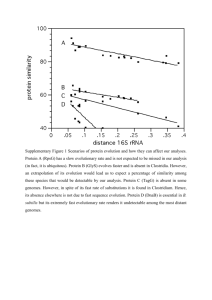
MorphologicalCharactersKey
advertisement

Supplementary Information for "Arthropod phylogeny based on eight molecular loci and morphology" Nature, V413, 157 MORPHOLOGICAL CHARACTERS Edgecombe et al. (2000) described 211 characters for Panarthropoda, most of which are used here. Characters in that work that pertained to Annelida or which are invariant in the ingroup for this study (synapomorphies of Panarthropoda) are excluded in the present analysis. For embryological characters 1-10, Halobiotus (Eibye-Jacobsen, 1997) is used for Eutardigrada; Glomeris (Dohle, 1964) is coded as a proxy for Sphaereotheriidae; Polyxenus is coded after Seifert (1960); Pauropodinae is based on Pauropus (Tiegs, 1947); Hanseniella is coded from Tiegs (1940); data for Chilopoda are coded for Scolopendra (Heymons, 1901), Scutigera (Dohle, 1970) and Lithobius (Hertzel, 1984); Campodea is based on Uzel (1898) and Tiegs (1942) with observations on the confamilial Lepidocampa by Ikeda and Machida (1998); japygids are scored based on Japyx (Silvestri, 1933), as interpreted by Ikeda and Machida (1998); Petrobius (Larink, 1969) and Pedetontus (Machida et al., 1990, 1994) are used for Machilidae; Lepisma (Heymons, 1897; Larink, 1983), Ctenolepisma and Therombia (Woodland, 1957) are used for Lepismatidae (summarised by Anderson, 1973); Baetis (Böhle, 1969) is coded as a proxy for Callibaetis (Baetidae); Blatta (Wheeler, 1889, summarised by Anderson, 1973) is coded for Blattodea; Locusta migratoria is coded from Roonwal (1936). Codings for Crustacea are based primarily on syntheses by Anderson (1973, 1982) and Weygoldt (1994). Opilio is the only taxon coded for Opiliones, using data from several other genera of Phalangioidea (e.g., Moritz, 1957; Juberthie, 1961, 1964). 1. Non-migratory gastrulation: 0, absent; 1, present. Anderson (1973) described a unique pattern of gastrulation in peripatopsid Onychophora, which has been regarded as an autapomorphy for Onychophora (Monge-Nájera, 1995). 2. Early cleavage: 0, total cleavage with radially oriented position of cleavage products; 1, intralecithal cleavage. A wide range of euarthropods share early total cleavage without oblique spindles, which Scholtz (1998) suggested is an autapomorphy of Euarthropoda. Tardigrades are described as having total cleavage, but the orientation is uncertain (Eibye-Jacobsen, 1997). Onychophora is coded from Anderson (1973, 1979), and Pycnogonida coded from Dogiel (1913). Edgecombe et al. (2000) coded Insecta as having complete superficial cleavage, but this is restricted to Dicondylia, Archaeognatha (Machilidae) having early total cleavage (Larink, 1997). The development of several species of Pycnogonida belonging to the families Ammotheidae, Callipallenidae, Endeidae and Phoxihilidiidae (Morgan, 1891; Meisenheimer, 1902; Sanchez, 1959) shows variation in the type of cleavage depending on the egg size. Species with small eggs have total cleavage with radially oriented position of cleavage cells (e.g. Endeis spinosa [Sanchez, 1959]), however different species of the family Ammotheidae are variable (Morgan, 1891; Meisenheimer, 1902; Sanchez, 1959). 3. 4. Blastokinesis with amnioserosal fold: 0, absent; 1, amniotic cavity open; 2, amniotic cavity closed (amnioserosal fold fuses beneath the embryo). Insect embryology is uniquely characterised by the division of the dorsal extra-embryonic ectoderm into an amnion and a serosa (Anderson, 1973; Machida and Ando, 1998). We follow Whiting et al. (1997) in regarding the closed amniotic cavity of Dicondylia as a modification of the open (Larink, 1983) amniotic cavity of Archaeognatha (i.e., the character is ordered). Aspects of the archaeognathan amnion, notably the yolk folds that encompass the embryo (Machida et al., 1994), may present unique characters for that group. Blastoderm cuticle (cuticular egg envelope): 0, absent; 1, present. Anderson (1973) identified a thin, highly resistant blastoderm cuticle beneath the chorion as shared by Progoneata, and lacking in Chilopoda, the latter largely based on Heymons’ (1901) work on Scolopendra. Machida and Ando (1998), however, cited Knoll’s (1974) study of Scutigera as indicating its presence within Chilopoda. Blastoderm cuticle is also present in Collembola and Diplura and, as in myriapods, is formed by the embryo and presumptive serosa (Machida and Ando, 1998). In Archaeognatha and Zygentoma, the cuticular egg envelope is formed by the serosa alone (without a contribution by the embryo) (Machida and Ando, 1998), whereas it is lacking in some pterygotes (e.g., Drosophila: Machida and Ando, 1998). Distinction from so-called blastoderm cuticle in Xiphosura (Anderson, 1973:370) is required. 5. 6. 7. 8. 9. 10. 11. 12. 13. Dorsal closure of embryo: 0, definitive dorsal closure (dorsal covering of embryo participates in the definitive dorsal closure); 1, provisional dorsal closure (embryonic dorsal covering degenerates without participating in the definitve closure, which is exclusively derived from the embryo). Definitive and provisional dorsal closure of the embryo are as defined by Machida and Ando (1998) to describe the cover of the embryo and hatching in myriapods (Pauropoda, Symphyla, Chilopoda) and hexapods (Collembola, Diplura, Insecta), respectively. Provisional closure in insects was summarised by Johannsen and Butt (1941:56-57). Ectoteloblasts forming part of metanaupliar/egg-naupliar region of germ band: 0, absent; 1, present, at anterior border of blastopore. Ectoteloblasts are specialised stem cells that give rise to the ectoderm of most postnaupliar segments in Malacostraca (Gerberding, 1997). They are absent in branchiopods [Daphnia based on the cladoceran Leptodora kindti (Gerberding, 1997)], and are lacking only in Amphipoda among the Malacostraca (Dohle and Scholtz, 1988; Weygoldt, 1994). Ectoteloblasts in leptostracans, stomatopods, syncarids and decapods are characterised by their circular or semi-circular arrangement at the anterior border of the blastopore, whilst those of peracarids are arranged in a single row (Weygoldt, 1994). Cells that form the early ectodermal material of the germ band in Cirripedia have been called ectotelobasts (Anderson, 1969, 1973), but Dohle (1972) considered morphological differences from the ectoteloblasts of Malacostraca to rule out homology (Gerberding, 1997). Scholtz (2000) summarised evidence for precise homologies in malacostracan teloblasts, with most groups sharing 19 ectoteloblasts