Supplementary Data (doc 1467K)
advertisement
")
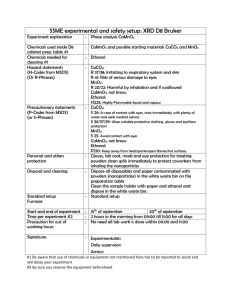
Supplementary Data to Ozburn et al. NPP Submission Methods and Materials Drugs. Ethanol (Aaper, Shelbyville, KT, USA), ketamine hydrochloride (Fort Dodge Animal Health, Fort Dodge, IA, USA), xylazine hydrochloride (Akom Inc., Decator, IL, USA) or pentobarbital sodium (Ovation Pharmaceuticals Inc., Deerfield, IL, USA) were dissolved in 0.9% saline and injected intraperitoneally (i.p.) in a volume of 0.2 ml per 10 g of body weight. Continuous Access Two-bottle Choice. Ethanol - As described in Ozburn et al. (2010), male mice were habituated to individual housing and sipper bottles for one week prior to the start of the experiment. Mice were offered water and 3% ethanol (v/v in tap water) for 2 days. After 3% ethanol, escalating concentrations (up to 21%) were offered versus water, 2 days each. Fluid intake was measured daily. Mice were weighed every four days. Ethanol preference (mL ethanol solution consumed/mL total fluid consumed), ethanol consumption (g pure ethanol/kg body weight/day), and total fluid consumption were measured. n=9/genotype. Quinine - As described in Blednov et al. (2010), male and female mice were habituated to individual housing and sipper bottles for one week prior to the start of the experiment. Mice were serially offered offered 0.03 and 0.06 mM quinine hemisulfate (in tap water) for 4 days each. Fluid intake was measured daily. Mice were weighed every four days. Quinine preference (mL quinine solution consumed/mL total fluid consumed) was measured. n=9-11/sex/genotype. Conditioned Taste Aversion. As described in Ozburn et al. (2010), mice were adapted to a water-restriction schedule (2 hr water/day; ZT2-4) over 7 days. At 48 hr intervals over the next 12 days, mice received 1 hr access to a solution of saccharin (0.15% wt/vol) at ZT2. After 1 hr access to saccharin, mice received injections of saline or ethanol (2 g/kg). All mice received 1 hr access to tap water 4 hr after injections to prevent dehydration. 2 hr access to tap water was given during intervening days. n=12/group/genotype. Acute Functional Tolerance. Testing was carried out similar to the protocol described by Erwin and Dietrich (1996). At ZT6, mice were placed on a rotarod (fixed 5 RPM) and required to stay on rotarod for 1 minute before testing. Mice were then injected with ethanol (1.75 g/kg) and placed on rotarod every 5 minutes until they could stay on rotarod for 1 minute. At this time, retro-orbital blood samples were collected (BEC1), and mice were injected with ethanol (1.75 g/kg) a second time and then placed on the rotarod every 5 minutes until they could stay on rotarod for 1 minute. Upon the second rotarod recovery, another blood sample was taken (BEC2). n=6/genotype. Ethanol Clearance. As described in Ozburn et al. (2010), mice were injected with ethanol (4 g/kg) at ZT 4 or ZT16 and rates of ethanol clearance were determined using a spectrophotometric enzyme assay. Blood samples (50 uL) were taken from retro-orbital sinus (at 30, 60, 120, 180, and 240 min post injection), added to 2 mL 3% perchloric acid, and centrifuged for 10 min at 1000 x g. Resulting supernatants were used to determine blood ethanol concentration (BEC) using an alcohol dehydrogenase enzyme assay. n=3-4/ZT/genotype. Statistical Analysis. All data are expressed as mean ± SEM. Significance for the acute functional tolerance assay was determined by Student’s t-test. Limited access drinking data, CPP, CTA, and ethanol metabolism data were analyzed by two-way analysis of variance (ANOVA). Repeated measures were applied where appropriate. In all experiments, p < 0.05 is considered significant. Results Continuous Access Two-bottle Choice. Ethanol – Male ClockΔ19 exhibit a strong trend toward increased ethanol preference. To determine if functional CLOCK is important for voluntary ethanol drinking in male mice, we measured ethanol preference and consumption in ClockΔ19 and WT littermates using the continuous access two-bottle choice paradigm. ClockΔ19 male mice exhibited no difference in ethanol intake and a strong trend for increased ethanol preference that was accompanied by reduced total fluid consumption (Fig. 1a,b,c; Ethanol consumption: genotype x concentration interaction – F(6,96)=0.63, p=0.71, main effect of genotype – F(1,16)=0.96, p=0.34, main effect of concentration – F(6,96)=5.4, p<0.0001; Ethanol preference: genotype x concentration – F(6,96)=0.61, p=0.27, main effect of genotype – F(1,16)=3.86, p=0.06, main effect of concentration – F(6,96)=7.35, p<0.0001; Total Intake: genotype x concentration – F(6,96)=1.21, p=0.30, main effect of genotype – F(1,16)=15.19, p<0.001, main effect of concentration – F(6,96)=5.59, p<0.0001). Quinine – ClockΔ19 exhibit similar quinine