for a variety of functions, including chewing, sucking, or piercing. Salivary glands are highly developed in many insects. They secrete mucus, enzymes, anticoagulants, agglutinins, venomous spreading agents or silk in various species. The crop serves for food storage and the proventriculus controls the passage of food into the midgut and contains teeth for the crushing or grinding of food in some species. The midgut is the principal site of digestion and absorption. Cells in the anterior midgut of many insects generate a thin sheath of peritrophic membrane, which protects it against physical damage but is permeable to nutrients. Malpighian tubules, which are analogous to the kidneys of vertebrates, release electrolytes, waste products, and water into the midgut. Figure 12.8. Schematic diagram of the digestive tract of an insect. (Modified from Wigglesworth 1962) Figure 12.9. Modifications of the gastrointestinal tract of insects. The foregut and hindgut are indicated by a red highlight. (From Wigglesworth 1962) The hindgut of insects aids in the recovery of electrolytes, water, and nitrogen (Fig. 12.10). The Malpighian tubules of the desert locust secrete Na+, K+, Cl-, ammonia-urate, proline and water, which are released into the intestinal contents at the juncture of the midgut and hindgut (Phillips et al. 1988). The rectal pad of their hindgut absorbs Na+ in cotransport with proline and in exchange for H+ or NH4+ (Fig. 12.11). Absorption of electrolytes and water from the rectum of cockroaches can produce extremely hypertonic solutions in the lumen (Wall 1971), and the hindgut is extremely voluminous in some herbivorous insects. Figure 12.10. Enterocirculation of water by the alimentary tract of insects. Letters indicate the midgut (a), Malpighian tubules (b), hindgut (c), and rectum (d). (From Wigglesworth 1962) Figure 12.11. Transport mechanisms in the apical and basolateral membranes of the locust rectal pad epithelium. Arrows through solid circles indicate carriermediated transport. Thick arrows indicate major ion pumps. Sodium is transported across the apical membrane in cotransport with amino acids and in exchange for intracellular H+ and the intercellular NH4+. (From Phillips et al. 1988) Many of the digestive enzymes of vertebrates have been demonstrated in various species of invertebrates (Vonk and Western 1984), and nutrients may be assimilated by similar mechanisms. For example, Na+ was absorbed electrogenically from the gut of the mollusk Alplysia californica (Gerencser 1988), and the rectal pad locusts demonstrated both Na+-dependent absorption of amino acids and Na+ - H+ exchange (Phillips et al. 1988). However, the electrogenic Cl- transport reported in the gut of the mollusk Alplysia californica (Gerencser 1988) and electrogenic exchange of two Na+ for one H+ described in the hepatopancreas brush border membranes of the freshwater prawn Macrobrachium rosenbergii and marine lobster Homarus americanus (Ahern et al. 1990) appear to be unique to invertebrates. Digestive System - Vertebrates: The chronology of terrestrial vertebrate evolution is illustrated in Figure 12.12. Amphibians are believed to have evolved from predacious lobe-finned fishes (Crossopteygii). Amniotic tetrapods appear to have evolved from amphibian anthrocosaurs, over 300 million years ago (mya). The reptiles evolved into the dinosaurs, which were the dominant terrestrial vertebrates from the mid-Triassic to the end of the Cretaceous period, and present-day chelonians, snakes, lizards and crocodilians. Birds are believed to be the modern-day descendants of carnivorous theropod dinosaurs (Gauthier 1986). Mammals appeared about 200 mya in the Late Triassic period of the Mesozoic Era (Lillegraven et al. 1979) and evolved into the Prototheria (monotremes) and, via the Pantotheria, into the Metatheria (marsupials) and Eutheria (placental mammals). The earliest Mesozoic mammals were small (20-30 g) carnivores that fed on invertebrates and other vertebrates (Crompton 1980). The Insectivora are regarded as their most direct modern descendants (Romer 1966). However, the major expansion and diversification of birds and mammals did not occur until after the end of Mesozoic (70 mya) and demise of the dinosaurs and most other vertebrates over 10 kg in body weight. Figure 12.12. Phylogenetic origins of various groups of vertebrates. A) Urodela, B) Lepospondylii, C) Apoda, D) Anura, E) Labyrinthodontia, F) Apisidospondylii, G) Chelonia, H) Anapsida, I) Cotylosauria, J) Eurapsida, K) Diapsida, L) Eosuchia, M) Squamata, N) Rhyncocephalia, O) Ornithischia, P)Thecodontia, Q) Synapsida, R) Parapsida, S) Pelycosauria, T) Pterosauria, U) Crocodilia, V) Aves, W) Saurischia, X) Prototheria, Y) Metatheria, Z) Pantotheria, AA) Therapsida, BB) Eutheria, CC) Ichthyosauria. (Modified from Torrey 1971) The digestive tract of vertebrates shows many features that are analogous to those of advanced invertebrates. Filter-feeders are found among fish (basking sharks, paddlefish), larval amphibians, birds (flamingos), and mammals (baleen whales). A beak is used for cutting, tearing, or crushing in the chelonians (turtles, terrapins, and tortoises) and birds. However, teeth are used for this purpose in most other vertebrates. Teeth are located in the jaws, other mouthparts, or the pharynx of fish, but confined to jaws of most vertebrates. Food