and 8 mesoteloblasts. The latter share detailed similarities in their arrangement (one median pair and three lateral pairs). Caudal papilla: 0, absent; 1, present. Scholtz (2000) identified an anteroventrally folded caudal papilla as unique to Malacostraca. The caudal papilla is a ventrally-pointing tube derived from the preanal growth zone, containing the proctodaeum. It is folded anteroventrally in egg-nauplii. A caudal papilla is lacking in some peracarid groups (Tanaidacea, Isopoda, Cumacea, Mictacea). Fat Body: 0, absent; 1, fat body cells develop from vitellophages in yolk; 2, fat body cells develop from walls of mesodermal somites. The presence of a cephalic storage organ, the fat body, has been identified as an atelocerate synapomorphy (Boudreaux, 1979). Anderson (1973) made a distinction between vitellophagal fat body cells (Symphyla + Pauropoda + Diplopoda) and an origin of the fat body in the mesoderm (Chilopoda + Hexapoda). Dohle (1980) upheld this distinction, and employed the former condition as evidence for monophyly of the progoneate myriapods. A partial uncertainty coding (states 1 or 2, but not 0) is employed for several taxa in which a fat body is present but its embryological origin is unknown. Fate map ordering of embryonic tissues: 0, presumptive mesoderm posterior to presumptive midgut; 1, presumptive mesoderm anterior to midgut; 2, mesoderm midventral, cells sink and proliferate, midgut internalises during cleavage; 3, mesoderm diffuse through the ectoderm; 4, midgut develops from anterior and posterior rudiments at each end of midventral mesoderm band. Fate map patterns follow Anderson (1973, 1979) and Schram (1978). The phylogenetic value of fate maps has been criticised (Weygoldt, 1979), but Anderson (1982) reiterated the similarity of patterns across some major groups (e.g., all Crustacea can be uniquely characterised by state 1 above fide Anderson, 1982). Embryological development: 0, with a growth zone giving rise to both the prosoma and opisthosoma; 1, with a growth zone giving rise to the opisthosoma (Anderson, 1973; Dunlop and Webster, 1999). A growth zone giving rise to the ambulatory segments of the prosoma and the opisthosoma is found in scorpions (Euscorpius: Yoshikura, 1975) and xiphosurans, while in other arachnids this growth zone gives rise to the opisthosoma only, the prosoma developing directly from the blastoderm. Based on the latter observation, we code Mastigoproctus as described for Typopeltis (Anderson, 1973). Engrailed expressed in mesoderm patterning: 0, present; 1, absent. Zrzavý and Štys (1995) surveyed "compartment-like patterning" in the mesoderm of annelids and arthropods, as marked by engrailed expression. Limited data are available to indicate the absence of such mesodermal patterning in some insects and crustaceans (Artemia, peracarids and decapods: see Dohle, 1997) versus its presence in at least some chilopods, onychophorans and annelids. Dohle (1997) characterised the difference between myriapods (Ethmostigmus: Whitington et al., 1991) and hexapods/crustaceans as the former having engrailed staining in mesoderm and in the cytoplasm whereas the latter have en-product confined to the nuclei of ectodermal cells. Epimorphic development: 0, absent; 1, present. Several arthropod groups have been diagnosed by epimorphosis, hatching with the complete complement of segments (e.g., Epimorpha within Chilopoda; Diplura + Insecta fide Kraus, 1998). Nauplius larva or egg-nauplius: 0, absent; 1, present. Edgecombe et al. (2000) coded the nauplius (a swimming larva) as inapplicable for terrestrial taxa. Within Entomostraca, the nauplius is lacking only in Cladocera (Gerberding, 1997). Most malacostracan exemplars considered here have direct development (e.g., Leptostraca and Peracarida), but can be scored for the presence of an eggnauplius. Some peracarids (Amphipoda, Tanaidacea, Cumacea, Isopoda), however, lack an egg- 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. nauplius (Scholtz, 2000). Dahms (2000) summarised specific details of the naupliar apparatus that imply a single origin for this larval type, but Scholtz (2000) argued that a free-living nauplius evolved from an egg-nauplius within Malacostraca. Bivalved secondary shield grows from tergite of maxillary segment during larval development (heliophora stage): 0, absent; 1, present. Walossek (1995) and Walossek and Müller (1998a) used a unique secondary shield during larval development as a synapomorphy for Onychura (= Conchostraca + Cladocera). Pupoid stage (motionless stage after hatching, pupoid remains encased in embryonic cuticle): 0, absent; 1, present. Anderson (1973) summarised evidence for a pupoid stage in Chilopoda, Diplopoda and Pauropoda. Dohle (1998), however, identified a pupoid stage as confined to diplopods and pauropods, and Enghoff et al. (1993) recognised the pupoid as a so-called hexapod stage. We recognise the peripatoid and foetoid stages of Epimorpha (Chilopoda) as character 255. Glomeris is coded as a proxy for Sphaerotheriidae. Sclerotisation of cuticle into hard, articulated tergal exoskeleton: 0, absent; 1, present. Cuticle calcification: 0, absent; 1, present. Enghoff (1984) regarded a calcified cuticle as a synapomorphy of Chilognatha. Calcium carbonate impregnation of the cuticle is also coded for Reptantia but not for the shell of Cirripedia (coding is based on the cuticle). Cilia in photoreceptors: 0, present; 1, absent. Wheeler et al. (1993) coded for a reduction in cilia in onychophorans and euarthropods relative to annelids. We modify the coding of Edgecombe et al. (2000: character 17), which coded the euarthropod state with reference to cilia being present only in sperm (versus also present in the photoreceptors in Onychophora). A phylum-level groundplan has been adopted. Tendon cells with tonofilaments penetrating epidermis: 0, absent; 1, present. Boudreaux (1979) and Wägele (1993) acknowledged tonofilaments as a euarthropod synapomorphy, and Dewel and Dewel (1997) confirmed their absence in onychophorans and tardigrades. A phylum-level groundplan has been adopted. Dorsal longitudinal ecdysial suture with forking on head: 0, absent; 1, present. Edgecombe et al. (2000: character 20) coded variation in ecdysial suture patterns in Panarthropoda as a multistate character. Their coding for myriapods and hexapods followed Boudreaux (1979), who regarded ecdysis involving transverse rupture between the head and trunk to be diagnostic of Myriapoda. While a