Supplementary Information (doc 92K)
advertisement
")
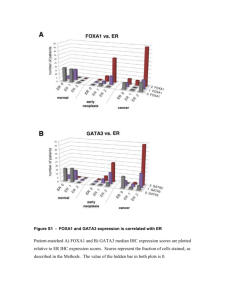
Supplementary Information GATA3 Inhibits Lysyl Oxidase Mediated Metastases of Human Basal Triple-Negative Breast Cancer Cells Isabel M. Chu, Aleksandra M. Michalowski, Mark Hoenerhoff, Kornelia M. Szauter, Dror Luger, Misako Sato, Kathy Flanders, Akira Oshima, Katalin Csiszar and Jeffrey E. Green Supplementary Materials and Methods Cells, transfection and lentivirus infection. Cells were maintained in DMEM/high glucose (Invitrogen, Carlsbad, CA, USA), supplemented with 10% fetal bovine serum (FBS) (Invitrogen), penicillin/streptomycin (Invitrogen) and sodium pyruvate (Invitrogen). BT474 cells were transfected with siGATA3#2 AACATCGACGGTCAAGGCAAC or siGATA3#3f: AAGCCAAGCGAAGGCTGTCT using Dharmafec™ according to the manufacturer’s instruction (Dharmacon, Lafayette, CO, USA). Lentiviruses expressing FUGW-FerH-IRESeGFP, FUGW-FerH-GATA3-IRES-eGFP, FUGW-FerH-IRES-DsRedExp2 and FUGW-FerHLOX-FLAG-IRES-DsRedExp2 were produced using gateway cloning technology and expressed following the procedures described previously (Hoenerhoff et al., 2009). GATA3 and LOX cDNAs were purchased from Open Biosystems (clone ID 3450299 and 30915233, respectively). Cells were selected in G418 (Invitrogen) and sorted by fluorescence-activated cell sorting for high GFP and/or DsRedExp2 expression. Methylation-specific PCR. DNA was isolated from stably transduced 231-Empty and 231GATA3 cells using the DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA, USA) according to 1 the manufacturer’s protocol. DNA was sulfonated using the EZ DNA MethylationTM Kit (ZYMO Research). Methylation-specific PCR was performed with the following primer sets: 5'AAGTTAGTGTGTTTTAGGATGTGTGT-3' and 5'-CTTCCCTTTCCCCTTTCTCAAT-3' for LOX template amplification (320 bp), 5'-GAATAAATAGTTGAGGGGCGGTC-3' and 5'GCGACAATCCCGAAAAACG-3' for methylated LOX (122 bp); 5'- TGTGAATAAATAGTTGAGGGGTGGTT-3' and 5'-CCACACAACAATCCCAAAAAACA-3' for unmethylated LOX (129 bp) (Kaneda et al., 2004). Bisulfite-treated DNA was PCR- amplified with annealing temperatures of 610 C, 650 C and 590 C for LOX template, methylated and unmethylated LOX DNA, respectively. Mice, necropsy and ex-vivo imaging. Mice were injected in the mammary fat pad #2 or #7 with 2 x 106 cells. Tumor growth was measured twice weekly by caliper. Mice were sacrificed once tumors reached 2 cm in diameter or when mice became clinically ill. For experimental metastasis studies, mice were injected with 1 106 cells via tail vein and were sacrificed after 8 weeks or when they became clinically ill. Primary tumors and/or lungs were collected, fixed in 4% paraformaldehyde, processed into paraffin blocks, sectioned and stained with haematoxylin and eosin (H&E). Lungs from tail-vein injected mice were inflated with 1 x PBS and analyzed by fluorescent whole organ microscopy imaging (Leica DM IRB) as previously described (Barkan et al., 2008; Barkan et al., 2010). Fluorescent GFP signal was measured at 10 magnification and analyzed using OpenLab software (Open Biosystems, Huntsville, AL, USA) to measure the surface area of the metastases, examining at least 20 fields per mouse, and expressed as total pixels per mouse lung. Quantification of TUNEL and Ki-67 staining from primary tumors and lung lesions. Immunohistocemically-stained sections (Ki-67 and TUNEL Assay) of mouse lung were analyzed 2 for positive brown staining and negative blue staining by image analysis (IA) techniques. Slides were digitally scanned on the Scanscope™ XT (Aperio Technologies, Vista, CA). Quantitative Reverse-Transcription PCR (Q-RT-PCR). cDNA was synthesized from 5 µg of total RNA using the Superscript II kit (Invitrogen). Q-RT-PCR was performed using IQ SYBR Green Supermix (Bio-Rad Laboratories, Hercules, CA, USA) and an iCycler Thermal Cycler (Bio-Rad Laboratories). The quantity of mRNA was normalized to the housekeeping gene cyclophilin B. LOX activity. LOX activity was measured as the fluorometric b-Aminopropionitrile (BAPN) inhibitable LOX activity assay using Amplex red (Palamakumbura et al., 2002). Cells were cultured in complete media until confluent at which time they were