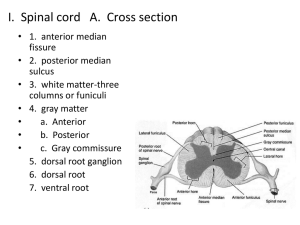
The Central Nervous System
advertisement

SPPA 2050 Speech Anatomy and Physiology Neuronal Structure and Function Now, let us move on to the neuron. As will be the general pattern of behavior for SPPA 205, we will begin discussion of the neuron with its basic structure, followed by its function. Although there is a great deal of diversity in the look of neurons, the vast majority of neurons have four structurally defined regions: the soma or cell body, the dendrites, the axon, and the presynaptic terminal (also called an axon terminal arbor, or “Bouton”). The cell body is the metabolic center of the neuron. Dendrites are often fine, tree-like extensions from the cell body. The axon also projects from the cell body and is a tubular extension that can be quite long with a terminal ending. Often the axon is wrapped in myelin, an insulating material (produced by oligodendrocytes and Schwann cells), punctuated with nodes of Ranvier. The myelin and nodes of Ranvier dramatically increase the speed at which neural signals can travel. Conduction velocity (the speed of the signal) is also influenced by the diameter of the axon. The presynaptic terminal of the axon is located very, very close to the dendrite or cell body of an adjacent neuron (or a muscle cell – remember the muscle). The region where cell-to-cell communication can occur is called the synapse and the region between the cells is called the synaptic cleft. Neurons are polarized, meaning that information flows in a predictable and consistent direction. This flow runs from the dendrites and cell body to the axon and finally to the axon terminal where it has the potential of communicating with other cells. The means of communication for the neuron is electrical and we will spend a little time discussing how this electrical charge is (1) generated at the receiving end of the neuron, (2) propagated along the length of the neuron, and (3) transmitted to the neighboring neurons. The neuron has a separation of electrical charge across its cell membrane. A separation in charge simply means an unequal number of positively and negatively charged elements (charged atoms or ions, principally sodium, which has a positive charge, potassium, which also has a positive charge and chlorine, which has a negative charge) inside and outside the cell. This is termed the resting membrane potential. The resting membrane potential is negative indicating that the inside of the cell is more negative than the outside of the cell. For a lot of neurons the resting membrane potential is about –50 - -70 millivolts (1 millivolt = 1/1000 of a volt). The resting membrane potential is established through active transport of ions across the cell membrane. Active transport simple means that cellular energy (the active part) is needed to move the ions. This is accomplished by a mechanism called the Sodium-Potassium pump (or Na-K pump). The Na-K pump moves Na+ (sodium) ions out of the cell and moves K+ (potassium) ions into the cell. Na+ and K+ have a single positive charge. The pump moves three Na+ ions out of the cell for every two K+ that it lets in the cell. This causes an electrical gradient, where the charge inside the cell is less than the charge outside the cell. Also, most of the Na + outside and K+ inside the cell creates a chemical concentration gradient. This electrochemical gradient is the resting membrane potential. If allowed, the Na+ ions would like to rush inside the cell to equalize both their numbers relative to the outside of the cell and the equalize the negative charge across the cell membrane. However, ions such as Na+ must pass through protein channels that are specially designed for them. These channels may be open or closed depending upon the electrochemical environment. At resting membrane potential, most of these channels are closed. Some ion channels have special receptor sites on the outside of the cell. These receptor sites are much like a lock in a door (which is the channel). It requires the correct key to open. This “key” takes the form of a biochemical (often a neurotransmitter or NT). If there is a NT in the area it can attach itself to the ion channel receptor site and change the permeability of that ion channel (e.g. unlock and open the door). Opening or closing these receptor channels (Figure 2.7) will change the permeability of the channels to ions and therefore can change the membrane potential. There is another type of ion channel that you need to be aware of. These channels do not have receptor sites and therefore do not operate in the lock and key manner. These voltage gated channels SPPA 2050 Speech Anatomy and Physiology (Figure 2.7) will change their permeability depending upon the membrane potential. If there is a change in the membrane potential, these channels may open (or close). For example, a NT may attach to a receptor site and open a Na+ channel. Given the electrochemical gradient that