AN2501 Extended Essay Derek Scott
advertisement

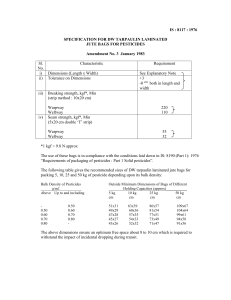
AN2501 Extended Essay Derek Scott “Discuss the possible roles of human keratinocyte growth factor in lung development.” Essay target was 1000 ± 100 words Essay was written by a student at level 2 of undergraduate degree. Student did not have access to electronic search engines to find journal articles. Several recent scientific studies have shown that keratinocyte growth factor (hKGF) plays an important role in lung repair and, obviously, development. Most of these studies have been conducted on continuous mouse, rabbit, rat or lung keratinocyte cell lines in the hope that, by learning more about the effects of KGF on the growth and development of the lungs, we may be able to develop new therapeutic or diagnostic methods of dealing with lung disease or malfunction. Keratinocyte growth factor (KGF) KGF is a member of the fibroblast growth factor (FGF) family, and is also known as FGF-7 (Finch et al., 1995; Housely et al., 1994). It is only one of several growth factors that have been identified as acting in lung growth and repair (see Table 1 and Figure 1), and it does so by affecting keratinocyte proliferation in a paracrine manner (Perkett, 1995; Marchese et al., 1990). KGF was first identified as a highly specific keratinocyte mitogen after its isolation from a rat lung stromal fibroblast line (Housely et al., 1994; Wilson et al., 1993). It is recognised by KGF receptors on certain epithelial cells (Marchese et al., 1990; Rubin et al., 1989). KGF is not only found in the lungs, but influences the proliferation and differentiation patterns of epithelial cells in the skin, gut and reproductive tracts also. The gut, in particular, responds to and synthesises KGF (Housely et al., 1994; Marchese et al., 1990). KGF also has a signal peptide for secretion (Perkett, 1995). Table 1. Selection of the growth factors involved in lung repair. This table depicts the wide variety of potential regulators of lung repair. The wide variety illustrates the difficulty in working out which growth factors might be the most important in lung growth and repair (Table taken from Perkett, 1995). Figure 1. Stages of lung injury and repair, with some of the potential growth factors involved. The cascade sequence which occurs in the lung during establishment of normal architecture. FGF – fibroblast growth factor, HGF – hepatocyte growth factor, IGF – insulin-like growth factor, KGF – keratinocyte growth factor, PDGF – platelet-derived growth factor, TGF – transforming growth factor, VEGF – vascular endothelial growth factor. (Image taken from Perkett, 1995). Role of Keratinocytes In the lung itself, there are two main kinds of keratinocytes (pneumocyte) – Type I and Type II. It is the Type II pneumocytes which are the important cells in lung development and also in recovery after injury since they proliferate and differentiate into Type I cells. These Type II cells are mitotically active and cuboidal in shape, in comparison to the flat, metabolically-inactive Type I cells which form about 95% of lung architecture. Type II cells are usually found in the corners of alveoli, and secrete pulmonary surfactant. This surfactant is a mixture of proteins and lipids that lowers alveolar surface tension, thus stabilising alveolar structure (see Figure 2). Type II cells are very prominent in foetal lungs and are very likely to be responsible for the developmental changes which occur in them. Without the proper development of Type II cells, the foetus cannot form functional lungs or surfactant, preventing its safe transition, preventing its safe transition from an amniotic to an external environment (Ulich et al., 1994; Panos et al., 1993 & 1995). The differentiation of Type II to Type I cells allows the establishment of normal alveolar parenchymal architecture since they line alveolar septae as rows of columnar cells (Yi et al., 1995; Ulich et al., 1994) (see Figure 3). It is thought that not only KGF, but glucocorticoids, oestrogen, thyroid hormones, bombesin and EGF promote and enhance foetal lung maturation and surfactant production in vivo. KGF is also important in the re-establishment of lung structure after injury since it prevents fibrosis of tissue (Perkett, 1995). An overdose of KGF, however, may cause fibrosis. KGF induces an