switched to serum free, phenol red free DMEM. LOX activity was measured as the fluorometric BAPN inhibitable LOX activity assay using Amplex red (Palamakumbura et al., 2002), that we have adapted for the 96well plate (Fogelgren et al., 2005). Briefly, three days post-confluent cultured media samples were concentrated using Amicon Centrifugal Filter Devices (Millipore). Triplicates of 100 g protein samples were added to the reaction mix consisting of 50 mM sodium borate (pH 8.2) (Sigma), 1.2 M urea, 10 mM 1,5-diaminopentane (cadaverine, Sigma) substrate, 50 M Amplex Red (Invitrogen) and 0.02 U horseradish peroxidase (HRP, Sigma). The protein samples were incubated at 37C in the presence or absence of 500 M BAPN and measured using a Polarstar Optima fluorometer (BMG Labtechnologies). The fluorescent product was excited at 560 nm and the emission was read at 590 nm every 5 minutes for 2 hours. All samples were assayed at least in triplicate. LOX activity was calculated as the increase in fluorescence above that of BAPN containing controls at the 2400 second (40 min) time point. 3 Flow analysis for myeloid cells. Ex-vivo immune cell preparation for FACS analysis was performed as previously described (Luger et al., 2008). Briefly, lungs were crushed and filtered through a 40 mm strainer in media to obtain single cell suspensions. Red blood cells were lysed using ACK and cells were labeled with fluorochrom CD45 (PE), CD11b (APC), Gr1 (FITC) and F4/80 (APC) (eBiosience, San Diego, CA, USA) antibodies. Immune cells were gated for CD45 + and then analysis of the other markers were based on this population. Samples were analyzed using FACS Caliber (BD). ELISA Assays. Equal cell numbers were seeded in 5% FBS culture media and media was collected after 2 days for analysis. Analytes were assayed by SearchLight Sample Testing Services (Aushon Biosystems). Each assay was performed in a 96-well custom arrayed plate with target specific anti-human antibodies. Microarray data processing. Total RNA was isolated by Trizol® (Invitrogen). RNA quality was checked on an Agilent Bioanalyzer (Agilent, Santa Clara, CA, USA). All samples used for microarray analysis had a high quality score (RIN >9). RNA (1 mg) was reverse transcribed with a T7-oligo(dT) primer and biotin labeled using the Affymetrix One Cycle Target Labeling kit (Affymetrix) following the manufacturer’s protocol. Three replicates of each group were prepared, labeled, and hybridized to Affymetrix human U133 plus 2.0 GeneChips and scanned on an Affymetrix GeneChip scanner 3000. Data were collected using Affymetrix GCOS software and processed into log base 2 gene expression measures using the gcRMA algorithm and quantile normalization (Wu et al., 2004) using BRB-ArrayTools developed by Richard Simon and the BRB-ArrayTools Development ArrayTools.html). 4 Team (http://linus.nci.nih.gov/BRB- Public microarray data sets. Retrospective studies were carried out with population based cohort of 295 breast cancer patients obtained from Van de Vijver (van de Vijver et al., 2002)and with the set of 51 breast cancer cell lines from Neve and colleagues (Neve et al., 2006). Prior to analyses each gene expression was standardized within a data set by means of Z-score transformation. Statistical analyses. Proliferation assays were analyzed by two-way repeated-measures ANOVA using Bonferroni post-tests to determine significant differences on individual days in GraphPad Prism version 5.01 (www.graphpad.com). GFP immunofluorescence and Q-RT-PCR mean differences were tested with an unpaired Student's t-test. Statistical significance was depicted in the figures as *P<0.05, **p<0.01 and ***p<0.001. Differentially expressed genes between 231-Empty and 231-GATA3 cells were identified with univariate unpaired Student’s ttest and p-value threshold of 0.001, using the BRB-ArrayTools statistical software. Retrospective studies employed two-way hierarchical clustering (1-correlation distance and average linkage) and heatmap display to analyze the microarray gene expression profiles. Oneway ANOVA planned comparisons were applied to estimate differences in LOX and GATA3 expression among breast cancer subtypes adopting the