head-trunk split is reasonably regarded as the groundplan state for Myriapoda, details of ecdysial patterns in various taxa complicate coding. For example, within Chilopoda, Lithobiomorpha moult at the transverse suture on the head shield (Lewis, 1981), and it is probable that Craterostigmus has the same ecdysial pattern, having identical sutures on the head shield (see character 21). In Geophilomorpha, Lewis (1961) described ecdysial rupture across the anterior part of the head shield in Strigamia. These deviations lead us to abandon coding head-trunk rupture across Myriapoda. Likewise, within Crustacea group-specific ecdysial patterns can be identified (e.g., biphasic moult with the break between pereionites 4 and 5 in Isopoda: Brusca and Wilson, 1991), but coding states that can be homologised across Panarthropoda is complicated. We have thus restricted coding of ecdysial patterns. Boudreaux (1979) claimed dorsal longitudinal ecdysis to be diagnostic for Hexapoda, but Snodgrass (1952:269) specified that the latter pertained to Insecta in particular, whereas Collembola and Protura have a head-trunk ecdysial split (Kaufman, 1967:16). Though Onychophora also have a dorsal longitudinal rupture of the cuticle, the insect pattern possesses details, such as dorsal forking (frontal line) on the head that permit distinction. In japygids the Y-shaped frontal line is lacking, but the posterior cornonal suture is present. Transverse and antenocellar sutures on head shield: 0, absent; 1, present. Edgecombe et al. (1999: character 7) coded an ecdysial complex of transverse and antenocellar sutures (sensu Crabill, 1960) as shared by Craterostigmus and Lithobiomorpha. The frontal line in some Geophilomorpha may be homologous with the transverse suture, but the antennocellar suture is lacking. Resilin protein: 0, absent; 1, present. Weygoldt (1986) indicated that the spiral protein resilin is known only from euarthropods (summarised by Hackman, 1984) and onychophorans. Nielsen (1995), however, mapped resilin onto the tree as a euarthropod synapomorphy, indicating its absence in tardigrades and onychophorans. We follow the phylum-level groundplan assigned by Nielsen (1995). Moulting gland: 0, absent; 1, present. Wägele (1993) cited a moulting gland as a diagnostic character of Mandibulata. This was based on a proposed homology between the Y-organ of Malacostraca (Fingerman, 1987) and the prothoracic gland of insects. Wägele noted that such moulting glands in insects and crustaceans are hypodermal derivations of the second maxilla, and are absent in chelicerates. An alleged ecdysial gland in some chilopods (Lithobiomorpha: Seifert and Rosenberg, 1974; glandula capitis in Scutigeromorpha: Seifert, 1979) may be homologous. Evidence for an ecdysial gland has not been found in other myriapods (Tombes, 1979) except for polyxenid millipedes (glandula perioesophagealis; Seifert, 1979). Studies of branchiopods have not discovered similar moulting glands although moulting hormones appear to be present (Martin, 1992). 24. Bismuth staining of Golgi complex beads: 0, not staining; 1, staining. Locke and Huie (1977) observed Golgi beads to stain with bismuth (also judged by the reaction with interchromatin and perichromatin granules present in most nuclei) in various euarthropods and tardigrades, but not in Onychophora (Epiperipatus), Annelida, Mollusca, Nematoda, or Platyhelminthes. Bismouth staining was observed for Limulus, an undetermined isopod, Orconectes (coding for Reptantia) and Locusta for the terminals here coded. However this pattern seems to be a putative synapomorphy for Arthropoda, since it has also been recorded in other chelicerates, millipedes and several neopteran insects. The three exemplars of Pycnogonida are here coded based on the undetermined pycnogonid of Locke and Huie (1977). 25. Metanephridia with sacculus with podocytes: 0, absent; 1, present. While metanephridia are probably plesiomorphic for arthropods (Fauchald and Rouse, 1997), the sacculus and podocytes are novel nephridial structures for onychophorans and euarthropods, lacking in tardigrades (Nielsen, 1998; Schmidt-Rhaesa et al., 1998) and pycnogonids (King, 1973). A groundplan coding has been adopted. 26. Distribution of segmental glands: 0, on many segments; 1, in at most last four cephalic segments and first two post-cephalic segments; 2, on second antennal and maxillary segments only. Definition of the basic euarthropod distribution of segmental glands, a reduction from that in Onychophora, follows Weygoldt (1986). We have not attempted to define all variants of segmental gland distribution within Euarthropoda, and state 1 above is an artificial grouping. A more advanced reduction in Crustacea, restricted to the antennal and maxillary segments, has been regarded as a crustacean synapomorphy (Lauterbach, 1983, 1986). Walossek and Müller (1990:410) considered remipedes (Schram and Lewis, 1989) and anostracans to deviate from this state in possessing additional cephalic segmental glands, but Wägele (1993) dismissed these as integumental glands and embryonic mesodermal cells, respectively. 27. Tömösváry organ ("temporal organs" at side of head behind insertion of antenna): 0, absent; 1, present. Homology of Tömösváry organs across the Myriapoda has been widely accepted (Snodgrass, 1952), but relationships to similarly positioned structures in hexapods are contentious. François (1969), for example, homologised the pseudocellus of Protura with Tömösváry organs, whereas Tuxen (1959) regarded them as antennal vestiges on the basis of musculation. The postantennal organs of Collembola may also be homologous (Haupt, 1979), and as such have been coded here. We have scored the temporal organs of Ellipura as homologous with those of Myriapoda, following Haupt (1979) and Bitsch and Bitsch (1998, 2000), though Dohle (1997) questioned the likelihood of this homology. The homologue of the Tömösváry organ in Craterostigmus is a ringed organ on the cephalic pleurite (Shear and Bonamo, 1988, fig. 50; Dohle, 1990, fig. 2), in a similar position as in Lithobiomorpha. Despite arguments that the organ could be non-functional in Craterostigmus (Borucki, 1996), it is unambiguously present. Štys and Zrzavý (1994) refer to the possibility of a Tömösváry organ in Diplura (Japygina), but this homology has not been substantiated. 28. Salivary gland reservoir: 0, absent; 1, present. Monge-Nájera (1995) identified a salivary gland reservoir as an onychophoran autapomorphy. 