exists, the Na+ will move into the cell making the membrane potential less negative. This change in membrane potential could trigger a voltage gated Na+ channel to open causing more opportunity for Na+ to enter the cell and further change membrane potential. There is a third type of channel that occurs in specialized settings. Many sensory neurons (those that sense touch, pressure etc) have ion channels that are opened/closed because of mechanical events in the environment. For example, the hair cell in the ear has a small projection that is moved, indirectly, by sound pressure. This movement of the ‘hair’ on the hair cell causes a mechanical change in the permeability of ion channels, which change the membrane potential. Neural signals are generated by changing this resting membrane potential (using the aforementioned mechanisms). Increasing the resting potential (making it more negative) is called hyperpolarization. Decreasing the resting potential (making it less negative) is called depolarization. Hyperpolarization decreases the cell’s ability to generate a signal called an action potential (discussed later) and is called inhibitory. Depolarization increases the cell’s ability to generate an action potential and is therefore called excitatory. As outlined before, signal transmission within the neuron involves receiving a signal from a source, propagating the signal along the length of the neuron, and transmitting the signal to other neurons. We’ll deal with each phase separately. Signal reception typically occurs at the dendrite and cell body. These signals are called potentials and either depolarize (excite) or hyperpolarize (inhibit) the cell (Figure 2.5). These potentials may come from sensory organs, such as a muscle spindle (generator potentials) or from other neurons (postsynaptic potentials or PSP). If the potential excites the cell, they are typically called excitatory postsynaptic potentials or EPSPs for short. Those that inhibit the cell are called inhibitory postsynaptic potentials or IPSPs. EPSPs and IPSPs are small and are often described as graded (can take on a variety of magnitudes – large or small), local (potential only occurs in the region of the receiving end of the neuron) and passive (potentials die out rather quickly). At any given point in time, there may be a large number of these potentials (both excitatory and inhibitory) being generated in different parts of the receiving end of the cell. There is a region called a trigger zone located near this receiving area that is quite sensitive to these changes in the membrane potential. At the trigger zone the various inhibitory and excitatory potentials will summate (add up). Summation can occur because there are numerous PSPs occurring at the same time in different places along the cell membrane (spatial summation) or a sequence of PSPs occur rapidly in time at the same point on the cell membrane (temporal summation) (Figure 2.6). If the sum of the potentials is excitatory and large enough to exceed some threshold value that is characteristic of that neuron, then an action potential will be initiated (Figures 2.8-9). The action potential is a very large, brief depolarization (up to 100 mV, about 1 ms in length). Unlike the receptor/synaptic potentials, which are local and passive, the action potential is active (is assisted by the cell) and will propagate the length of the axon without a reduction in its amplitude. While the changes in the receptor/synaptic potential are “graded”, meaning they can be produced at a variety of levels (positive, negative, bigger, smaller), the action potential is an all-or-none phenomenon. There cannot be a “graded” or partial response. It either happens or it doesn’t. Once it does happen, the action potential quickly moves down the length of the axon to the presynaptic terminal. The myelin sheath speeds up the rate of travel as does the nodes of Ranvier. The action potentials actually skip from one node to the next. Once the action potential reaches the presynaptic terminal, the next phase of signal transmission begins – the transmission to neighboring cells. This is described in the next paragraph. Recall that the action potential is a brief, large depolarization. Following this depolarization, there is a rapid return back toward the resting SPPA 2050 Speech Anatomy and Physiology potential. This process takes a little time and actually results in a hyperpolarization of the cell for a short period of time. What this means is that for a brief period of time, it is impossible to generate another action potential and for a longer period of time it is more difficult to generate an action potential. This brief period where another action potential is impossible is called the refractory period of the neuron. It is important because it provides a limit on how fast a neuron can fire (which can be as high as 1000/sec). In fact, the only way that a neuron can code information is through how fast it fires (rate code) and when it fires (temporal code). Refer to the figure below to get a visual image of this process. The curve above the trace of the membrane potential simply implies that the required excitatory potential to elicit another action potential is higher when closer in time to the previous action potential. Once the action potential reaches the presynaptic terminal, there has to be a way for information to span the synaptic cleft to communicate with neighboring cells. This requires a change in the mode of communication from electrical to biochemical. When the action potential reaches the presynaptic terminal, it initiates something called a secretory potential, which mediates the release of a chemical called a neurotransmitter into the synaptic cleft. The neurotransmitter diffuses across the cleft to the neighboring cell where it interacts with the cell membrane of the postsynaptic cell (if it is a neuron, it will be the dendrite or cell body) and produces a change in the membrane potential of that cell (either excitatory or inhibitory, depending on what the neurotransmitter is). Now this potential is unlikely to be big enough to elicit an action potential all on its own. If this part is starting to sound familiar, that is because we already talked about it earlier. This is the postsynaptic potential, but now it is happening in the next cell in the chain or network. Everything we discussed earlier now applies to this new neuron. We’ve come full circle in the physiology of the neuron. Now a few points about neurotransmitters (NT). NT’s come in a variety of types. There are dozens of NTs in the nervous system (e.g. glutamate, glycine, seretonin, dopamine, acetylcholine). Some are located throughout the NS, while others are more localized. Some are excitatory and some are inhibitory. Some we hear about often because too much or too little of them are often linked to certain diseases such as Parkinson’s Disease (lack of the NT dopamine) and that drugs (legal and otherwise) tend to influence the amount of NT or mimic its action. Finally, for a little perspective, in the functioning human, a neuron will have numerous synapses on its dendrite and cell body (the motor neuron can have 10,000) and can project its influence on a very large number of cells. So let’s take a single neuron (we’ll call her Fred). Whether Fred rises to action and speaks depends on the activities of the hundreds of Fred’s neighbors. If their voices are loud enough, and Fred does rise to action she will in turn be a single voice in a chorus of thousands. But if enough of them are saying the same thing, the results are behaviors (e.g. speaking, thinking, sleeping) that we take for granted. The Central Nervous System The broad goal of this Chapter is to provide a general introduction to the central nervous system (CNS) and its role in communication. It is not meant to be comprehensive and provides only a cursory overview of many important CNS structures. See your text readings for more detailed treatment of the various sections. Unit Organization I. Anatomy of the Nervous System I. II. III. Anatomy of the Nervous System Afferent and Efferent Pathways Centers and Circuits for the Neural Control of Speech A. The Central Nervous System (CNS) i. Basic organizational features of the CNS SPPA 2050 Speech Anatomy and Physiology a. “Slicing and Dicing” the CNS The central nervous system can be divided into the spinal cord and the brain. The brain can be further divided into a number of structures. The following is one way to organize these structures. Spinal cord Brain Hindbrain Medulla Pons Cerebellum Midbrain Forebrain Thalamus Hypothalamus Basal nuclei (ganglia) Cerebrum Further, the brain can be organized along functional lines. For example, later on, we will briefly discuss the limbic system, which is distributed across a number of structures outlined above. You will often read about a structure called the brainstem. This is a term that refers collectively to the medulla, pons and midbrain. b. Histologic organization of the CNS If you inspect the gross appearance of CNS tissue, you will quickly note that that some tissue is white in color and other tissue takes on a grayish color. This is an important distinction since these different tissues contain different kinds of material. White matter contains neuronal fibers that communicate between different parts of the CNS. It is white in appearance because it contains myelin, a fatty substance that insulates the fibers. It is sometimes useful to think of white matter as being analogous to the wiring in a house. Both serve to primarily connect different regions of their respective structures (house or CNS). When you think white matter, think connections between brain regions. In contrast, gray matter contains neuronal cell bodies and synapses that allow communication between neurons. As we learned way back in Unit 1, the cell body and synapses are the primary location of integration