previous classification of the 295 cancer patients (van de Vijver et al., 2002) and 51 breast cancer cell lines (Neve et al., 2006). Pearson’s correlation coefficient was applied to determine the association between expression of GATA3 and LOX in the cancer patients and cell lines. In the survival analyses, gene expression of LOX and GATA3 was dichotomized into low and high levels of expression (lower or higher than the mean) and the relationship of LOX and GATA3 expression and 10-year survival was initially analyzed by Cox proportional-hazards regression including an interaction effect. After initially identifying a significant interaction 5 between the expression of GATA3 and LOX on patient survival, a further analysis was carried out by classifying patients into groups expressing low and high levels of GATA3 (below the mean and above the mean of all patients, respectively) and further classifying the patients into groups expressing low and high levels of LOX (below and above the mean within each GATA3 stratum). Survival curves were estimated using the Kaplan-Meier method and the long-rank test was used to test the null hypothesis of no difference in survival between the groups of low and high levels of gene expression. Survival curves were estimated using the Kaplan-Meier method and the long-rank test for patients stratified by the GATA3 levels. The retrospective analyses were performed with R statistical programming and the stats and survival packages (R version 2.8.1). 6 Supplementary Figure Legends Supplementary Figure S1. GATA3 over-expression reduces primary tumor growth. (a) MB231 cells were transduced with a lentivirus system to express GATA3 (231-GATA3) or control vector (231-Empty). Western blot demonstrates expression of GATA3 in 231-GATA3 cells but not 231-Empty cells. -actin was used as a loading control. (b) 231-Empty and 231-GATA3 cells show equal basal levels of cell death in 2D culture conditions as measured by ELISA assay for cytoplasmic histone-associated-DNA-fragments. (c) Flow cytometric analysis showing cell cycle profiles of 231-Empty vs. 231-GATA3 cells. There is no difference in %S-phase in both cell lines. (d) Bright field images of 231-Empty and 231-GATA3 cells grown with complete media in 2D culture. Supplementary Figure S2. Orthotopic implantation of 231-Empty and 231-GATA3 cells. (a) 231-GATA3 exhibit reduced primary tumor growth compared to 231-Empty cells (tumor size measured every 2-4 days (vol.=l x w2 x 0.4). (b) increased survival of mice receiving 231GATA3 vs. 231-Empty cells. Mice were euthanized when the tumor reached 2 cm in diameter. (c) Primary 231-GATA3 mammary tumor xenografts display a more prominent epithelioid phenotype compared to a predominant spindyloid appearance of 231-Empty mammary xenografts. H&E staining. (d) Immunohistochemical staining of primary tumors confirmed positive staining for GATA3 in only 231-GATA3 tumors. There was an association of Ecadherin and CK8 staining with GATA3 expression in 231-GATA3 tumors. 231-Empty tumors were negative for GATA3 and had increased LOX staining compared to 231-GATA3 tumors. 7 Supplementary Figure S3. Reduced lung metastases in mice receiving 231-GATA3 cells by tail vein injection compared to 231-Empty cells. (a) Boyden Chamber assay of 231-Empty and 231-GATA3 cells showed a non-statistically significant trend for reduced number of cells invading through a Matrigel coated chamber. (b) Relative tumor cell infiltration in lungs as measured by immunofluorescence of lungs from tail vein injected mice with 231-Empty or 231GATA3 cells expressing GFP. Lungs were collected at the indicated times. (c) Immunofluorescence picture of the lung of 231-Empty and 231-GATA3 tail vein injected mice Supplementary Figure S4. H&E staining of metastatic lung lesions. Staining of (a) 231-Empty and 231-GATA3 lung lesions and of (b) 231-GATA3-Empty and 232-GATA3-LOX lung lesions from tail vein injected mice. Supplementary Figure S5. GATA3 promotes global transcriptome changes in MB231 cells. Bar Chart showing cellular and molecular functions altered upon GATA3 over-expression in MB231 cells. Supplementary Figure S6. GATA3 staining of metastatic lung