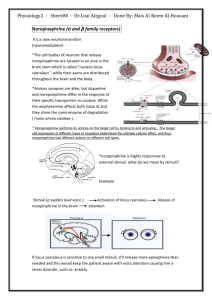
Tyrosine-Derived Neurotransmitters
advertisement

Tyrosine-Derived Neurotransmitters The majority of tyrosine that does not get incorporated into proteins is catabolized for energy production. One other significant fate of tyrosine is conversion to the catecholamines. The catecholamine neurotransmitters are dopamine, norepinephrine, and epinephrine (see also Biochemistry of Nerve Transmission). Norepinephrine is the principal neurotransmitter of sympathetic postganglionic endings. Both norepinephrine and the methylated derivative, epinephrine are stored in synaptic knobs of neurons that secrete it, however, epinephrine is not a mediator at postganglionic sympathetic endings. Tyrosine is transported into catecholamine-secreting neurons and adrenal medullary cells where catechaolamine synthesis takes place. The first step in the process requires tyrosine hydroxylase, which like phenylalanine hydroxylase requires tetrahydrobiopterin (H4B, or written as BH4) as cofactor. The dependence of tyrosine hydroxylase on H4B necessitates the coupling to the action of dihydropteridine reductase (DHPR) as is the situation for phenylalanine hydroxylase and tryptophan hydroxylase (see below). The hydroxylation reaction generates DOPA (3,4-dihydrophenylalanine). DOPA decarboxylase converts DOPA to dopamine, dopamine β-hydroxylase converts dopamine to norepinephrine and phenylethanolamine N-methyltransferase converts norepinephrine to epinephrine. This latter reaction is one of several in the body that uses SAM as a methyl donor generating S-adenosylhomocysteine. Within the substantia nigra and some other regions of the brain, synthesis proceeds only to dopamine. Within the adrenal medulla dopamine is converted to norepinephrine and epinephrine. Synthesis of the Catecholamines from Tyrosine. Once synthesized, dopamine, norepinephrine and epinephrine are packaged in granulated vesicles. Within these vesicles, norepinephrine and epinephrine are bound to ATP and a protein called chromogranin A. The actions of norepinephrine and epinephrine are exerted via receptor-mediated signal transduction events. There are three distinct types of adrenergic receptors: α1, α2, β. Within each class of adrenergic receptor there are several sub-classes. The α1 class contains the α1A, α1B, and α1D receptors. The α1 receptor class are coupled to Gq-type G-proteins that activate PLCγ resulting in increases in IP3 and DAG release from membrane PIP2. The α2 class contains the α2A, α2B, and α2C receptors. The α2 class of adrenergic receptors are coupled to Gi-type G-proteins that inhibit the activation of adenylate cyclase and therefore, activation results in reductions in cAMP levels. The β class of receptors is composed of three subtypes: β1, β2, and β3 each of which couple to Gs-type G-proteins resulting in activation of adenylate cyclase and increases in cAMP with concomitant activation of PKA. Dopamine binds to dopamineric receptors identified as D-type receptors and there are four subclasses identified as D1, D2, D4, and D5. Activation of the dopaminergic receptors results in activation of adenylate cyclase (D1 and D5) or inhibition of adenylate cyclase (D2 and D4). Epinephrine and norepinephrine are catabolized to inactive compounds through the sequential actions of catecholamine-O-methyltransferase (COMT) and monoamine oxidase (MAO). Compounds that inhibit the action of MAO have been shown to have beneficial effects in the treatment of clinical depression, even when tricyclic antidepressants are ineffective. The utility of MAO inhibitors was discovered serendipitously when patients treated for tuberculosis with isoniazid showed signs of an improvement in mood; isoniazid was subsequently found to work by inhibiting MAO. Metabolism of the catecholamine neurotransmitters. Only clinically important enzymes are included in this diagram. The catabolic byproducts of the catecholamines, whose levels in the cerebrospinal fluid are indicative of defects in catabolism, are in blue underlined text. Abbreviations: TH = tyrosine hydroxylase, DHPR = dihydropteridine reductase, H2B = dihydrobiopterin, H4B = tetrahydrobiopterin, MAO = monoamine oxidase, COMT = catecholamine-O-methyltransferase, MHPG = 3-methoxy-4-hydroxyphenylglycol, DOPAC = dihydroxyphenylacetic acid. Tryptophan-Derived Neurotransmitters Tryptopan serves as the precursor for the synthesis of serotonin (5-hydroxytryptamine, 5-HT, see also Biochemistry of Nerve Transmission) and melatonin (N-acetyl-5-methoxytryptamine). Pathway for serotonin and melatonin synthesis from tryptophan. Abbreviations: THP = tryptophan hydroxylase, DHPR = dihydropteridine reductase, H2B = dihydrobiopterin, H4B = tetrahyrobiopterin, 5-HT = 5-hydroxytryptophan, AADC = aromatic L-amino acid decarboxylase, SNA = serotonin N-acetylase, HOMT = hydroxyindole-O-methyltransferase. Serotonin is synthesized through 2-step process involving a tetrahydrobiopterin-dependent hydroxylation reaction (catalyzed by tryptophan-5-monooxygenase, also called tryptophan hydroxylase) and then a decarboxylation catalyzed by aromatic L-amino acid decarboxylase. The hydroxylase is normally not saturated and as a result, an increased uptake of tryptophan in the diet will lead to increased brain serotonin content. Serotonin is present at highest concentrations in platelets and in the gastrointestinal tract. Lesser amounts are found in the brain and the retina. Serotonin containing neurons have their cell bodies in the midline raphe nuclei of the brain stem and project to portions of the hypothalamus, the limbic system, the neocortex and the spinal cord. After release from serotonergic neurons, most of the released serotonin is recaptured by an active reuptake mechanism. The function of the antidepressant, Prozac®, and related drugs called selective serotonin re-uptake inhibitors (SSRIs), is to inhibit this reuptake process, thereby, resulting in prolonged serotonin presence in the synaptic cleft. The function of serotonin is exerted upon its interaction with specific receptors. Several serotonin receptors have been cloned and are identified as 5HT1, 5HT2, 5HT3, 5HT4, 5HT5, 5HT6, and 5HT7. Within the 5HT1 group there are subtypes 5HT1A, 5HT1B, 5HT1D, 5HT1E, and 5HT1F. There are three 5HT2 subtypes, 5HT2A, 5HT2B, and 5HT2C as well as two 5HT5 subtypes, 5HT5a and 5HT5B. Most of these receptors are coupled to G-proteins that affect the activities of either adenylate cyclase or phospholipase Cν (PLCγ). The 5HT3 class of receptors are ion channels. Some serotonin receptors are presynaptic and others postsynaptic. The 5HT2A receptors mediate platelet aggregation and smooth muscle contraction. The 5HT2C receptors are suspected in control of food intake as mice lacking this gene become obese from increased food intake and are also subject to fatal seizures. The 5HT 3 receptors are present in the gastrointestinal tract and are related to vomiting. Also present in the gastrointestinal tract are 5HT4 receptors where they function in secretion and peristalsis. The 5HT6 and 5HT7 receptors are distributed throughout the limbic system of the brain and the 5HT 6 receptors have high affinity for antidepressant drugs. Melatonin is derived from serotonin within the pineal gland and the retina, where the necessary N-acetyltransferase enzyme is found. The pineal parenchymal cells secrete melatonin into the blood and cerebrospinal fluid. Synthesis and secretion of melatonin increases during the dark period of the day and is maintained at a low level during daylight hours. This diurnal variation in melatonin synthesis is brought about by norepinephrine secreted by the postganglionic sympathetic nerves that innervate the pineal gland. The effects of norepinephrine are exerted through interaction with β-adrenergic receptors. This leads to increased levels of cAMP, which in turn activate the N-acetyltransferase required for melatonin synthesis. Melatonin functions by inhibiting the synthesis and secretion of other neurotransmitters such as dopamine and GABA. Creatine Biosynthesis Creatine is synthesized in the liver by methylation of guanidoacetate using SAM as the methyl donor. Guanidoacetate itself is formed in the kidney from the amino acids arginine and glycine. Synthesis of Creatine and Creatinine Creatine is used as a storage form of high energy phosphate. The phosphate of ATP is transferred to creatine, generating creatine phosphate, through the action of creatine phosphokinase. The reaction is reversible such that when energy demand is high (e.g. during muscle exertion) creatine phosphate donates its phosphate to ADP to yield ATP. Both creatine and creatine phosphate are found in muscle, brain and blood. Creatinine is formed in muscle from creatine phosphate by a nonenzymatic dehydration and loss of phosphate. The amount of creatinine produced is related to muscle mass and remains remarkably constant from day to day. Creatinine is excreted by the kidneys and the level of excretion (creatinine clearance rate) is a measure of renal function. Glutathione Functions Glutathione (abbreviated GSH) is a tripeptide composed of glutamate, cysteine and glycine that has numerous important functions within cells. Glutathione serves as a reductant; is conjugated to drugs to make them more water soluble; is