can be ground to small particles by pharyngeal teeth or gizzard-like stomach in some fish, the gizzard of birds, or a combination of large cheek teeth and lateral or anterior-posterior movements of the mandible in most mammals. A stomach is absent in cyclostomes, some advanced species of fish, and the larval amphibians, but present in all other vertebrates. The stomach of most vertebrates is a unilateral dilatation of the digestive tract that serves for the storage of food and initial stages of digestion. However, the crop and proventriculus perform these functions in birds. Eight of the 20 mammalian orders include species with a stomach that is expanded and either haustrated or divided into permanent compartments. The stomach of most vertebrates contains regions of proper gastric glandular mucosa, pyloric glandular mucosa, and an additional region of cardiac mucosa in reptiles, some adult amphibians, and most mammals. It also includes a region of stratified squamous epithelium in species belonging to half of the mammalian orders. This led Oppel (1897) and Bensley (1902-1903) to the conclusion that the appearance of cardiac glandular and stratified squamous epithelium represented a regression of highly specialized proper gastric glandular mucosa to a less complex cardiac glandular mucosa, and then to a nonglandular stratified squamous epithelium. The presence of stratified squamous epithelium in the stomachs of ant and termite-eaters, and many herbivores suggests that it serves a protective function against physical damage from food, analogous to chitin in the foregut of mollusks and insects, or the peritrophic membrane in the midgut of insects. However, the absorption and buffering of SCFA by the stratified squamous region of the ruminant forestomach also protects gastric epithelium against the damaging effects of rapid SCFA absorption at the low pH produced by the proper gastric glandular region of the stomach. Therefore, expansion of the stratified squamous epithelial region may have been the most parsimonious response to the need for gastric expansion in herbivores. As with many of the advanced invertebrates, the intestines of fish, larval amphibians, and some mammals lack a distinct hindgut. However, the intestine of most terrestrial insects and vertebrates consists of a midgut and hindgut. The midgut of these animals is the principal site for digestion and absorption. The hindgut aids in the recovery of electrolytes, nitrogen, and water and serves as the principal site of microbial digestion in most species. The kidneys of vertebrates carry out the excretory functions of the insect’s Malpighian tubules. However, these excretions enter the gut at the cloaca of the adult amphibians, reptiles, and birds, which may account for the appearance of antiperistalis in the hindgut of terrestrial vertebrates. Endothermy required a more rapid processing of food and digesta by birds and mammals, and additional mechanisms for the selective retention of bacteria and plant material by the herbivores. Haustra in the hindgut of many and the foregut of some herbivorous mammals aid in digesta retention. The colonic separation mechanisms of avian and mammalian cecum fermenters and compartmentalization of the stomach of many mammalian foregut fermenters provided another means for the selective retention of bacteria and plant fiber. Other than the phagocytic properties of midgut cells in some mammalian neonates, food is digested initially by extracellular enzymes secreted by the salivary, gastric, or pancreatic glands of vertebrates and reduced to smaller units by enzymes in the brush border and cytosol of midgut absorptive cells. Many of the digestive enzymes and mechanisms of nutrient absorption appear to have first evolved in the invertebrates. Although the exocrine pancreatic glands are distributed along the midgut of cyclostomes and some of the more advanced species of fish, they are consolidated in a compact pancreas in most vertebrates. The liver is also a compact organ in all vertebrates and no longer serves as a site for digestion or absorption of nutrients. However, the midgut functions as the principal site of digestion and absorption, and the hindgut conserves electrolytes, water, and nitrogen much as they do in the insects. Neuroendocrine Control: Neurotransmitter-like substances, peptides, and other messenger molecules are found in protozoa, and a nervous system is present in sponges and well developed in annelids, mollusks, arthropods, and vertebrates (Fig. 12.13). The motor, secretory, digestive and absorptive functions of the vertebrate digestive system are controlled and integrated by a variety of peptides and other substances secreted by neurons and endocrine cells. The neurons of vertebrates secrete purines, amines, peptides and other agents that either modulate the release of neurotransmitters by other neurons or have a direct effect on muscle, secretory, or absorptive cells. Extrinsic innervation of the gastrointestinal tract by vagal (cranial) nerves is limited to the stomach of fish (Fig. 11.4). The sacral cholinergic nerve supply to the hindgut seems to have appeared with the evolution of a distinct hindgut in the adult amphibians (Fig, 11.5). Figure 12.13. Evolution of biochemical elements of the nervous and endocrine systems. (Modified from Le Roith et al. 1982) Figure 