and modulation of neural activity. Where the white matter is analogous to the wiring in a house, the gray matter is analogous to the switch panels in a house. Gary matter and switch panels are points where circuits converge and diverge. So, when you think gray matter, think cell bodies and connections between individual neurons. Gray matter is often organized in focal regions of the brain. A focal collection of nerve cells within the CNS is called a nucleus. For example, the cell bodies of the cranial nerves are located in various nuclei within the brainstem. It is important to recognize that damage to either white matter or gray matter can disrupt normal neural function. c. The CNS is symmetric This is simply a reminder for you. As with the rest of the body, the CNS is largely symmetric. Sectioning the CNS along the midsagittal plane will yield two anatomical structures that are roughly mirror images of each other. So, remember, when we discuss a structure like the thalamus, it is important to recall there is a left thalamus and a right thalamus. We will learn in later discussions, certain neural functions are often localized to left or right parts of the CNS. ii. The Cerebrum The cerebrum is the largest part of the human brain. It has a wrinkled surface, which dramatically increases its surface area (works the same way as crumpling a piece of paper you plan to throw in the trash). The cerebral surface is termed the cerebral cortex (bark). The cerebral cortex is largely gray matter that is organized into a series of histologically distinct layers. Increasing the surface area of the cerebrum dramatically increases the number of cell bodies and synapses and hence increases the opportunity for variations in neural connectivity. This wrinkled cortex contains outfoldings and infoldings. An outfolding is termed a gyrus, and an infolding is termed a sulcus. Particularly deep sulci are called fissures. There is sufficient uniformity among brains to prompt anatomists to name many of these gyri and sulci. There are some key fissures that serve to separate the cerebral cortex into smaller parts. The longitudinal fissure separates the cerebral cortex into a left and right hemisphere. Within each hemisphere, there are a number of identifiable lobes. These include the frontal lobe, the parietal lobe, the temporal lobe, the occipital lobe and the limbic lobe. The central sulcus/fissure (Rolandic fissure) separates the frontal and parietal lobes and the lateral SPPA 2050 Speech Anatomy and Physiology sulcus/fissure (Sylvian fissure) separates the temporal lobe from the fontal/parietal lobes. Regions of the cerebral cortex are associated with particular bodily functions. The frontal lobe tends to be involved with motor and executive function. The temporal lobe is principally involved in audition and olfaction. The parietal lobe is involved in general sensation and sensory integration. Finally the occipital lobe in associated with visual function. cerebral cortex. The thalamus is involved in integrating peripheral sensory information of different types with information from other parts of the CNS. Therefore, damage to the thalamus can affect basic sensory and more complex brain functions (such as language formulation). Similar to the basal ganglia, the thalamus consists of a number of distinct nuclei. As was noted earlier, white matter contains axonal fibers that communicate between brain areas. These bundles of fibers are often termed fasciculi. White matter fiber tracts are organized based on the structures between which they communicate. Association fibers connect cortical areas in the same hemisphere. Association fibers can be short (i.e. gyrus to next gyrus) or long (i.e. connecting lobes). Examples of long association fibers are the superior longitudinal fasciculus, which connects the frontal, parietal, occipital, and temporal lobes and the arcuate fasciculus, which connects the frontal, parietal and temporal lobes. Commissural fibers connect hemispheres. Examples include the corpus callosum and the anterior commissure. Finally, projection fibers connect cerebral structures with structures in the brainstem and spinal cord. • Mediodorsal nucleus: also involved in • Anterior nucleus: involved in the brain’s emotional circuits iii. Basal nuclei/ganglia (BG) There is a set of nuclei deep to the cerebral cortex that are collectively termed the basal nuclei or basal ganglia. The nuclei that comprise the BG are (at least) the caudate (taillike) nucleus, the lenticular (lens-like) nucleus, the subthalamic nucleus and the substantia nigra. The lenticular nucleus can be divided into the putamen and globus pallidus. The caudate and putamen are often collectively termed the striatum. The BG is most notably involved in the control of movement. It has been suggested that the BG helps stabilize the motor system. Diseases related to BG function include Parkinson’s Disease (reduced movement) and Huntington’s Disease (uncontrollable “dancing” like movement). iv. Thalamus and Hypothalamus The thalamus is an anatomically and functionally complex structure located deep to the cerebral cortex near the BG. It is often termed the “gateway to the cortex” because all sensory information (exception olfaction) must pass through thalamus before reaching the • • • • emotion Ventral anterior and ventral lateral nuclei: receives input from basal nuclei and cerebellum Ventral posterior lateral (VPL) and ventral posterior medial (VPM): relays information on general body sensation (VPM: sensory from head) Medial geniculate body: processes auditory information Lateral geniculate body: processes visual information The hypothalamus is located inferior to the thalamus (hence “hypo”) and is heavily involved in the controls of autonomic nervous system (ANS) function and endocrine system. The hypothalamus is involved in the release of hormones, controlling food and water intake, regulating sexual behavior, sleep cycles, and emotional responses. It has only an indirect role in speech communication. v. Cerebellum The term cerebellum means “little brain”. Even though it is called the little brain, it contains roughly half the neurons in the entire brain. The cerebellum is principally involved in motor control. It aids in coordinating muscles to produce smooth fluid motion, monitors ongoing “state” of body and makes appropriate adjustments in motor commands in response to that “state”. This structure has also been implicated in learning new motor tasks. Damage to the cerebellum can result in ataxia, a condition characterized by the decomposition of movement, errors in timing and scaling of movement, and tremor during movement activities. The cerebellum is located dorsal to the brainstem and is attached to it by the superior, middle and inferior cerebellar peduncles. The gray matter SPPA 2050 Speech Anatomy and Physiology of the cerebellum is located in a cortex (bark or covering) & in structures referred to as the deep cerebellar nuclei. The cerebellar cortex is the primary information receiving area of the cerebellum. Information (1) about body position (from the brainstem and spinal cord), (2) regarding vestibular status (balance) and (3) from other motor control centers passes to the cerebellar cortex through the inferior and middle cerebellar peduncles. The deep cerebellar nuclei are the primary information sending areas of the cerebellum. Information is sent via the superior cerebellar peduncle to structures such as the thalamus, the brainstem and spinal cord, and the cerebral cortex. the body. Like the rest of the CNS, it contains gray and white matter. The gray matter is a butterfly-shaped (“H”-shaped) area in the middle of the spinal cord. Each “wing” of the butterfly is called a horn. There is a ventral horn which contains the cell bodies of motor neurons, and a dorsal horn, which receives sensory information. The white matter that surrounds the gray matter contain tracts that project (1) to the brain from the body, (2) from the body to the brain and (3) from one segmental level to another segmental level. Tracts from white matter terminate and arise from spinal cord gray matter. It is important to remember that the stretch reflex is mediated within the spinal cord. vi. Limbic System The limbic system is not a single structure, but contains a number of CNS structures including the limbic lobe, the fornix, the hippocampus, the amygdala, the mammilary bodies and the anterior nucleus of thalamus. The limbic system is important for regulating emotional and visceral responses. Its specific role in speech and language is not known, although damage in the area can cause mutism (lack of any speech). B. Peripheral Nervous System vii. Brainstem The brainstem includes the midbrain, the pons and the medulla. It connects the spinal cord with the forebrain. The brainstem performs a variety of functions. It regulates some essential life functions including the maintenance of body temperature, control of respiration and heart rate, swallowing and digestion. The brainstem also contains many nuclei and fiber tracts. These tracts contain fibers that project to/from higher CNS structures from/to spinal cord/brainstem. Nuclei include the cranial nerve nuclei, which contain cell bodies of neurons that comprise the cranial nerves. Damage to the brainstem can be devastating. It can disrupt basic bodily function causing death or an inability to survive without artificial means. Damage to this area can also cause severe motor and sensory impairment of the body. C. Nourishing the CNS viii. Spinal cord The spinal cord is that part of the CNS that runs through the vertebral foramen. The spinal cord is organized length-wise as a series of segments that are quite similar in structure. Each segment is associated with a pair of spinal nerves. There is a tendency for a segment to be associated with function at that particular level of The peripheral nervous system, which