involved in amino acid transport across cell membranes (the γ-glutamyl cycle); is a substrate for the peptidoleukotrienes; serves as a cofactor for some enzymatic reactions and as an aid in the rearrangement of protein disulfide bonds. GSH is synthesized in the cytosol of all mammalian cells. The rate of GSH synthesis is dependent upon the availability of cysteine and the activity of the rate-limiting enzyme, γ-glutamylcysteine synthetase (GCS). In the liver, major factors that determine the availability of cysteine are diet, membrane transport activities of the three sulfur amino acids cysteine, cystine and methionine, and the conversion of methionine to cysteine (see the Amino Acid Metabolism page). Numerous conditions can alter the level of GSH synthesis via changes in GCS activity and GCS gene expression such as oxidative stress, antioxidant levels, hormones, cell proliferation, and diabetes mellitus. Synthesis of Glutathione (GSH) Structure of GSSG The role of GSH as a reductant is extremely important particularly in the highly oxidizing environment of the erythrocyte. The sulfhydryl of GSH can be used to reduce peroxides formed during oxygen transport. Endogenously produced hydrogen peroxide (H2O2) is reduced by GSH in the presence of selenium-dependent GSH peroxidase. Hydrogen peroxide can also be reduced by catalase, which is present only in the peroxisomes. In the mitochondria, GSH is particularly important because mitochondria lack catalase. The resulting oxidized form of GSH consists of two molecules disulfide bonded together (abbreviated GSSG). The enzyme glutathione reductase utilizes NADPH as a cofactor to reduce GSSG back to two moles of GSH. Hence, the pentose phosphate pathway is an extremely important pathway of erythrocytes for the continuing production of the NADPH needed by glutathione reductase. In fact as much as 10% of glucose consumption, by erythrocytes, may be mediated by the pentose phosphate pathway. Detoxification of xenobiotics or their metabolites is another major function of GSH. These compounds form conjugates with GSH either spontaneously or enzymatically in reactions catalyzed by GSH S-transferase. The conjugates formed are usually excreted from the cell and, in the case of the liver they are excreted in the bile. Several mechanisms exist for the transport of amino acids across cell membranes. Many are symport or antiport mechanisms that couple amino acid transport to sodium transport. The γ-glutamyl cycle is an example of a group transfer mechanism of amino acid transport. Although this mechanism requires more energy input, it is rapid and has a high capacity. The cycle functions primarily in the kidney, particularly renal epithelial cells. The enzyme γ-glutamyl transpeptidase is located in the cell membrane and shuttles GSH to the cell surface to interact with an amino acid. Reaction with an amino acid liberates cysteinylglycine and generates a γ-glutamyl-amino acid which is transported into the cell and hydrolyzed to release the amino acid. Glutamate is released as 5-oxoproline and the cysteinylglycine is cleaved to its component amino acids. Regeneration of GSH requires an ATP-dependent conversion of 5-oxoproline to glutamate and then the 2 additional moles of ATP that are required during the normal generation of GSH. Polyamine Biosynthesis One of the earliest signals that cells have entered their replication cycle is the appearance of elevated levels of mRNA for ornithine decarboxylase (ODC), and then increased levels of the enzyme, which is the first enzyme in the pathway to synthesis of the polyamines. Because of the latter, and because the polyamines are highly cationic and tend to bind nucleic acids with high affinity, it is believed that the polyamines are important participants in DNA synthesis, or in the regulation of that process. The key features of the pathway are that it involves putrescine, an ornithine catabolite, and S-adenosylmethionine (SAM) as a donor of 2 propylamine residues. The first propylamine conjugation yields spermidine and addition of another to spermidine yields spermine. The function of ODC is to produce the 4-carbon saturated diamine, putrescine. At the same time, SAM decarboxylase cleaves the SAM carboxyl residue, producing decarboxylated SAM (S-adenosymethylthiopropylamine), which retains the methyl group usually involved in SAM methyltransferase activity. SAM decarboxylase activity is regulated by product inhibition and allosterically stimulated by putrescine. Spermidine synthase catalyzes the condensation reaction, producing spermidine and 5'-methylthioadenosine. A second propylamine residue is added to spermidine