11.4. Diagrammatic representation of the autonomic cholinergic excitatory (red line), adrenergic (yellow line) and nonadrenergic inhibitory (blue line) nerves to the stomach of vertebrates. (From Burnstock 1969). (From CD Chapter 11) Figure 11.5. Diagrammatic representation of the autonomic cholinergic excitatory (red line), adrenergic (yellow line) and nonadrenergic inhibitory (blue line) nerves to the intestine of vertebrates. (From Burnstock 1969). (From CD Chapter 11) The motor, secretory, digestive, and absorptive activities of the digestive system are affected by a variety of peptides secreted by endocrine cells. Many of these peptides are released by both nerves and endocrine glands and serve as neurotransmitters, neuromodulators, hormones, or paracrine agents. This led to the theory that hormones evolved from neuroectodermal tissue (Pearse 1969). Barrington (1962) advised caution in interpreting fragmentary evidence collected from a few species and pointed out that many adaptations are determined by the evolution of receptors and the modulation of programming by receptor cells, rather than changes in the structure of these peptides. However, some families of hormones appear to have evolved from ancestral peptides that served initially as neurotransmitters or modulating agents. Pancreatic polypeptide-like activity has been reported in the nervous system of earthworms, mollusks, and insects, and substance P has been identified in the nervous system of coelenterates, prochordates, and all classes of vertebrates (Stevens and Hume 1995). Vasoactive intestinal polypeptide (VIP) is considered the ancestral form of peptide in the secretin family, because of its presence in the nervous tissue of prochordates, but secretin activity was also reported in prochordates and mollusks. The CCK/gastrin family of peptides is believed to have evolved from an ancestral peptide rather than through parallel evolution (Vigna 1983; 1986). CCK-like peptides are found in all classes of vertebrates and it is the only member of this family found in cyclostomes. Although, extracts of the hagfish intestine stimulated contraction of the guinea pig gallbladder, the hagfish gallbladder was not stimulated by either these extracts or mammalian CCK, suggesting the absence of appropriate receptors. Vigna (1986) concluded that a CCK-like agent was present in prochordates and persisted in mammals, but its regulation of gallbladder contraction did not appear until after the evolution of cyclostomes. A CCK-gastrin like hormone stimulates HCl secretion in chondricthyean fish, but appears to have been lost in present-day osteicthyeans and possibly replaced by bombesin. Bioassay procedures that discriminate between CCK and gastrin suggest their divergence between the evolution of elasmobranch and teleost fish. However, the gallbladder of coho salmon responded to both CCK and gastrin (Vigna and Gorbman 1977), and radioimmunoassay and immunostaining studies indicate the appearance of a separate gastrin-like peptide at the divergence of amphibians and reptiles. Therefore, Vigna (1986) concluded that the functional evolution of these hormones involved recruitment of new targets for old hormones, new cellular sources for old hormones, and old targets for new hormones. Evolution of herbivores: One of the major advances in evolution was the advent of animals that can derive a substantial amount of their nutritional requirements from the leaves, petioles, or stems of plants. The abundance and availability of this plant material throughout the year opened the way to a much wider range of diets and ecological niches. However, the ability to derive nutrients from these fibrous portions of the plant, requires the ingestion of large quantities of plant material, its breakdown into small particles, and either its rapid passage through the gut or its retention for microbial fermentation of the cell walls. The first option was adopted by some invertebrates, most fish, the larval amphibians, emu, and panda. The second option requires cellulytic enzymes of either endogenous or microbial origin. Evolution of herbivores - Invertebrates: Buchner (1965) discussed the widespread distribution of endosymbiosis among invertebrates. Algae in the cells of Paramecium bursa provide their host with O2 and carbohydrates, and allow its survival in the absence of a normal food supply if there is sufficient light for photosynthesis. Bacteria in the endoplasm of the amoebae Pelomyxa are believed to be responsible for their ability to digest filter paper. A similar inclusion of bacteria has been observed in cells lining the digestive tract of some invertebrates, and large numbers of bacteria (and sometimes, protozoa) are found in the lumen of the digestive tract of many advanced species. Methanogenic bacteria were found in the digestive tracts of cockroaches, termites, millipedes, and scarab beetles (Hackstein and Stumm 1994). Microbial fermentation has been demonstrated in the gut of annelids, mollusks, echinoderms, and insects. The microbes are most concentrated in the crop of cockroaches, the midgut or midgut ceca of some herbivores, and in the expanded hindgut of termites (Fig, 12.14). Figure 12.14. Alimentary tract of termite Eutermes; a) esophagus, b) crop, c) proventriculus, d) midgut, e) Malpighian tubules, f) hindgut, g) rectal valve, h) rectal