29. Malpighian tubules formed as endodermal extensions of the midgut: 0, absent, 1, present. Shultz (1990) claimed that endodermal Malpighian tubules are unique to Arachnida and, despite their absence in some ingroup taxa (such as Opiliones), resolved them as an arachnid autapomorphy. Hexapod and myriapod Malpighian tubules, in contrast, are extensions of the ectodermal hindgut (see character 30). The non-homology of these structures is generally recognised, and we have accordingly coded them as separate characters. 30. Malpighian tubules formed as ectodermal extensions of the hindgut: 0, absent; 1, single pair of Malpighian tubules at juncture of midgut and hindgut; 2, multiple pairs of tubules at anterior end of hindgut. The presence of Malpighian tubules in Collembola is dubious (Clarke, 1979; Bitsch and Bitsch, 1998; here coded as absent), while Protura have several pairs of papillae behind the midguthindgut junction (see character 31). Distinct conditions can be recognised within the myriapods and the ectognath hexapods, and serve as the basis for states 1 and 2 above. The homology of insect Malpighian tubules (whether ectodermal or entodermal) is controversial, but an ectodermal origin is best supported (Dohle, 1997). One or a small pair of supernumerary Malpighian tubules is present in some chilopods (Prunescu and Prunescu, 1996). The so-called Malpighian tubules of eutardigrades are not in contact with cuticle and as such do not appear to be ectodermal in origin (Møbjerg and Dahl, 1996). 31. Form of ectodermal Malpighian tubules: 0, elongate; 1, papillate. The Malpighian tubules are elongate in myriapods and most hexapods, Bitsch and Bitsch (1998: character 14) interpreting this as the basal state for Atelocerata. Bitsch and Bitsch coded the very similar papillae of Protura and Campodeina as homologous, a procedure adopted here. 32. Neck organ: 0, absent; 1, present. Martin and Laverack (1992) reviewed the so-called dorsal organ of crustaceans, and codings for Crustacea follow their citation of its presence. They regarded this organ as homologous in Malacostraca and Branchiopoda, whereas Walossek (1993) specified more precise details shared by Branchiopoda and Maxillopoda, wherein a ‘neck organ’ may have evolved around a more primitive body, the dorsal organ. The term ‘neck organ’ avoids confusion with the region of extra-embryonic ectoderm that is commonly called a ‘dorsal organ’ in many groups of arthropods (Fioroni, 1980). 33. Hemocyanin: 0, absent; 1, present. Codings for the presence of hemocyanin follow Clarke (1979). In pycnogonids, hemocyanin is only found dissolved in the plasma (Arnaud and Bamber, 1987), without cyanocytes sensu Gupta (1979) (but see the comment on Ammothea by Aranud and Bamber, 1987:20). Here all the pycnogonids represented have been coded for the presence of hemocyanin. Some remipedes have large hemocyanin crystals scattered throughout the head and swimming appendage tissue (J. Yager, pers. comm., 1998). Elsewhere in Crustaeca, hemocyanin occurs in stomatopods and decapods (Goodwin, 1960). Within Chilopoda, Scutigeromorpha have hemocyanin as the oxygen transport molecule (Mangum et al., 1985), versus gaseous exchange between the tracheae and tissues in Pleurostigmophora (Hilken, 1997). Hemocyanin is lacking in hexapods (Beintema et al., 1994) except for a single report in Schistocerca (Burmester, 2001), but is present in some diplopods (Spirostreptus: Jaenickel et al., 1999; coding is applied to Juliformia based on these data). Hemocyanin has been sequenced in Limulus (Mangum et al. 1985) and Tachypleus (Linzen et al., 1985), and is known for Mastigoproctus (Redmond, 1968), scorpions and some spiders (Clarke, 1979; Strazny and Perry, 1987), codings adopted here for Buthidae and Mygalomorphae. The gene sequence data of hemocyanin are known for several chelicerates and decapod crustaceans (Burmester, 2001). 34. Hemoglobin: 0, absent; 1, present. Hemoglobin has a restricted distribution within Arthropoda. It occurs in many entomostracan crustaceans (see Goodwin, 1960, Table 1) but not in Malacostraca. Clarke (1979) indicated that it is unreported in Chelicerata. Within the Insecta, the occurrence of hemocyanin is exceptional (e.g., present in some chironomid larvae and various parasitic Diptera) and is not general for any hexapod order. Hemoglobin has been reported in barnacles (Southward, 1963), but Waite and Walker (1988) were unable to detect it (or any respiratory pigments) in several balanomorphs. 35. Subcutaneous hemal channels in body wall: 0, absent; 1, present. Presence is unique to Onychophora (Brusca and Brusca, 1990; Monge-Nájera, 1995). A phylum-level groundplan has been adopted. 36. Dorsal heart with segmental ostia and pericardial sinus: 0, absent; 1, present. The dorsal, ostiate heart and pericardial sinus/septum is shared by Onychophora and Euarthropoda, but it is absent in tardigrades and pauropods. We have not coded with the assumption that these absences are due to miniaturisation. A heart is lacking in Thoracica, the rostral sinus serving this function (Schram and Hof, 1998), whereas Calanoidea retain a well-developed heart with at least one ostium (Boxshall, 1982). 37. Circumesophageal circulatory vessel ring with ventral, trumpet-shaped opening towards head: 0, absent; 1, present. Gereben-Krenn and Pass (1999) documented detailed similarity in the circumesophageal circulatory vessel ring of Diplura (Campodea and Catajapyx), and made homologies with vessel rings in machilids, lepismatids and collembolans. Homology with the similarly positioned mandibular arch of some Chilopoda (documented by Fahlander, 1938 and Rilling, 1968 for Lithobius) was suggested by Gereben-Krenn and Pass. This homology is strengthened by the presence of a similar anterior-opening, trumpet-shaped vessel in Lithobius. These structures are, however, absent in Scutigeromorpha (Fahlander, 1938) as well as Pterygota and Protura (Bitsch and Bitsch, 1998). We are unaware of potentially homologous vessels in Crustacea. In Malacostraca (e.g., Leptostraca, Anaspidacea, Lophogastrida), the dorsal vessel has an anterior extension that supplies the rostrum, both pairs of antennae and the labrum but not the mandibles. This vessel system is dorsal and anterior to the esophagus and the mouth, and lacks a circumesophageal ring (S. Richter, pers. comm., 2000) (see Schmitz, 1992, fig.70 for Amphipoda). 