up-regulated expression of the protein v6 integrin in developing tissues and at wound sites. This affects cell spreading, migration, and growth during reorganisation of epithelia in development, repair and neoplasia (Breuss et al., 1995). Epithelial-mesenchymal interactions mediate aspects of normal lung development and growth during establishment of normal alveolar architecture (Panos et al., 1993). It must be remembered that lung growth and repair requires a coordinated cascade of events with growth factors being the key regulators (Ulich et al., 1994) (see Figure 1). Effects of KGF on Pneumocytes Ulich et al. (1994) tried to examine the in vivo effects of KGF by injecting hKGF intratracheally in rats. Three days after injection, the Type II cells in the lungs exhibited hyperplasia which appeared histologically similar to healing epithelium in lungs. By day 6, the epithelium had returned to normal (see Figure 4). Adult rat lungs expressed mRNA for KGF and its receptor, indicating that KGF is involved in epithelial maintenance. The effects of KGF were doseand time-dependent. The results of histological sections were confirmed by immunohistochemical marking of Surfactant protein B (SP-B). SP-B is produced in hyperplastic alveolar lining cells. Lamellar inclusions were also seen, features which are characteristic of surfactant-secreting Type II cells. KGF did not seem to have any effect on the larger, conducting airways. The work of Panos et al. (1995) established that intratracheal administration of recombinant hKGF stimulated Type II cell proliferation in vivo, and could reduce the effects of hyperoxia- induced injury in rats. Immunohistochemical analysis was used to determine intrapulmonary distribution and cellular localisation of KGF instilled into the rats’ tracheas. Six hours after administration, KGF was detected in lung parenchyma and along alveolar epithelial cell membranes. By 18-24 hours, KGF was detected intracellularly in alveolar epithelial cells and intra-alveolar macrophages. KGF was not found after 48 hours or in any sections taken from the lung tissue. Intra-tracheal instillation of 5 mg/kg KGF stimulated a marked, time-dependent increase in Type II cells that were labelled (see Figure 5). The increase in cell proliferation was documented by flow cytometric analysis of isolated Type II cells, which revealed a 5-fold increase in the number of cells which were in the S and G2/M phases of the cell cycle. Rats were also treated with KGF and placed in a hyperoxic environment to test if KGF affected lung injury responses. Animals treated with 1-5 mg/kg showed much lower mortality rates. Lower doses of KGF gave the same results as those found with rats that were untreated or who were given heatdenatured KGF (see Figure 6). The treated rats which survived the hyperoxia had minimal haemorrhage and no exudate in the intraalveolar space. The work of Rannels & Rannels in 1988 had already given the background to other more recent studies that Type II cells have a well-known proliferative response after exposure to insults such as high concentrations of oxygen or nitrogen dioxide. Typically, the cells would enlarge, divide and spread to re-epithelialize an alveolar basement membrane denuded by loss of the Type I cells. In rats, it only took 45 days after lung bilobectomy for components of parenchymal architecture (including capillary and alveolar surface area, volumes and cell populations) to be restored to normal. Everett et al. (1990) also demonstrated that Type II cells (and therefore KGF) played an integral role in lung architecture maintenance by showing that hyperoxia and bacterial infection were potent stimuli of alveolar epithelial and interstitial hyperplasia. They saw decreased surfactant production in human lung injury patients also. Thus, in certain lung injuries that include respiratory failure, cell cycle kinetics are altered and, possibly, cellular phenotypes are changed. KGF is obviously extremely important in lung growth and development, but whether it will actually be useful in promoting human lung development or repair when given artificially to humans is not yet known, since it has not yet undergone clinical trials. Much is still to be understood about the control of KGF bioactivity (e.g. receptors, binding proteins, interactions etc.) (Panos et al., 1993; Finch