pouch, i) terminal rectum. (Modified from Wigglesworth 1962) Although cellulose digestion is well documented in a number of insects, there is disagreement over how it is accomplished. Wigglesworth (1984) stated that protozoa are the chief agents of cellulose digestion in woodeating termites. Martin (1991) contended that cellulose is digested by bacteria and protozoa, based partly on the assumption that the complete cellulase complex of exo-1,4-glucanase and endo-1,4-glucanase is required and insects are unable to synthesize the exo-glucanase (cellobiohydrolase). However, Slaytor (1992) concluded that there is no evidence that an exo1,4-glucanase is either involved in or required for cellulose digestion in termites or wood-eating cockroaches, and endo-1,4-glucanase is found in the salivary glands, foregut, and midgut of these insects. He concluded that although there is evidence that bacteria are involved in cellulose digestion in the gut of these and other invertebrates, the evidence is often weak. Evolution of herbivores - Vertebrate herbivores: The earliest vertebrate herbivores were probably fish that adopted pharyngeal teeth, a gizzardlike stomach, or microfiltration for the reduction of aquatic plants to a smaller particle size and used their midgut as the principal site for microbial fermentation. However, the expansion of vertebrates into terrestrial habitats required a larger gut capacity and longer retention time for the fermentation of plants that contained higher levels of structural carbohydrates. Although gut capacity increases with body mass, an increase in body mass also requires the ingestion of larger quantities of the more readily available but less readily fermentable forage. Evolution of the hindgut for the conservation of electrolytes and water by terrestrial vertebrates provided a site for the more prolonged retention of digesta and multiplication of larger populations of indigenous bacteria. Figure 12.12. Phylogenetic origins of various groups of vertebrates. A) Urodela, B) Lepospondylii, C) Apoda, D) Anura, E) Labyrinthodontia, F) Apisidospondylii, G) Chelonia, H) Anapsida, I) Cotylosauria, J) Eurapsida, K) Diapsida, L) Eosuchia, M) Squamata, N) Rhyncocephalia, O) Ornithischia, P)Thecodontia, Q) Synapsida, R) Parapsida, S) Pelycosauria, T) Pterosauria, U) Crocodilia, V) Aves, W) Saurischia, X) Prototheria, Y) Metatheria, Z) Pantotheria, AA) Therapsida, BB) Eutheria, CC) Ichthyosauria. (Modified from Torrey 1971) Dinosaurs: The appearance of herbivorous prosauropods in the Late Triassic was followed by the rapid radiation of two major groups; the saurischians and ornithiscia (Fig. 12.12). Saurischian dinosaurs included a wide range of herbivorous species and the ornithiscians appear to have been exclusively herbivores (Fastovsky and Weishampel 1996). Gymnosperms (cycads, cycadoids and conifers) and pteridophytes (ferns and other free-sporing plants) were the predominant terrestrial plants during the Triassic and Jurassic, but angiosperms (flowering plants) became the predominant plants during the Cretaceous Period (Coe et al. 1985; Taggart and Cross 1997). The body masses of species in 220 dinosaur genera were estimated to have ranged from 1-70,000 kg on nearly every continent and during most stages of the Mesozoic (Peczkis 1994). Although the Jurassic ornithischians were relatively small, the herbivorous sauropods of the late Jurassic included the largest terrestrial vertebrates of all time. The largest sauropods peaked in diversity during the Late Jurassic, but by the late Cretaceous the dominant herbivores were ornithischians of intermediate body weight. Studies of bone histology and growth rates, and other characteristics indicate that the dinosaurs were endotherms (Bakker 1971; Robertshaw 1984; Barrick and Showers 1994; Chinsamy and Dodson 1995; Fisher et al. 2000). However, Chinsamy and Dodson (1995) concluded that elevated growth rates and endothermy may have arisen independently in different groups of dinosaurs, and the general consensus is that the metabolic rate of dinosaurs fell between that of modern reptiles and mammals (Fastovsky and Weishampel 1996). The saurischian herbivores had cropping teeth, and the presence of polished stones with their fossil remains suggests that they used a gastric mill for the breakdown of plant material (Bakker 1987; Barrett and Upchurch 1995; Currie 1997) in a manner similar to that seen in the modern day mullet, birds, and some reptiles. However, the Cretaceous ornithiscians included two diverse groups of herbivores with a jaw musculature and dental batteries suitable for the reduction of plant material to a small particle size (Fastovsky and Weishampel 1996). The hadrosaur masticatory apparatus included cheek teeth that formed oblique shearing blades and a jaw articulation that allowed lateral rotation of the jaws (Fig. 12.15). The masticatory apparatus of ceratopsians consisted of a dense cluster of cheek teeth with uneven occluding surfaces and massive jaw muscles. Figure. 