preference. Preference for non-alcohol tastants are important determine for mutant and WT comparisons and can relate to ethanol preference. For instance, some studies have revealed a positive correlation between preference for sweet solutions and alcohol in mice (Blednov et al., 2012), while others have reported no relationship (Blednov et al., 2007). It has been previously shown that ClockΔ19 mice exhibit a higher preference for the sweet tastant, sucrose (6% average increase in preference; Roybal et al. 2005). To determine if functional CLOCK is also important for the bitter tastant preference, we measured quinine preference in ClockΔ19 and WT littermates using the continuous access two-bottle choice paradigm. ClockΔ19 and WT mice exhibited similar quinine preference and both genotypes exhibited decreased preference at the higher quinine concentration (Fig. 2a,b; Females - genotype x concentration interaction – n/s, main effect of genotype – n/s, main effect of concentration – F(1,18)=19.56, p<0.001; Males genotype x concentration interaction – n/s, main effect of genotype – n/s, main effect of concentration – F(1,18)=19.99, p<0.001). ClockΔ19 and WT littermates display similar ethanol aversion, acute functional tolerance, and clearance rates. We have previously shown that Clock has a role in the neurobiological processes involved in both the behavioral response to cocaine and the incentive motivation for cocaine, prompting a more rigorous examination of ethanol responses and related phenotypes in ClockΔ19 and WT littermates. Differences in ethanol intake can be due to differences in the aversive properties of ethanol. Ethanol-induced conditioned taste aversion (CTA) is often used as a measure of the aversive properties of drugs of abuse. CTA strongly and negatively correlates with ethanol intake (Green and Grahame, 2007). To examine the possibility that differences in ethanol intake could be due to differential responses to the aversive effects of ethanol, we measured ethanol-induced CTA and hypothesized reduced CTA in ClockΔ19 mice. ClockΔ19 and WT mice expressed CTA to a similar extent suggesting a similar sensitivity to the aversive properties of ethanol (Fig. 4a; main effect of treatment – F(1,44)=52.05, p<0.0001). Increased ethanol drinking could be due to increased development of acute functional tolerance, as well as increased ethanol clearance. However, ClockΔ19 and WT mice acquire similar acute functional tolerance and exhibit similar rates of ethanol clearance at both times of day tested (Fig. 4b,c). Since ClockΔ19 mice exhibited increased ethanol intake not accompanied by obvious correlations with other ethanol-related behaviors, we further examined their pharmacological responses to other drugs of abuse with sedative properties. Supplemental Figures and Legends Supplemental Figure 1. Male Clock∆19 mice exhibit a strong trend toward increased ethanol preference. (A) Ethanol consumption (B) Ethanol preference (C) Total fluid intake. Supplemental Figure 2. Clock∆19 and WT mice exhibit a similar taste aversion to quinine. (A) Female quinine solution preference. (B) Male quinine solution preference. Supplemental Figure 3. Clock∆19 and WT mice exhibit similar behavioral and metabolic responses to ethanol. (A) Ethanol-induced conditioned taste aversion to saccharin (B) Ethanol-induced acute functional tolerance (C) Rates of ethanol clearance. References Blednov YA, Mayfield RD, Belknap J, Harris RA. (2012) Behavioral actions of alcohol: phenotypic relations from multivariate analysis of mutant mouse data. Genes Brain Behav. 11:424-435. Blednov YA, Cravatt BF, Boehm SL 2nd, Walker D, Harris RA (2007) Role of endocannabinoids in alcohol consumption and intoxication: studies of mice lacking fatty acid amide hydrolase. Neuropsychopharmacology 32:1570-1582. Blednov YA, Ozburn AR, Walker D, Ahmed S, Belknap JK, Harris RA (2010) Hybrid mice as genetic models of high alcohol consumption. Behav Genet 40:93110. Erwin VG, Deitrich RA (1996) Genetic selection and characterization of mouse lines for acute functional tolerance to ethanol. J Pharmacol Exp Ther 279:13101317. Green AS, Grahame NJ (2007) Ethanol drinking in rodents: is free-choice drinking related to the reinforcing effects of ethanol? Alcohol 42:1-11. McClung CA, Sidiropoulou K, Vitaterna M, Takahashi JS, White FJ, Cooper DC, Nestler EJ (2005) Regulation of dopaminergic transmission and cocaine reward by the Clock gene. Proc Natl Acad Sci USA 102:9377-9381. Ozburn AR, Harris RA, Blednov YA. (2010) Behavioral differences between C57BL/6J x FVB/NJ and C57BL/6J x NZB/B1NJ F1 hybrid mice: relation to control of ethanol intake. Behav Genet 40:551-563. Ozburn AR, Larson EB, Self DW, McClung CA (2012) Cocaine selfadministration behaviors in ClockΔ19 mice. Psychopharmacology 223:169-177.