includes the cranial and spinal nerves was discussed in Unit 1. However, you need to know the following. You should know all 12 cranial nerves by name and number. In addition, you need to know the motor and sensory function associated with CN V, VII, IX, X, XI, and XII. Refer to the appropriate table. You should also recall there are 31 spinal nerves. i. Vascular Supply to CNS Blood is the means by which the brain is supplied oxygen and nutrients. Although the brain is only 2 % of the total body mass, it uses about 20 % of blood in the body. Glycogen is the energy source for the brain. Unlike other body parts, the brain can’t keep stores of glycogen. Therefore, without blood (and its oxygen and nutrients), the brain quickly becomes “malnourished”. Neural cell death occurs rapidly (within a few minutes). Unfortunately, neurons within the CNS do not routinely regenerate so injuries become permanent. Blood is supplied to the body from the heart via arteries and blood is returned to the heart via veins. In the CNS the term sinus refers to a collection of veins. All arterial blood to the brain come from the internal carotid and vertebral arteries (remember symmetry –there are two of each). The internal carotid artery branches into a middle cerebral artery and an anterior cerebral artery. The middle cerebral artery supplies most of the brain’s lateral surface, major portions of the frontal and temporal lobes (including the insula), the basal ganglia and the SPPA 2050 Speech Anatomy and Physiology thalamus. The anterior cerebral artery supplies the front and medial surface of the brain. The left and right vertebral arteries combine at the midline to form the basilar artery. The basilar artery provides many small arterial branches that supply the brainstem and cerebellum. The basilar artery then splits to form the posterior cerebral artery. The posterior cerebral artery supplies the posterior temporal lobe and the occipital lobe. The left and right anterior cerebral arteries have a small “communicating” artery that connects them. There are also communicating arteries between the left (and right) posterior cerebral artery and the left (and right) internal carotid artery. This complex of shared arterial circulation (or anastamosis) is called the Circle of Willis. There is a real advantage to this somewhat complex anatomical setup (other than to provide good test questions for instructors). A blockage of one or more of the four arteries that bring blood into the brain can occur and yet all areas of the brain can still receive the precious blood they need to survive. Mother nature can be quite clever. responsible for headaches (you take the good with the bad). The middle meningeal layer is the arachnoid membrane. This is a web-like structure that filled with some cerebrospinal fluid. This fluid can serve to provide shock absorption against blows to the head. The inner meningeal layer is called the pia mater. It is very closely associated with the brain surface and follows it rough wrinkled appearance. In the world of anatomy it is common to speak of anatomical “spaces”. Often, these are not actual empty spaces but potential spaces. Like a collapsed balloon, these anatomical spaces are typically empty, but have the potential to be filled with stuff. Between each meningeal layer is a designated space. The space between the skull and dura mater is term the extradural space. The space between dura and arachnoid mater is called the subdural space and the space between arachnoid and pia mater is called the subarachnoid space. These terms are useful to describe clinical conditions (see below). Clinical note Clinical note A blockage, due to thrombosis (clot), embolism (object floating through the bloodstream) or bursting (hemorrhage) of blood vessels in the brain is collectively termed a stroke or cerebrovascular accident (CVA) which can have devastating effects on communication Protecting the CNS Brain tissue is soft and gelatinous. Therefore, it is prone to injury. There need to be ways to protect this critical structure from wear and tear of everyday life. One form of protection comes from the meninges, which are a layered wrapping around the brain and spinal cord. Another form of protection comes from a network of fluid filled cavities inside the brain called the ventricles. The meninges are a layered wrapping of the ii. Protecting the CNS: The meninges The meninges provide an external wrapping to brain and spinal cord. There are three anatomically distinct layers or “mater”. The most superficial layer is a thick tough, fibrous layer called the dura mater. This provides a good solid protection. However, this layer also contains many of the sensory receptors that are Many diseases and disorders associated with the brain can actually be traced to the meninges. Meningitis is an inflammation of the meninges that can result in temporary and/or permanently impaired neurologic function. A meningioma is atumor of