12.15. Unlike mammals (A), ornithopod dinosaurs (B) had jaws of equal width and cheek teeth that interlocked to form oblique, shearing surfaces. (From Norman and Weishampel 1985) Marshall and Stevens (2000) concluded that the large body size of most sauropod herbivores and the rarity and inherent inefficiency of foregut fermentation in present-day herbivores with a gastric mill suggest that the sauropod herbivores were colon fermenters. However, the masticatory apparatus of the ornithiscian hadrosaurs and ceratopsians would have satisfied the requirements for foregut fermentation, and the body mass of many species (Tables 12.1a,b) was less than that of the largest modern-day hippos or Tertiary ground sloths. Furthermore, the maximum body weight of hadrosaur and ceratopsian foregut fermenters could have exceeded that of present-day mammalian foregut fermenters if they had a lower rate of metabolism or a kangaroo-like forestomach less restrictive on forage intake. Table 12.1a. Body masses calculated from scale model (M), pelvic height (P), femur diameter (F), or humerus diameter (H). Asterisk denotes body mass cited by authors, all other values are estimates from information provided by authors. (modified from Peczkis 1994) Table 12.1b. Body masses calculated from scale model (M), pelvic height (P), femur diameter (F), or humerus diameter (H). Asterisk denotes body mass cited by authors, all other values are estimates from information provided by authors. (modified from Peczkis 1994) A hadrosaur dinosaur Gryptosaurus from the late Cretaceous of Alberta, Canada. (Weishampel & Young 1996) A ceratopsian dinosaur Styracosaurus of western America from the late Cretaceous of western America. (Weishampel & Young 1996) Farlow (1987) concluded that foregut fermentation would be of little advantage to herbivorous dinosaurs beyond its ability to remove plant toxins that interfere with metabolism. However, a more complete extraction of energy from forage and greater ability to conserve water would have allowed their adaptation to climates and habitats far less suitable to other herbivores. Therefore, evolution of foregut fermenters may have contributed to the diversification and distribution of the Cretaceous ornithiscians. The digestive strategies of ornithiscians could be explored by studies of microbial fermentation of the present-day angiosperms, gymnosperms, and pteridophytes that were listed by Taggart and Cross (1997) as similar to those of the Cretaceous Period. Bacteria could be collected from the hindgut or foregut of modern-day herbivores, including those that have adapted to high levels of tannin in their diet, such as the koalas (Osawa et al. 1993), spruce grouse (Pendergast and Boad 1973), and some tortoises (Swain 1976). Viable cultures can be maintained for up to four weeks to determine their adaptability to substrates and measure microbial fermentation over a range of temperatures and retention times (Dr. Vivek Fellner, Department of Animal Science, North Carolina State University - personal communication). Models based on chemical reactor theory could then be employed to estimate the optimal digestive strategy of herbivorous dinosaurs as a function of their gut capacity (body mass), body temperature, and the digestibility of their diet. The massive extinction of plants and animals at the end of the Cretaceous Period removed the dinosaurs (Fig. 12.12) and the other terrestrial vertebrates over 10 kg in body weight. The paucity of present-day reptilian herbivores has been attributed to an inefficient masticatory apparatus and the small body mass (gut capacity) of most species. A small body size provides a greater ratio between surface area and body mass for rapid equilibration between the body and environmental temperature. However, the fact that the largest present-day reptilian herbivores are an arboreal lizard and tortoises with a protective carapace suggests that their size may also have been limited by predation by mammals. Figure 12.12. Phylogenetic origins of various groups of vertebrates. A) Urodela, B) Lepospondylii, C) Apoda, D) Anura, E) Labyrinthodontia, F) Apisidospondylii, G) Chelonia, H) Anapsida, I) Cotylosauria, J) Eurapsida, K) Diapsida, L) Eosuchia, M) Squamata, N) Rhyncocephalia, O) Ornithischia, P)Thecodontia, Q) Synapsida, R) Parapsida, S) Pelycosauria, T) Pterosauria, U) Crocodilia, V) Aves, W) Saurischia, X) Prototheria, Y) Metatheria, Z) Pantotheria, AA) Therapsida, BB) Eutheria, CC) Ichthyosauria. (Modified from Torrey 1971) Birds and mammals: The earliest mammals are believed to have been small carnivores that appeared in the Jurassic Period (Crompton and Parker 1978). Marsupial and eutherian mammals diverged during the Early Cretaceous and the ancestors of modern-day eutherian herbivores had appeared by the Late Cretaceous (Fig. 12.12). The earliest herbivores may have been multituberculates, which were abundant during the Mesozoic and persisted into the Cenozoic (Krause 1982; Carroll 1988; Wall and Krause 1992). Figure 12.16 shows the evolution of angiosperms, rodents, artiodactyls, perissodactyls, and macropod marsupials during the Tertiary Period of the Cenozoic. Artiodactyls, perissodactyls, proboscideans, hyracoids, and sirenians stemmed from a common source of Cretaceous ungulates or hoofed animals (Prothero 1994). The climate of the Paleocene and Eocene Epochs of the Tertiary was warmer and wetter than that of today, and rain forests extended over much of the globe. However, a gradual cooling since the beginning of the Eocene was accompanied by the diversification of ungulates, rodents, and lagomorphs (Carroll 1987; Prothero 1994). Figure 12.16. Diversification of angiosperms, rodents, ungulates, and