Intro Metabolism
advertisement

Note Set 10
1
2000
Chapter 12: INTRODUCTION to METABOLISM
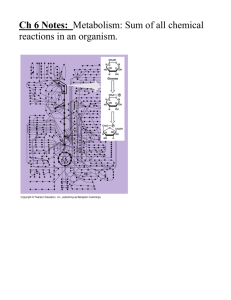
Living cells carry out thousands of reactions at once. Each reaction is controlled so that
unwanted or toxic accumulations or deficiencies of intermediates do not occur. Reaction
are often mechanistically complex and stereoselective…and even more remarkable, the
reactions all proceed under quite mild conditions.
A First Look at Metabolism
Specific reaction pathways
Control mechanisms that regulate flux (intracellular reaction rates through pathways)
Simplified view: Fig 12.1
Two major categories: Catabolism (degradation of complex substances) and Anabolism
(synthesis of complex organic molecules)
Both occur in three stages of complexity:
Stage 1. Interconversion of complex lipids with monomeric intermediates
Stage 2. Interconversion of monomeric sugars, amino acids and lipids with still simpler
organic compounds
Stage 3. Ultimate degradation to, or synthesis from, inorganic compounds such as CO2,
H2O, and NH3
Energy-yielding pathways also provide intermediates for biosynthesis. For example,
TCA cycle α-ketoglutarate is a precursor for glutamate
Intermediary metabolism (IM): All reaction concerned with generating and storing
metabolic energy and using the energy for biosynthesis of low-molecular-weight
compounds and energy storage compounds (ATP, some carbohydrates such as
glycogen, fat). Does not include nucleic acid or protein biosynthesis. So, IM can be
thought of as those reactions that do not involve a nucleic acid template
Energy metabolism: The part of IM consisting of pathways that generate or store energy
Central pathways (CPs): the quantitatively major pathways that are present in all
aerobic organisms and account for large amounts of mass transfer and energy
generation
CPs source of fuel: Autotrophs (self-feeding), like most plant, make glucose and most
others molecules from inorganic carbon supplied as CO2. Heterotrophs (feeding on
others) use other organic compounds they consume to synthesize their organic
molecules
Note Set 10
2
2000
Respiration is the coupling of energy generation to the oxidation of nutrients by oxygen.
Aerobic organisms depend on respiration. These include virtually all multicellular
organisms and many bacteria.
Anaerobic organisms sometimes must live in an oxygen-free environment and do not
derive their metabolic energy from oxygen
The sun can be thought of as the ultimate source of biological energy for organisms that
use CO2 that undergoes photosynthetic carbon fixation. However, organisms like
thermophiles that live at 100°C in hydrothermal vents in the ocean or geothermal
vents in active volcanoes do not get most of their energy from sunlight…
Freeways on the Metabolic Road Map
Central Pathways of Energy Metabolism
Fig 12.2 is the basic road map. Useful for overview. First consideration: energy
metabolism and and the central pathways.
Catabolism of carbohydrates and lipids, and their biosynthesis. Also see directional
flow on the map, sometimes toward catabolism, sometimes toward anabolism,
sometimes going both ways
Glycolysis, a Stage 2 pathway for degradation of carbohydrates in both aerobic and
anaerobic cells (Fig 12.3)
Leads to pyruvate, 3-carbon keto acid
Pyruvate can be reduced to ethanol and CO2 or lactate anerobically
Oxidation to acetyl-CoA (and CO2) is fate in respiration, then to citric acid cycle, the
main Stage 3
TCA accepts accepts simple carbon compounds from carbohydrates, but also lipids
and protein, and oxidizes them to CO2 (TCA) (Fig 12.4). An “on ramp” where all
catabolic pathways converge
Then on to electron transport and oxidative phosphorylation. The oxidative reactions
have produced reduced electron carriers (eg NADH) that are reoxidized to drive
synthesis of ATP
Fat feeds in by breakdown to acetyl-CoA by β-oxidation (Fig 12.5)
Amino acids feed in too (Fig 12.6)
If acetyl-CoA is not oxidized, it can be used to make FAT and STEROIDS
Biosynthesis like this uses NADPH to provide electrons
Note Set 10
3
2000
Fig 12.7 shows some bioxynthetic pathways for carbohydrates such as
gluconeogenesis and glycogen synthesis (in animals, starch in plants)
Fig 12.8 shows photosynthesis, the way green plants capture sunlight energy to drive
synthesis of ATP (energy) and reducing power (NADPH). Both are used for
carbohydrate synthesis.