the meninges that can invade the brain and cause serious impairment in function. Hematoma (bruising) or hemorrhage (bleeding) can occur into meningeal spaces (subdural, subarachnoid). iii. Protecting the CNS: The ventricles and cerebrospinal fluid Within the brain, there are four interconnecting fluid-filled spaces or ventricles. This fluid is called cerebrospinal fluid (CSF) and it is produced by the choroid plexus within the ventricles. CSF is considered to be principally protective, providing shock absorption. However, CSF may also have some nutritive function. There are two lateral ventricles, which are within each cerebral hemisphere. The lateral ventricles communicate with the third ventricle, which is in the region of the thalamus. The third ventricle communicates with the fourth ventricle via the cerebral aquaduct, which is located ventral to the cerebellum and dorsal to the pons. The fourth ventricle SPPA 2050 Speech Anatomy and Physiology communicates with the central canal, which runs down the center of the spinal cord. (e.g. back). This is yet another example of the relationship between structure and function. Clinical note In order for a sensory stimulus to reach the somatosensory cortex (and thus be perceived), the information must pass over at least three neurons. The first order sensory neuron has its cell body in dorsal root ganglion or cranial nerve nuclei and its axon in a peripheral nerve. It communicates the sensory stimulus to a second order sensory neuron located in either the dorsal gray matter of the spinal cord or a brainstem nucleus. This neuron transmits the sensory information from the spinal cord/brainstem to the contralateral thalamus where it synapses onto a third order sensory neuron. This neuron carries information to the somatosensory cortex via a large fiber bundle called the corona radiata. Note that the information crossed to the contralateral side of the body when it was a second order neuron. An excess CSF is called hydrocephalus (literally, water on the brain), which can occur for a number of reasons and impair normal nervous system function II. Afferent and Efferent Pathways This section provides a very basic introduction to the major afferent (sensory) and efferent (motor) pathways in the CNS. These pathways are critical for conscious and unconscious sensation, voluntary movement, and sensorimotor integration. Before proceeding, it is important to point out that at the level of the cerebral cortex (and most subcortical structures), bodily representation is “crossed”. That means that the sinistral side of the body is represented in the dextral cortex and vice versa. For this to occur, afferent and efferent pathways must cross the midline (decussate) of the CNS. The term contralateral refers to the opposite side and the term ipsilateral refers to the same side. For example, we might say that the left cerebral cortex controls the contralateral side of the body. We might also say that the left vagus nerve provides the motor supply to the ipsilateral intrinsic muscles of the larynx. i. Sensory Pathways The primary cortical representation of somatosensory (somato – body) function is located in a long strip of cortex just posterior to the central sulcus. This structure is called the post-central gyrus, but is also referred to as the sensory strip, the primary sensory cortex or the somatosensory cortex. In terms of the cortical map of Brodmann, this areas is assocated with Brodmann’s Areas 1, 2, 3 and 5. We say that the somatosensory cortex is somatotopically organized. This means that the bodily location of sensory information is preserved within the cortex. There is a representation of the whole body along the length of the somatorsensory strip. Further, some body parts that we recognize to be highly sensitive (e.g. lips, tongue, fingers) have larger cortical representation than less sensitive parts ii. Motor Pathways The primary cortical representation of movement is located in a long strip of cortex just anterior to the central sulcus. This structure is called the pre-central gyrus, but is also referred to as the motor strip, the primary motor cortex or the motor cortex. In terms of the cortical map of Brodmann, this areas is assocated with Brodmann’s Area 4. Analogous to the somatosensory cortex, body movements are highly localized and there is a map of body parts in this strip of cortex. We often say that the primary motor and sensory cortices contains homunculi or “little men”. Those structures that require more motor precision (e.g. tongue and fingers) have larger cortical representations than muscles systems that produce more gross movements (e.g. trunk muscles). Communicating a motor command to the body can involve as few as two neurons. The upper motorneuron (UMN) has its cell body in the motor cortex. Its axonal fibers descend from the cortex through the internal capsule (white matter that contains many fibers going to and from the cortex) and then