macropod marsupials during the Tertiary. The width of columns is a compromise between species diversity and density. (data on angiosperms: Van Soest 1994; data on rodents: Romer 1966; data on ungulates: Janis 1976; data on macropod marsupials: Hume 1978) (Modified from Stevens and Hume 1995) The Oligocene saw the appearance of grasslands, with a higher cellulose-lignin ratio, and further diversification of rodents, artiodactyls, perissodactyls, and macropod marsupials (Fig. 12.16). It was also accompanied by the diversification of cetaceans, which are closely related to the artiodactyls (Árnason et al. 1991; Adachi et al. 1993; Milinkovitch et al. 1993) and may have originated from terrestrial herbivores. This could account for the multicompartmental forestomach and high concentrations of bacteria and SCFA in both toothed and baleen cetaceans. Separation of the camelids and ruminants in the Middle Eocene suggests that rumination had already evolved in this lineage, but the appearance of the omasum as an important functional organ in the advanced ruminants seems to have occurred after their separation from tragulids in the Late Oligocene. The cooler, drier climates of the Miocene were accompanied by an expansion of grasslands and diversification of the perrisodactyls, proboscideans, hyracoids, sirenians, and lagomorphs (Romer 1966; Prothero 1994). However, the most pronounced diversification and radiation was seen in the advanced ruminants, rodents, and macropod marsupials (Fig. 12.16). The Cricetidae, which includes lemmings, voles, and many of the other herbivorous rodents, underwent a major expansion. The earliest fossils of macropod Potoridae were small ratkangaroos, with a dentition that was adapted to nonabrasive materials and has changed relatively little since (Bartholomai 1972; Sanson 1989). The earliest Macropodidae (wallabies and kangaroos) inhabited wet forests (Flannery 1984) and were probably similar to extant small (2-8 kg) wallabies, with lophodont dentition suitable for soft, nonabrasive forage (Hume 1978; Freudenberger et al. 1989). The latter half of the Tertiary saw a reduction in global temperatures and an increase in the body size of many mammals. Some became megaherbivores, over 1000 kg in body weight. The eutherian megaherbivores included hippos, camelids, bison, rhinoceroses, mammoths, mastodons, sirenians, and giant ground sloths (Owen-Smith 1988). The giant ground sloth Megatherium reached weights of 3400 kg. Baluchiterium, a rhinoscerotid that appeared in Asia during the Oligocene and early Miocene, had a shoulder height of 5 m and is believed to have been the largest terrestrial mammal of all time. The marsupial herbivores of Australasia included the giant wombat Phascolonus (150 kg), tapir-like Palorchestes (300 kg), wallaby Protomodon (50 kg), browsing, short-faced kangaroo Procoptodon, and Diprotodon the largest (1150 kg) marsupial herbivore (Hume 1999). The lower body weights of the largest marsupials, in contrast to eutherian mammals, has been attributed to the arid climates and poor replenishment of soil nutrients by tectonic activity in this region over the past 60 million years (Flannery 1994). By the end of the Pleistocene Epoch of glaciations most of the megaherbivores were extinct. The final extinction of most megaherbivores is attributed to a combination of climatic change and human predation (Owen-Smith 1988). The only modern-day megaherbivores are the elephants, white rhinos, giraffes, and hippos. Figure 12.16 shows that the diversification of mammalian herbivores since the beginning of the Miocene was greatly influenced by the evolution of rodent cecum fermenters and the artiodactyl and marsupial foregut fermenters. This would be further supported by the inclusion of the modern day cecum fermenting lagomorphs and arboreal marsupials, and foregut fermenting sloths and colobid monkeys. Therefore, the evolution of cecum and foregut fermenters played an important role in the diversification and distribution of mammalian herbivores. Fossil records of ptarmigan and ostriches were found in the Miocene (Johnsgard 1983; MourerChauviré et al. 1996). However, rhea and emu fossils date back only to the Pliocene, and fossil records of New Zealand moas and Madagascar elephant birds extend only to the Pleistocene (Carroll 1988). Figure 12.16. Diversification of angiosperms, rodents, ungulates, and macropod marsupials during the Tertiary. The width of columns is a compromise between species diversity and density. (data on angiosperms: Van Soest 1994; data on rodents: Romer 1966; data on ungulates: Janis 1976; data on macropod marsupials: Hume 1978) (Modified from Stevens and Hume 1995) The earliest mammalian herbivores may have been colon fermenters, as suggested by Hume and Warner (1980). However, the appearance of cecum fermenters would have allowed an earlier evolution of herbivory in smaller species. The earliest cecum fermenters may have fed on invertebrates and used cecal bacteria to break down chitin, the structural carbohydrate in the integument of many invertebrates. Chitinolytic bacteria are found in the midgut of fish, large ceca of many, and forestomach of baleen whales that feed on invertebrates (Stevens and Hume 1995). Evolution of a colonic separation mechanism allowed the selective retention of smaller, more rapidly digestible