38. Slit sensilla: 0, absent; 1, present. Slit sensilla are small clefts or slits in the cuticle, used in detecting compressional forces acting on the exoskeleton (Shultz, 1990). They have been recognised as a synapomorphy for Arachnida, but are (doubtfully) present in the extinct Eurypterida (Dunlop and Braddy, 1997). 39. Ganglion formation: 0, ganglia formed from an invagination of the ventral organ; 1, neuroblasts. Neuroblasts are the precursor cells of the embryonic ganglia in insects and crustaceans that divide according to the stem cell mode, and are positioned on the ventral germ band adjacent to the midline (Dohle, 2001). In Myriapoda, neuroblasts are variably described as lacking (Osorio and Bacon, 1994; Zrzavý and Štys, 1994) or present (Scutigera: Knoll, 1974). Since recent treatments (e.g., Gerberding, 1997) consider myriapod ganglia to develop without neuroblasts, we are reluctant to 40. 41. 42. 43. 44. 45. accept Knoll’s (1974) interpretation of Scutigera. Scutigera appears to have some larger cells in the neurogenic area but the asymmetric division that characterises neuroblasts is not shown (G. Scholtz, pers. comm., 1999). Dohle (1997, 2001) indicated that onychophoran and chelicerate ganglion formation resembles that of centipedes and millipedes. Weygoldt (1985, 1998) likewise observed similarity between the nervous system in chelicerates, myriapods and onychophorans (Paraperipatus; Pflugfelder, 1948), in each case arising by invagination of the ventral organ. Gerberding (1997) showed that Cladocera (Daphnia coded here based on Leptodora) have cells with the characteristics of neuroblasts, and that their morphology is identical to that in Malacostraca (Reptantia coded using Cherax; Scholtz, 1992; Isopoda coded based on Dohle and Scholtz, 1988). The absence of cells with the characteristics of neuroblasts in pauropods [Pauropus], symphylans [Hanseniella] and diplopods [Glomeris] in older studies was noted by Whitington and Bacon (1998), and absence in these groups is coded based on these data. Insect and crustacean neuroblasts differ in that the former delaminate from the surface and form a separate layer whereas the latter lie superficially (Gerberding, 1997), and do not differentiate in waves as is the case in insects (Dohle, 2001). Edgecombe et al. (2000: character 39) coded the development of neuroblasts in Araneae and Scorpiones based on reports by Yoshikura (1955) and Mathew (1956). These data require confirmation because all other indications suggest ganglion development in chelicerates occurs by invagination. Early differentiating neurons vMP2, aCC, pCC, RP2 and MP4: 0, absent; 1, present. The identity and relative positions of early differentiating neurons exhibit impressive similarities between insects and some malacostracan crustaceans (Whitington et al., 1993). This similarity (state 1 above) may be defined in terms of putative homologies of at least five neurons (Dohle, 2001). Data for Myriapoda are limited to Ethmostigmus (coded for Scolopendridae), for which Whitington et al. (1991) documented different patterns of segmental neurons,with the pioneer neurons of insects and eumalacostracans unidentifable. Neurons involved in early axon growth in Ethmostigmus are widely separated from each other, and axons originating from the brain initiate early central axon pathways instead of neurons in each segmental neuromere, as is the case for insects (Dohle, 2001). Globuli cells: 0, confined mainly to brain, in massive clusters; 1, making up majority of neuropil and ventral layer of ventral nerve cord. Schürmann (1995) recognised that onychophorans are specialised relative to annelids and other arthropods in that globuli cells are the main cell type in the brain and also form a massive ventral layer in the ventral nerve cord. Corpora allata: 0, absent; 1, present. Corpora allata are present in insects, proturans, collembolans and diplurans (Japygina but not Campodeina) (Cassagnau and Juberthie, 1983), and are regarded as a hexapod apomorphy (Wägele, 1993). A groundplan coding is adopted. Intrinsic secretory cells in protocerebral neurohemal organ: 0, absent; 1, present. Gupta (1983) reviewed the distribution of intrinsic secretory cells in neurohemal organs, assuming that the presence of these cells was a derived state. This character has been coded based on the state of the primary protocerebral neurohemal organ: the sinus gland in some crustaceans, the cephalic gland in symphylans, the cerebral gland in chilopods, Gabe’s organ in diplopods, the corpora cardiaca in hexapods, Schneider’s Organ I in spiders, the Paraganglionic plates in Opiliones (groundplan coded from Juberthie, 1983) and the stomatogastric ganglia of scorpions [see Tombes (1979) and references in Gupta (1983)]. In isopods, some intrinsic secretory cells are present in the lateral cephalic nerve plexus, but not in the sinus gland (Wägele, 1983). In some Diplopoda, intrinsic secretory cells are associated with the paraesophageal body or the periesophageal blood sinuses that are variably associated with the paraesophageal body (Juberthie-Jupeau, 1983), but coding is based solely on Gabe’s organ. Enlarged epipharyngeal ganglia: 0, absent; 1, present. Protura and Collembola share specialised masses of sensory and secretory cells in the epipharyngeal region (François, 1969; Kristensen, 1991). Ganglia of pre-oesophageal brain: 0, protocerebrum; 1, protocerebrum and deutocerebrum: 2, protocerebrum and tritocerebrum; 3, proto-, deuto- and tritocerebra. A tripartite brain has been proposed as a synapomorphy for Mandibulata (Brusca and Brusca, 1990), or a character uniting tardigrades and euarthropods (Nielsen, 1995). The most recent assessment of homologies of the tardigrade brain, however, suggests that its components (dorsal and ventral cones, internal cirrus and their respective ganglia) are homologous only to the protocerebrum of arthropods, and not to the deutocerebrum or tritocerebrum (Dewel and Dewel, 1996). Furthermore, the incorporation of the tritocerebrum into a pre-oesophageal brain is not shared by all crustaceans; Walossek and Müller (1998b) showed that the brain includes the proto- and deutocerebral segments only in Entomostraca, such as branchiopods. Coding for this character recognises a distinction between the "bipartite" brain of Entomostraca (and Leptostraca fide Calman, 1909) and that of Chelicerata (including Pycnogonida) in that the latter is classically considered to lack the deutocerebrum (state 2 above). The alternative homology scheme suggested by Hox gene expression data (Damen et al., 1998; Telford and Thomas, 1998) equates states 1 and 2. 