through the brainstem. At this point most of these fiber cross over (decussate) to the contralateral side of the body. The UMN then synapses onto a lower motorneuron (LMN), whose cell body is located either in cranial nerve nuclei in the brainstem (head and neck muscles) or in the SPPA 2050 Speech Anatomy and Physiology ventral horn of the spinal cord (below the head). The LMN leaves the CNS and synapses onto the muscle fiber. Because there are no intervening neurons between the LMN and the muscle, the LMN is sometimes called the final common pathway. Any instruction for a muscle to move must pass through this neuron. This part of the motor system is sometimes called the direct motor system because it contains so few synapses. It is also called the pyramidal motor system because the white matter tracts through which the UMN run are pyramidal shaped when they pass through the medulla. Clinical Note Damage to upper motorneuron and lower motorneuron result in quite different clinical syndromes. UMN damage can result in spastic paralysis, increased muscle tone and exaggerated reflexes. In contrast, damage to LMN can result in flaccid paralysis, absent or reduced reflexes, decreased muscle tone and atrophy of muscle. Extrapyramidal (Indirect) Motor System The pyramidal motor system is associated with voluntary (willful) movement. It includes primary motor cortex (60 %) and other cortical structures including the premotor (anterior to motor strip) and sensory cortices (40%). However, this is not the only neural system that controls movement. As mentioned earlier there are other neural structures (e.g. the cerebellum and the basal nuclei) that a involved in regulating movement. It is common in neurology to use the term extrapyramidal or indirect motor system to refer to a system that influences and regulates the motor instructions that are sent to the periphery. The basal nuclei/ganglia is the principle structure in this system. While voluntary movement is initiated by the direct system, this indirect system is crucial to the production of stabile, skillfully executed movements. As we learned earlier, damage to this system can seriously disrupt voluntary movement control. Sensorimotor Regulation Although we discuss sensory and motor function separately, it is important to recognize these systems are inextricably linked. For example, when we move our bodies, sensations are generated (recall the muscle spindle). We make use of this sensory information to evaluate whether our movements are reaching their intended targets and make adjustments if necessary. Speech production stimulates a number of sensory modalities including touch, kinesthesia/proprioception, and audition. Although short term disruptions to sensory systems do tend not to interfere with speech production, longer term sensory loss (i.e. hearing loss) can result in deterioration of speech motor skills. III. Centers and Circuits for the Neural Control of Speech It is often suggested that high level neural functions are “localized” to a region of the brain. For example, language is predominantly represented in the dominant (i.e. left) hemisphere. This argument could be extended to suggest that there are very specific anatomical locations for the range of language and cognitive functions. However, this does not seem to be the case. Although specific brain structures appear critical for speech and language, no single structure “houses” all the machinery needed. For example, stimulating some portion of the brain can result in events necessary for speech (e.g. vocalization), but not “full-blown” speech. It appears that speech and language functions are distributed across many structures that include, but are not limited to Broca’s area Broca’s Area is located anterior to the primary motor area (the face and mouth area) on or near the inferior frontal gyrus of the dominant hemisphere (usually left). This area is associated with planning speech production and damage in this region can result in an expressive or Broca’s aphasia (loss of language) that compromises an individuals ability to express themselves using language, while preserving an ability to understand others. Supplementary motor area The supplementary motor area (SMA) appears to be involved in planning motor sequences including preparation of movement, performing “internally” generated motor plans and is implicated in the planning of propositional speech. Wernicke’s area Wernicke’s area is important for the understanding and formulation of language. Damage in this area often results in an aphasia SPPA 2050 Speech Anatomy and Physiology with problems understanding or repeating language (hence the term receptive or Wernicke’s aphasia). With deficits of this type, patients exhibit spoken language that while fluent, makes little sense Other important areas Primary motor cortex (“speech” muscles) Basal nuclei Cerebellum Thalamus Somatosensory cortex Primary & association areas of auditory cortex