plant particles in the cecum and rapid transit of larger digesta particles through a relatively short colon. Periodic release of cecal contents (cecotrophy) would provide highly nutritious feces and enhance the advantages of the coprophagy, which is seen in most other mammals only on nutrient deficient diets (Stevens and Hume 1995). Lagomorphs, herbivorous rodents and arboreal marsupials, some herbivorous primates, and most avian herbivores have retained this strategy. Expansion in the body size of many herbivores may have increased the diameter of the colon, and reduced the effectiveness of the colonic separation mechanism. An increase in the length of the colon would also increase its absorptive and microbial digestive capacity, which would reduce the nutritional value of coprophagy. Therefore, the colon became the principal site of microbial fermentation in the largest avian and mammalian herbivores. Forestomach fermentation probably evolved in a series of changes that began with its expansion for food storage and use as a secondary site to cecum fermentation, as seen in the present-day hyracoids and some herbivorous rodents. This would be followed by further foregut expansion in species that fed on a mixture of plant concentrates and fiber, such as the present-day peccaries that inhabit Amazon forests (Bodmer 1989) and the smallest ruminants. Cooler, drier climates during the Miocene reduced the rapid rate of plant growth and lignification, and increased the spread of grasslands with a higher cellulose/lignin ratio. Cooler climates also encouraged the evolution of larger species, including the megaherbivores. A longer digesta retention time set limits on the forage intake and body mass of foregut fermenters. However, foregut fermentation allowed the recovery of microbial protein and Bvitamins and adaptation to regions where the climates were more arid, the forage was less digestible, or the plants were more toxic. This proved especially effective when combined with rumination and omasal filtration of digesta in the advanced ruminants. The diversification and distribution of vertebrates were influenced by many factors, including reproductive efficiency and defenses against predation. However, the advent of herbivores that could subsist on the fibrous portion of plants played a major role in the diversification and distribution of mammals and dinosaurs. The success of herbivorous mammals can be attributed to an efficient masticatory apparatus, and to the evolution of a colonic separation mechanism and expanded cecum in the smallest herbivores and an expanded proximal colon or forestomach in the larger herbivores. Convergence on the strategies of cecum and forestomach fermentation allowed the evolution of small and intermediate sized endothermic herbivores, and their distribution to a wider range of habitats. An evolution of foregut fermenters may have played a similar role in the diversification and distribution of Cretaceous dinosaurs. Summary & Conclusions: The digestive system of vertebrates consists of a headgut (mouthparts and pharynx), foregut (esophagus and stomach), midgut, hindgut, exocrine pancreas, and biliary system. The headgut serves for the procurement and physical breakdown of food. Food is passed through the esophagus to the stomach, where it is stored and undergoes the initial stages of digestion by HCl and pepsin. The midgut is principal site of digestion, which is aided by enzymes secreted by the pancreas and located in the lumen-facing membranes and contents of the midgut epithelial cells, and by bile salts that are secreted by the liver and stored in the gall bladder. The midgut is also the principal site for the absorption of nutrients by passive diffusion or carrier-mediated transport across its epithelial cells. Digestion is aided by large quantities of electrolytes and water secreted by the oral glands, pancreas, biliary system and gastrointestinal tract. Most of the electrolytes and water are reabsorbed by the midgut and hindgut. The hindgut is also the principal site for the microbial production and conservation of nutrients in most species. All of these activities are integrated and controlled by the nervous system, hormones and paracrine agents. The effectiveness and efficiency of this basic design is demonstrated by a n analogous arrangement in the digestive system of many advanced invertebrates. Despite these common characteristics, the vertebrate digestive system shows a wide range of adaptations to the diet, habitat, or other characteristics of the species. Food can be reduced to a smaller particle size by teeth that are located in the jaws, oral cavity, or pharynx, or by microfiltration, a beak, or a gastric mill. The stomach is absent in cyclostomes, some advanced species of fish, and the larval amphibians, and its functions are served by the crop and proventriculus in birds. The stomach secretes neither HCl nor pepsinogen in a few species of mammals, and it includes an expanded, haustrated or compartmentalized forestomach in a few others. Pancreatic tissue is distributed along the intestine of some fish, and a gall bladder is absent in some fish and mammals. The hindgut varies from a short segment of intestine that is difficult to distinguish from the midgut in most fish, the larval amphibians, and some birds and mammals, to a voluminous, haustrated, and