producing spermine. The signal for regulating ODC activity is unknown, but since the product of its activity, putrescine, regulates SAM decarboxylase activity, it appears that polyamine production is principally regulated by ODC concentration. The butylamino group of spermidine is used in a posttranslational modification reaction important to the process of translation. A specific lysine residue in the translational initiation factor eIF-4D is modified. Following the modification the residue is hydroxylated yielding a residue in the protein termed hypusine. Nitric Oxide Synthesis and Function Vasodilators, such as acetylcholine and bradykinin, do not exert their effects upon the vascular smooth muscle cell in the absence of the overlying endothelium. When acetylcholine (or bradykinin) binds its receptor on the surface of endothelial cells, a signal cascade, coupled to the activation phospholipase C-γ (PLCγ), is initiated. The PLCγ-mediated release of inositol trisphosphate, IP3 (from membrane associated phosphatidylinositol-4,5-bisphosphate, PIP2), leads to the release of intracellular stores of Ca2+. In turn, the elevation in Ca2+ leads to the liberation of endothelium-derived relaxing factor (EDRF) which then diffuses into the adjacent smooth muscle. Within smooth muscle cells, EDRF reacts with the heme moiety of a soluble guanylyl cyclase, resulting in activation of the latter and a consequent elevation of intracellular levels of cGMP. The net effect is the activation of cGMP-dependent protein kinase (PKG) and the phosphorylation of substrates leading to smooth muscle cell relaxation. The coronary artery vasodilator, nitroglycerin, acts to increase intracellular release of EDRF and thus the activation of the cGMP signal cascade. Quite unexpectedly, EDRF was found to be the free radical diatomic gas, nitric oxide, NO. So stunning was the elucidation of the pathway to and actions of NO that Drs. Murad, Ignarro and Furchgott were awarded the Nobel Prize in 1998 for their work on this system. NO is formed by the action of NO synthase, (NOS) on the amino acid arginine. arginine ——> citrulline + NO There are 3 isozymes of NOS in mammalian cells: Neuronal NOS (nNOS), also called NOS-1 Inducible or macrophage NOS (iNOS), also called NOS-2 Endothelial NOS (eNOS), also called NOS-3. Nitric oxide synthases are very complex enzymes, employing five redox cofactors: NADPH, FAD, FMN, heme and tetrahydrobiopterin (H4B). NO can also be formed from nitrite, derived from vasodilators such as glycerin trinitrate (nitroglycerin) during their metabolism. The half-life of NO is extremely short, lasting only 2-4 seconds. This is because it is a highly reactive free radical and interacts with oxygen and superoxide. NO is inhibited by hemoglobin and other heme proteins which bind it tightly. Both eNOS and nNOS are constitutively expressed and regulated by The calcium regulation is imparted be the associated calmodulin subunits, thus explaining how vasodilators such as acetylcholine effect smooth muscle relaxation as a consequence of increasing intracellular endothelial cell calcium levels. Although iNOS contains calmodulin subunits, its activity is unaffected by changes in Ca2+ concentration. iNOS is transcriptionally activated in macrophages, neutrophils, and smooth muscle cells. Ca2+. The major functions of NO production through activation of iNOS are associated with the bactericidal and tumoricidal actions of macrophages. Overproduction of NO via iNOS is associated with cytokine-induced septic shock such as occurs post-operatively in patients with bacterial infections. Bacteria produce endotoxins such as lipopolysaccharide (LPS) that activate iNOS in macrophages. Nitric oxide is involved in a number of other important cellular processes in addition to its impact on vascular smooth muscle cells. Events initiated by NO that are important for blood coagulation include inhibition of platelet aggregation and adhesion and inhibition of neutrophil adhesion to platelets and to the vascular endothelium. NO is also generated by cells of the immune system and as such is involved in non-specific host defense mechanisms and macrophage-mediated killing. NO also inhibits the proliferation of tumor cells and microorganisms. Additional cellular responses to NO include induction of apoptosis (programmed cell death), DNA breakage and mutation. Chemical inhibitors of NOS are available and can markedly decrease production of NO. The effect is a dramatic increase in blood pressure due to vasoconstriction. Another important cardiovascular effect of NO is exerted through the production of cGMP, which acts to inhibit platelet aggregation.