lesions. Immunohistochemical staining for GATA3 of (a) 231-Empty and 231-GATA3 lung lesions and of (b) 231-GATA3Empty and 232-GATA3-LOX lung lesions from tail vein injected mice. Supplementary Figure S7. LOX staining of metastatic lung lesions. Immunohistochemical staining for LOX of (a) 231-Empty and 231-GATA3 lung lesions and of (b) 231-GATA3-Empty and 232-GATA3-LOX lung lesions from tail vein injected mice. 8 Supplementary Figure S8. Microarray analysis of 231-Empty vs. 231-GATA3 cells. (a) Hierarchical clustering analysis of 51 breast cancer cell lines (Neve et al., 2006) and the 231Empty and 231-GATA3 cells. Two hundred forty nine unique genes from the subtype predictor signature (Neve et al., 2006) are included in the analysis (multiple microarray probes per gene were reduced to the probe with the highest median intensity across samples). The heatmap gene expression values are relative differences within the respective datasets (gene-wise Z-score transformation in the 51 cell line collection and the 231-Empty and 231-GATA3 cells). The twoway hierarchical clustering of the combined data sets employs a distance metric of one minus Pearson’s correlation coefficient and average linkage algorithm. (b) Q-RT-PCR validation of changes in gene expression in the 231-Empty vs. 231-GATA3 cells. (c) Western blot showing CK18 and E-cadherin in 231-Emtpy vs. 231-GATA3. b-actin was used as a loading control. There is re-expression of E-cadherin and increased CK18 in 231-GATA3 vs. 231-Empty cells. Supplementary Figure S9. LOX reexpression does not reverse the phenotypic changes promoted by GATA3. (a) Bright field image of 231-GATA3-Empty and 231-GATA3-LOX cells. (b) Western blot of 231-GATA3 and 231-GATA3-LOX cells showed continued expression of E-cadherin upon reexpression of LOX. Dataset S1. The expression of 1273 probe sets were altered between 231-GATA3 and 231Empty cells (776 up- and 497 down-regulated in 231-GATA3 cells with fold change > 1.5 and p < 0.001, and false discovery rate (FDR) of 3%. See Excel file Chu_Dataset S1. 9 Dataset S2. List of genes increased in 231-GATA3 cells associated with the luminal phenotype compared to 231-Empty cells and list of genes reduced in 231-GATA3 associated with the basal compared to 231-Empty cells. References Barkan D, El Touny LH, Michalowski AM, Smith JA, Chu I, Davis AS, et al. (2010). Metastatic growth from dormant cells induced by a col-I-enriched fibrotic environment. Cancer Res. 70:5706-5716. Barkan D, Kleinman H, Simmons JL, Asmussen H, Kamaraju AK, Hoenorhoff MJ, et al. (2008). Inhibition of metastatic outgrowth from single dormant tumor cells by targeting the cytoskeleton. Cancer Res. 68:6241-6250. Fogelgren B, Polgar N, Szauter KM, Ujfaludi Z, Laczko R, Fong KS, et al. (2005). Cellular fibronectin binds to lysyl oxidase with high affinity and is critical for its proteolytic activation. J.Biol.Chem. 280:24690-24697. Hoenerhoff MJ, Chu I, Barkan D, Liu ZY, Datta S, Dimri GP, et al. (2009). BMI1 cooperates with H-RAS to induce an aggressive breast cancer phenotype with brain metastases. Oncogene 28:3022-3032. Kaneda A, Wakazono K, Tsukamoto T, Watanabe N, Yagi Y, Tatematsu M, et al. (2004). Lysyl oxidase is a tumor suppressor gene inactivated by methylation and loss of heterozygosity in human gastric cancers. Cancer Res. 64:6410-6415. Luger D, Silver PB, Tang J, Cua D, Chen Z, Iwakura Y, et al. (2008). Either a Th17 or a Th1 effector response can drive autoimmunity: conditions of disease induction affect dominant effector category. J.Exp.Med. 205:799-810. Neve RM, Chin K, Fridlyand J, Yeh J, Baehner FL, Fevr T, et al. (2006). A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 10:515-527. Palamakumbura AH, Trackman PC (2002). A fluorometric assay for detection of lysyl oxidase enzyme activity in biological samples. Anal.Biochem. 300:245-251. van de Vijver MJ, He YD, van't Veer LJ, Dai H, Hart AA, Voskuil DW, et al. (2002). A geneexpression signature as a predictor of survival in breast cancer. N.Engl.J.Med. 347:1999-2009. 10 Wu Z, Irizarry R, Gentleman R, Martinez-Murillo F, Spencer F (2004). A Model Basedd Background Adjustment for Oligonucleotide Expression Arrays. J Am Stat Assoc. 99:909-918. 11