compartmentalized large intestine in some mammals. The hindgut of amphibians, reptiles and birds aids in the recovery of electrolytes, nitrogen, and water from the urinary excretions, as well as the secretions of the digestive system. Although many of the endogenous enzymes, indigenous microbes, mechanisms for secretion and absorption, and neuroendocrine agents are found in all classes of vertebrates, they can vary in their presence, composition, or location. Many adaptations of the digestive system are related to the habitat or diet of the species. Gill rakers or pharyngeal teeth allow some fish to swallow smaller particles of food without loss through the gills. Absorption of Na+ and Cl- by the esophagus aids others in their adaptation to a marine environment. Retention of digesta in a more highly developed hindgut allows terrestrial vertebrates to conserve electrolytes and water. A longer hindgut allows the more efficient conservation of water in species that inhabit arid environments. The digestive tract of carnivores and animals that feed on plant concentrates tend to be relatively simple in structure, with a rapid rate of digesta passage and episodic release of digestive fluids and enzymes. Omnivores tend to have a more complex digestive tract and longer digesta retention time. The high fiber diet of herbivores requires the ingestion of large quantities of plant material, a larger gut capacity, a longer digesta retention time, and a more continuous and voluminous secretion of electrolytes and water. The evolution of herbivores played an important role in the diversification and distribution of vertebrates, especially the mammals and dinosaurs. The success of mammalian herbivores can be attributed to an improved masticatory apparatus and the appearance of cecum and foregut fermenters. The success of the herbivorous dinosaurs also can be attributed to an effective masticatory apparatus or gastric mill and, possibly, the appearance of foregut fermenting ornithiscians. The previous sections include many examples of the contributions of comparative physiology to the understanding of basic physiological mechanisms. Many of these contributions have derived from the differences rather than the similarities among species. The nonglandular, stratified squamous epithelium of the frog skin and ruminant forestomach offered a much simpler system for the study of electrolyte and short-chain fatty acid transport mechanisms. Relationships between the rates of metabolism, food intake, digesta passage, digestion, and absorption are best studied in either ectotherms or endotherms with a temperature span wider than that of most birds and mammals. The ability to access and sample the forestomach contents of ruminants provided the basic information on the composition of indigenous gut bacteria and their contribution to the production and conservation of nutrients. The equine large intestine provided a unique model for the compartmental analysis of secretion, absorption, and digesta flow. Variations in the neurotransmitters, neuromodulators, hormones and paracrine agents of different classes of vertebrates helped describe their functions and how they evolved. One of the most compelling reasons for the study of comparative physiology of the digestive system is the information it provides for the maintenance of domesticated and captive animals, conservation of wildlife, and preservation of endangered species. Many diseases of domesticated and captive animals can be traced to an improper diet or feeding schedule, and the survival of free-ranging species rests on a delicate balance between plants, herbivores, omnivores, and carnivores. Interruption of the food chain at any level can affect their survival. Removal or poisoning of their natural habitat can guarantee their extinction. The digestive tract is the major portal of entry for both nutrients and the toxic agents. Yet the digestive systems of many mammalian species have received little or no study. Birds and reptiles have received less study, and the amphibians and fish that comprise almost half of the vertebrate species have received the least attention. Although survival of these animals is dependent on the food chain, we know very little about the composition of the diet of many of these species and how it changes with the seasons or migration. For example, how do the endogenous and microbial enzymes match the storage carbohydrates of algae or the wax esters of plankton in the diet of many animals at the base of the food chain? Do the endogenous and microbial chitinases play a significant role in the many species that feed on marine or terrestrial invertebrates? Is the ability of the hindgut of some birds to switch from Na+ - H+ exchange to eletrogenic Na+ absorption on low Na+ diets shared by some reptiles or other vertebrates? The renewed interest in the comparative physiology of the digestive system and the application of new methods and techniques to a wider range of species is encouraging. These studies should provide a better understanding of the basic mechanisms and how they are disrupted by abnormal diets, unnatural feeding practices, and digestive diseases. They will also provide information that is needed for the maintenance of domesticated and captive animals, conservation of wildlife, and preservation of endangered species.