46. Ganglia of post-oral appendages fused into single nerve mass: 0, absent; 1, present. Zrzavý et al. (1998) coded fusion of anterior ganglia as a synapomorphy for pycnogonids and euchelicerates. We do not regard the fusion of the palp and oviger nerves to the subesophageal ganglion in pycnogonids (Arnaud and Bamber, 1987, fig. 12) to be as reliable an apomorphy as the fusion of all post-oral, cephalic, limb-bearing segments to the subesophageal nerve mass in euchelicerates. The coding used here is thus at the same level of generality as Moura and Christofferson’s (1996) citation of fusion of post-cephalic ganglia into a subesophageal mass (euchelicerate apomorphy). An additional state might be recognised for arachnids, which fuse abdominal ganglia to the nerve mass (Wegerhoff and Breidbach, 1995). 47. Fan-shaped body with neurons extending laterally into protocerebral lobes: 0, absent; 1, present. Strausfeld (1998) optimised an unpaired neuropil called a fan-shaped body (Williams, 1975) as a synapomorphy of crustaceans and hexapods. Strausfeld described the equivalent of the fan-shaped body as present in all crustaceans so far examined, but since his sampling (1998, fig. 2) is limited to Eumalacostraca we have confined coding to crustacean groups explicitly treated by him (Isopoda and Reptantia) except to record the absence of a "central body" in the cephalocarid Hutchinsoniella (Elofsson and Hessler, 1990), and to code for its presence in Anaspides (Hanström, 1934; see Coineau, 1994, fig. 400). Except where indicated otherwise, coding for this and other brain characters (48-52) mirrors Strausfeld’s sample as follows: Euperipatoides, Lithobius, Limulus, Mastigoproctus, Thermobia, Periplaneta and Drosophila are coded as exemplars; Ligia is coded for Oniscidea; Neanurinae is coded for Collembola; Locusta is coded based on the confamilial Barytettix; Orthoporus is coded for Juliformia; Centruroides is coded as an exemplar of Buthidae; Opilio is coded based on the confamilial Rilaena (Wegeroff and Breidbach, 1995); Lecythorhynchus is coded as an exemplar of Ammotheidae. For several brain characters, Hutchinsoniella is coded from Elofsson and Hessler (1990). 48. Arcuate body in brain: 0, absent; 1, present. Pycnogonids and chelicerates (spiders, Limulus, scorpions, opilionids and amblypygids) share an unpaired crescent-shaped neuropil adjacent to the posterior margin of the supraesophageal mass. Strausfeld (1998) regarded this so-called arcuate body, which has highly conserved structure (e.g., palisades of columnar neurons intersected by discrete layers of tangential processes), to be a chelicerate synapomorphy. 49. Nucleation of neuropils: 0, neuropils comprise discrete homolateral nuclei with mirror symmetry on each side of midline; 1, neuropil region extends across midline. Strausfeld (1998) united Onychophora and Diplopoda based on brain characters. A putative synapomorphy of this group is the continuity of neuropils across the midline (versus distinctly nucleated neuropils in chilopods and hexapods). Chelicerates, however, also have midline confluence of the neuropils (Strausfeld et al., 1995: character 13). 50. Ellipsoid body in brain: 0, absent; 1, present. In insects, the ellipsoid body is a generally fusiform neuropil that crosses the midline beneath and slightly anterior to the fan-shaped body, from which it receives palisades of axons (Strausfeld, 1998). Strausfeld optimised the ellipsoid body as a pterygote synapomorphy. 51. Noduli in brain: 0, absent; 1, present. Noduli (Strausfeld, 1998) are a pair of ball-shaped neuropils connected to the fan-shaped body and ellipsoid body in Hexapoda. 52. Protocerebral bridge: 0, absent; 1, present. A bridge of neuropil (protocerebral bridge of Strausfeld, 1998) lies posteriorly in the brain, supports the fan-shaped body, and connects the left and right protocerebral hemispheres in ptyergote insects and the isopod Ligia. In both groups the bridge is composed of small bushy dendrites that intersect axons between the two halves of the brain, and these dendrites supply a complex pattern of axonal projections to the fan-shaped body. This bridge is lacking in decapods and apterygote hexapods fide Strausfeld (1998), and in Hutchinsoniella fide Elofsson and Hessler (1990). 53. Cephalon composed of one pair of preoral appendages and three pairs of postoral appendages: 0, absent; 1, present. The composition of the fundamental euarthropod head has considered information from fossils as well as extant taxa. Weygoldt’s (1986) proposal that the basic euarthropod cephalon included four pairs of post-oral appendages is in conflict with well-known paleontological data (e.g., three pairs of post-oral biramous limbs in trilobites). Walossek’s (1993) contention that the crown-group euarthropod cephalon is composed of preoral antennae and an additional three pairs of limbs is corroborated by engrailed expression in crustaceans and insects, in which an engrailed stripe on the first maxillary segment indicates the original posterior limit of the head (Scholtz, 1998). 54. Cephalic kinesis: 0, absent; 1, present. Cephalic kinesis, defined as movable ophthalmic and antennular segments and an articulated rostrum (Kunze, 1983), is shared by the Leptostraca and the Stomatopoda but not other Crustacea. Siewing (1963) emphasised shared complicated musculature in defence of homology between the movable "procephalon" of Leptostraca and Stomatopoda. 55. Flattened head capsule, with head bent posterior to the clypeus, accommodating antennae at 56. 57. 58. 59. 60. 61. 62. 63. 64. 