et al., 1989; Ulich et al., 1994). Directed delivery of KGF into the lungs may be a form of therapy whereby alveolar epithelium could be preserved/restored during exposure to hyperoxia or other injurious agents. (Panos et al., 1995). KGF might facilitate reepithelialisation of the airways or assist in the survival of babies born with respiratory distress syndrome, where their ability to produce pulmonary surfactant is compromised. From experimental evidence (Yi et al., 1995), it is thought that there would be little chance of permanent, deleterious side-effects due to treatment with KGF, since its actions are stopped or even reversed after cessation of treatment. Overdoses could even be reversed by targeting KGF with monoclonal antibodies (Sugahara et al., 1995). In summary, a better understanding of KGF and other factors will allow us to understand our own lung development more clearly, as well as allowing us to conceive therapeutic interventions that could prevent some chronic lung diseases (Perkett, 1995). Reference List Breuss, J.M, Gallo, J., DeLisser, H.M., Klimanskaya, I.V., Folkesson, H.G., Pittet, G.F., Nishimura, S.L., Aldape, K., Landers, D.V., Carpenter, W. et al. (1995). Expression of the beta 6 integrin subunit in development, neoplasia and tissue repair suggests a role in epithelial remodeling. Journal of Cell Science, 108(6), 2241-51. Everett, M.M., King, R.J., Jones, M.B., Martin, H.M. (1990). Lung fibroblasts from animals breathing 100% oxygen produce growth factors for alveolar type II cells. American Journal of Physiology, 259, L247-54. Finch, P.W., Rubin, J.S., Miki, T., Ron, D., Aaronson, S.A. (1989). Human KGF is FGF-related with properties of a paracrine effector of epithelial cell growth. Science, 245, 752-55. Housely, R.M., Morris, C.F., Boyle, W., Ring, B., Blitz, R., Tarpely, J.E., Aukerman, S.L., Devine, P.L., Whitehead, R.H., Pierce, J.F. (1994). Keratinocyte growth factor induces proliferation of hepatocytes and epithelial cells throughout the rat gastrointestinal tract. Journal of Clinical Investigation, 94(5), 1764-77. Marchese, C., Rubin, J., Ron, D., Faggoni, A., Torrisi, M.R., Messina, A., Frati, L., Aaronson, S.A. (1990). Human keratinocyte growth factor activity on proliferation and differentiation of human keratinocytes: differentiation response distinguishes KGF from EGF family. Journal of Cellular Physiology, 44, 326-32. Panos, R.J., Rubin, J.S., Csaky, K.G., Aaronson, S.A., Mason, R.J. (1993) Keratinocyte growth factor and hepatocyte growth factor/scatter factor are heparin-binding growth factors for alveolar type II cells in fibroblast-conditioned medium. Journal of Clinical Investigation, 92(2), 969-77. Panos, R.J., Bak, P.M., Simonet, W.S., Rubin, J.S., Smith, L.J. (1995) Intratracheal instillation of keratinocyte growth factor decreases hyperoxiainduced mortality in rats. Journal of Clinical Investigation, 96(6), 2026-33. Perkett, E.A. (1995) Role of growth factors in lung repair and diseases. Current Opinion in Paediatrics, 7(3), 241-49. Rannels, S.R. & Rannels, D.E. (1988). Alterations in type II pneumocytes cultured after partial pneumonectomy. American Journal of Physiology, 254, C684-90. Rubin, J.S., Osada, H., Finch, P.W., Taylor, W.G., Rudikoff, S., Aaronson, S.A. (1989) Purification and characterization of a newly identified growth factor specific for epithelial cells. Proceedings of the National Academy of the USA, 86, 802-6. Sugahara, K., Rubin, J.S., Mason, R.J., Aronsen, E.L., Shannon, J.M. (1995) Keratinocyte growth factor increases mRNAs for SP-A and SP-B in adult rat alveolar type II cells in culture. American Journal of Physiology, 269(3 Pt 1), L344-50. Ulich, T.R., Yi, E.S., Longmuir, K., Yin, S., Blitz, R., Morris, C.F., Housely, R.M., Pierce, G.F. (1994) Keratinocyte growth factor is a growth factor for type II pneumocytes in vivo. Journal of Clinical Investigation, 93(3), 1298-306. Wilson, S.E., Walker, J.W., Chang, E.L., He, Y.G. (1993) Hepatocyte growth factor, keratinocyte growth factor, their receptors, fibroblast growth factor receptor-2, and the cells of the cornea. Investigative Opthalmology and Visual Science, 34(8), 2544-61. Yi, E.S., Shabaik, A.S., Lacey, D.L., Bedoya, A.A., Yin, S., Housely, R.M. (1995) Keratinocyte growth factor causes proliferation of urothelium in vivo. Journal of Urology, 154(4), 1566-70.