Distinct Pathways for Biosynthesis and Degradation
Pathways seldom work precisely as the reversal of each other although they may use
some enzymes in common. There are always different enzymes catalyzing their
regulated steps. Ort, pathways may occur in different compartments, as fatty acid
synthesis occurs in the cytosol, breakdown in the mitochondria. If they both took place
in the same compartment, there would be a futile cycle as in “make it then break it
down ad infinitum.” ATP and all components would be wasted this way
1. The pathways have to be exergonic in the direction of flux…this could not be true
BOTH ways
2. The flux must respond to the bioenergetic status of the cell, as in when ATP is high or
low
A very important consideration is to understand that the enzymes regulating reverse
steps in pathways such as glycolysis and gluconeogenesis respond to allosteric
effectors such that one enzyme is inhibited by conditions that activate the other. Two
opposed cellular reactions that are independantly controlled like this are called a
substrate cycle
Some Bioenergetic considerations
Oxidation as a Metabolic Energy Source
Unfavorable or endergonic reactions can proceed if coupled to a favorable exergonic
reaction.
Most of the energy needed is derived from oxidation of organic substrates, and oxygen,
the ultimate e- acceptor is a strong oxidant
Biological Oxidations: Energy Release in Small Increments
∆G the same if glucose polymer oxidized by burning in a fire or through the TCA cycle
and oxidative phosphorylation
C6H12O6 + 6CO2 à 6CO2 + 6H2O
∆G°’ = -2870 kJ/mol
If burned, all energy released as heat. In CETOP (citric acid cycle, electron transport,
oxidative phosphorylation) , about 40% is used to make ATP from ADP and PI
Note Set 10
4
2000
In CETOP e- are not transferred directly to lxygen but rather go through a series of
coupled oxidation-reduction reactions with e- passed to intermediates like NAD+ and
then finally O2. This sequence is called the electron transport chain or respiratory
chain, and oxygen is called the terminal electron acceptor. Small energy transfers
waste less energy than large ones.
Energy from other places than oxidation by oxygen, i.e. using a different e- acceptor:
Fermentations, no net change in oxidation state of substrate and product
Sulfur to sulfide (vent critters)
Nitrite to ammonia (some bacteria)
Energy Yields, Respiratory Quotients, and Reducing Equivalents
Caloric content: the number of calories from combustion
Calories = nutrition, Joules = biochemistry
Fat is more highly reduced than carbohydrate so has a higher caloric content and more
Joules…too. More reduced = more calories = more Joules
Respiratory Quotient (RQ) = {CO2 produced/O2 consumed}. The lower the RQ for a
substrate, the more O2 consumed per carbon oxidized, and thus more ATP could be
produced the lower the RQ
Fig 12.9 Major source of electrons for biosynthesis is NADPH. NAD enzymes usually
oxidize substrates (dehydrogenases), and NADPH enzymes reduce substrates
(reductases)
ATP as a Free Energy Currency
See Study Guide CD and Book
Thermodynamic Properties of ATP
See Study Guide CD and Book
Other High-Energy Nucleotides
See Study Guide CD and Book
Adenylate Energy Charge
See Study Guide CD and Book
Major Metabolic Control Mechanisms
Control of enzyme Levels
By: Synthesis and degradation and regulation…
Forms of regulation so far all modify the activity of enzymes already present
Enzymes can also be regulated at level of synthesis and degradation
Note Set 10
5
2000
Regulation of synthesis
Used for enzymes that are required under special circumstances, in specific cells, or at
particular stages of development
"housekeeping" enzymes are usually not regulated this way
Regulation occurs at the level of transcription of the gene or translation of the mRNA
encoding the enzyme
Examples:
E. coli lac operon
Enzyme induction occurs when there is a need to use lactose as a carbon
Enzyme synthesis is controlled at the level of transcription
Genes are not transcribed if there is glucose available and are transcribed only in
the presence of lactose
Egg protein synthesis in chicken oviduct
Required genes are transcribed at sexual maturity in response to estradiol
Regulation by enzyme degradation (proteolysis)
Enzyme produced for a particular situation may not be needed later, and may be
deleterious then
Enzymes have very different half lives
Some last many days, others only minutes
Specific mechanisms exist for proteolytic degradation
Some enzymes are targeted for rapid degradation
Rapidly degraded enzymes are often those that function at key control points
They rapidly disappear if not needed, and also can be rapidly resynthesized if needed
again
Damaged enzymes are also rapidly degraded
A faulty enzyme in a major pathway is not only worthless, it can also be damaging to
metabolic function
Note Set 10
6
2000
Different kinds of control are employed for different puposes, and the activity of any
particular enzyme can be regulated at several levels
Control of Enzyme Activity
See Study Guide CD and Book
Compartmentation
See Study Guide CD and Book
Hormonal Regulation
See Study Guide CD and Book
Distributive Control of Metabolism
See Study Guide CD and Book
Experimental Analysis of Metabolism
Goals of the Study of Metabolism
See Study Guide CD and Book
Levels of Organization at which Metabolism is Studied
Whole Organism
Isolated or Perfused Organ
Whole Cells
Cell-Free Systems
Purified Components
Metabolic Probes
See Study Guide CD and Book