65. anterior margin of head: 0, absent; 1, present. Dohle (1985) and Shear and Bonamo (1988) emphasised the peculiar construction of the flattened head capsule of Pleurostigmophora, with the clypeal region of the head becoming ventral and the mouth pushed backwards. Snodgrass (1952:212) considered the ventral configuration of the clypeus in Pleurostigmophora to result from accommodating the antennae at the anterior margin of the head. These details permit distinction from superficially similar flattened head shields in other arthropods (e.g., remipedes). Clypeofrontal sulcus (epistomal suture):0, absent; 1, present. Bitsch and Bitsch (2000: character 5) distinguished an undivided clypeofrons in Symphyla and most basal hexapods from a condition in which a clypeofrontal sulcus is present (Zygentoma, most Pterygota). Identity of the clypeofrons involves the attachment of the dorsal muscles of the preoral cavity and pharynx (Bitsch and Bitsch, 2000), rendering applicability outside Hexapoda and Myriapoda dubious. Codings follow Bitsch and Bitsch, with absence in Blattodea indicated by Snodgrass (1935). We regard the presence of a homologue of the insect clypeofrontal sulcus in chilopods as dubious (Bitsch and Bitsch coded for this sulcus in Lithobius). The juncture between the "frons" and clypeus in Pleurostigmophora is marked at the base of the sclerotised bridge between the antennae by reduced sclerotisation, usually without a suture or sulcus, and lacking internal expression as an epistomal ridge that is often present in insects. Lateral eyes: 0, absent; 1, simple lens with cup-shaped retina; 2, stemmata with rhabdom of multilayered retinular cells; 3, facetted; 4, onychophoran eye. Compound eyes (state 3) are present in fossil scorpions (Kjellsvig-Waering, 1986) and diplurans (Kukalová-Peck, 1991), whereas extant taxa possess states 1 and 0, respectively. Fossil diplopods have been considered to have compound eyes (Kraus, 1974), though Spies (1981) interpreted them as pseudofacetted; extant Chilognatha have stemmata (state 2). Rather than coding the pseudofacetted eye of Scutigeromorpha as an uninformative autapomorphy, we follow Paulus (1979, 1986, 1989, 2000) in interpreting this eye as a modification of myriapod stemmata (state 2) based on ultrastructural similarities. Myriapod lateral eyes possess a rhabdom composed of two (Scutigeromorpha and Polyxenida) or many (Pleurostigmophora and Chilognatha) layers of retinular cells. Paulus (1986) considered the layering of the rhabdom as a probable synapomorphy for Myriapoda, noting a similar construction only in the larval eyes of certain insects. Compound eyes medial margins: 0, separate; 1, medially contiguous. The medial coalescence of the compound eye has been treated as a shared derived character of the Archaeognatha (Hennig, 1981; Kristensen, 1991). The approximation of the antennal bases in archaeognathans is regarded as a correlated character (cf. Kraus, 1998), an effect of a medial repositioning of cephalic structures. Compound eye stalked, basally articulated: 0, absent (eye sessile); 1, present. Coding is restricted to arthropods with lateral facetted eyes. Compound eyes internalised early in ontogeny, shifted dorsally into a cuticular pocket: 0, absent; 1, present. Walossek (1995) defended monophyly of Phyllopoda (including Notostraca, Conchostraca and Cladocera) based on the internalisation of the eyes. Ophthalmic ridges: 0, absent; 1, present. The lateral eyes of Xiphosura are uniquely set against ophthalmic ridges. Some representatives of the extinct taxon Chasmataspida, which is probably most closely related to Eurypterida, may also have ophthalmic ridges (Anderson et al., 2000). "Tetraconate" eye (two corneageneous cells, four Semper cells in crystalline cone, retinula with eight cells): 0, absent (variable, higher number of parts); 1, present. Despite some variation in precise numbers of subunits within crustacean and hexapod eyes, Paulus (1979, 1989) postulated that a groundpattern (state 1 above) could be interpreted for the common ancestor of these clades. Melzer et al. (1997) observed that processes from the cone cells pass between the retinula cells to the proximal basement membrane in the same, highly detailed manner in Crustacea (Triops, Lepidurus, Panulirus, Squilla, Argulus) and Insecta (Machilis, Drosophila) (see also Dohle, 2001). Attempts to interpret myriapod stemmata as a modified mandibulate ommatidium (Paulus, 1986, 1989) have not been compelling. Paulus (2000) suggested that remnants of Semper cells could be present in Polyxenus, but admitted that the evidence is not convincing. Two primary pigment cells in ommatidium: 0, two corneagenous cells lacking pigment grains; 1, two corneagenous cells contain pigment grains. Paulus (1979) homologised a pair of corneagenous cells in Crustacea with the two primary pigment cells of hexapods. Scoring for this character is restricted to those taxa with two corneagenous cells, part of the tetraconate eye (character 62). Crystalline cone cells: 0, tetrapartite crystalline cone; 1, cone bipartitite, with two accessory cells. Richter (1999) united Anaspidacea, Euphausiacea and Peracarida based on a modification of a tetrapartite crystalline cone (shared by many crustaceans and hexapods) into a bipartite cone with two accessory cells. We have not coded for all variations in cone numbers (e.g., five-part cone in some Phyllopoda). Reduction of processes of crystalline cone-producing cells: 0, all cells have processes that pass through clear zone and rhabdom; 1, only accessory cells have processes. Richter (1999) observed 66. 67. 68. 69. 70. 71. 72. 73. 74. 75. 76. 77. that the malacostracan taxon Xenommacarida has processes of the cone cells restricted to the accessory cells, whereas these processes are present in all cone cells in other Malacostraca, in Branchiopoda, in Branchiura, and in insects. Distally displaced nuclei of accessory crystalline cone cells: 0, absent; 1, present. In Euphausiacea and Peracarida, nuclei of the accessory cells are displaced distally relative to the nuclei of the coneproducing cells, versus all nuclei positioned on top of the cone in Anaspidacea (Richter, 1999). Optic neuropiles: 0, no chiasmata; 1, one chiasma (between lamina ganglionaris and medulla); 2, two chiasmata (between lamina ganglionaris and medulla / between medulla and lobula). The presence of two chiasmata between the neuropiles in some malacostracans and in insects has been cited as evidence for a sister group relationship between these taxa (Osorio et al., 1995), or as defining a clade of Malacostraca, Remipedia, and Atelocerata (Moura and Christoffersen, 1996). However, the Leptostraca have only one chiasma in the optic lobe (Elofsson and Dahl, 1970), and Collembola have only two neuropiles (Paulus, 1979). Hexapod codings by Edgecombe et al (2000) are modified following descriptions by Strausfeld (1998) (chiasmata lacking in Machilidae; lobula absent but chiasma present between lamina and medulla in Lepisma and Thermobia). Lateral eye rhabdomes with quadratic network: 0, absent; 1, present (Weygoldt and Paulus, 1979; Dunlop and Webster, 1999). Star-shaped lateral eye rhabdomes are found in xiphosurans and scorpions, whilst they are quadratic in all other arachnids bearing lateral eyes (Dunlop and Webster, 1999), and arranged to form a "network" as described by Paulus (1979). The star-shaped arrangement is evidently plesiomorphic, based on its occurrence outside Chelicerata (e.g., Zygentoma: Paulus, 1979, fig. 6.13; Chilopoda, Paulus, 1986, fig. 3B). Number of median eyes: 0, none; 1, four; 2, two; 3, three. Paulus (1979) summarised evidence for four median eyes being a general condition in Euarthropoda. This number is reduced to two within Chelicerata. The lack of median eyes in Myriapoda has been interpreted as a synapomorphy (Boudreaux, 1979; Ax, 1999), whereas Kraus and Kraus (1994) cited the loss of median eyes as occurring independently in Chilopoda and in Progoneata. Inverted median eye: 0, absent; 1, present. In arachnids, the retina cells develop from an inverted invagination of the epidermis (Paulus, 1979). Median eyes fused to naupliar eyes: 0, absent; 1, present. Naupliar eyes are the median eyes of Crustacea, and the close association of the median eyes to form a functional unit has been proposed as a crustacean synapomorphy (Lauterbach, 1983; Weygoldt, 1986; Kraus, 1998). Naupliar eyes are not, however, present in all the crustacean taxa we have scored (e.g., peracarids; Cephalocarida: Elofsson and Hessler, 1990). This character is scored as inapplicable for myriapods (lacking median eyes). Type of sensory cells in naupliar eye: 0, inverse; 1, everse (Elofsson, 1966). Although Eloffsen (1966) dismissed pan-crustacean homologies in the naupliar eyes, Paulus (1979) did not regard the differences between those types with everse and inverse sensory cells as so fundamental as to disallow homology, and we concur. Tapetal cells in cups of naupliar eye: 0, absent; 1, present. Schram and Hof (1998) coded tapetal cells as present only in some maxillopodan crustaceans, and this pertains to Thoracica and Copepoda (Boxshall, 1992, table 1). They are located between the pigment and sensory cells (Elofsson, 1966). Dorsal frontal organs: 0, absent; 1, present. Elofsson (1965, 1966) regarded paired dorsal and ventral frontal organs as characteristic of Malacostraca, and the dorsal frontal organs as unique to malacostracans; they are, however, lacking in peracarids and leptostracans. Paulus (1972, 1979) suggested that homologous organs are present in Collembola but not other hexapods (e.g., frontal organs lacking in Archaeognatha and Tricholepidion fide Paulus). Frontal organs are entirely lacking in maxillopodans (Eloffson, 1966). Posterior medial frontal organ: 0, absent; 1, present. A posterior medial frontal organ is shared by Conchostraca (Spinicaudata) and Notostraca (Elofsson, 1966; Martin, 1992). Schram and Hof (1998) coded a distal frontal organ as another synapomorphy of Phyllopoda, though Elofsson (1966) stated that it is restricted to Conchostraca. Bec oculair: 0, absent; 1, present. Schram and Hof (1998) coded for this capsule in the malacostracan head that houses the naupliar eyes and frontal organs. Ocular tubercle: 0, absent; 1, present. Generally, the Opiliones present two central eyes situated on an ocular tubercle or ocularium. The ocular prominence can be lacking, or the eyes can be situated on separate tubercles. The ocular tubercle is absent in Cyphophthalmi (which lack median eyes), although it is present in some opilionid taxa that have secondarily lost their median eyes (e.g., Maiorerus and Hoplobunus). Thus, these two characters are coded as independent. A similar ocular tubercle to that of the Opiliones is found in Pycnogonida. Various Palaeozoic chelicerates possess a median eye tubercle to some degree (e.g., scorpions: Kjellsvig-Waering, 1986), leading Dunlop (1997, 1999) to consider it a potential autapomorphy for Chelicerata. The character coded here is a 78. 79. 80. 81. 82. 83. 84. 85. 86. more pronounced development than the low mound observed in many chelicerates. Even though certain laniatorids lack an eye tubercle, all the species for which molecular data are available present the eye tubercle, and it thus has been coded as present for the supraspecific terminal. Trichobothria innervated by several sensory cells, with dendrites having only indirect contact with the hair base: 0, absent; 1, present. Trichobothria (=bothriotrichs) are complex mechanoreceptors developed in several terrestrial arthropod groups. Reissland and Görner (1985) provided criteria for distinguishing the trichobothria of arachnids and myriapods from the ‘thread-hairs’ of insects, and we follow them by coding for non-homology with insect thread-hairs. Coding presence of trichobothria as apomorphic for extant Tetrapulmonata follows Dunlop (1996a: character 16). Selden et al. (1991: character 38) recorded trichobothria in extant scorpions, pseudoscorpions, and mites as well, and cited their absence in solifuges, ricinuleids, and opilionids. Trichobothria are lacking in early fossil scorpions (Jeram, 1998). Detailed structural similarities of myriapod trichobothria are coded as character 79. Basal bulb in trichobothria: 0, absent; 1, present. Trichobothria have distinctive modifications in polyxenid millipedes, pauropods and symphylans, notably a hair that forms a basal bulb (Haupt, 1979). Despite the variable position of such trichobothria on the body, the basal bulb has been proposed as a synapomorphy of Progoneata (Kraus and Kraus, 1994). This hypothesis requires that loss of trichobothria is a reversal in chilognathan millipedes (Enghoff, 1984). Oral papillae with slime glands and adhesive glands: 0, absent; 1, present. Oral papillae and their associated glands are unique to Onychophora (Brusca and Brusca, 1990). The slime glands may be modifications of the crural glands (character 297) that are developed in a variable number of legs (Storch and Ruhberg, 1993). Head/mouth orientation: 0, head prognathous, mouth directed anteroventrally; 1, head hypognathous, mouth directed ventrally; 2, mouth directed posteriorly. A hypognathous orientation of the head, with the mouthparts being directed downwards instead of forwards, is coded by Bitsch and Bitsch (2000: character 3) in Ectognatha and in some Collembola. A posteriorly directed mouth has been proposed as a characteristic feature of the TCC or schizoramian group (Cisne, 1974). This condition is present in xiphosurids, whereas the anteroventral orientation of the mouth is regarded as an arachnid synapomorphy (Shultz, 1990). The latter is acquired in ontogeny, modified from a posteriorly-directed state in the embryo of arachnids. The opening of the mouth on the proboscis in pycnogonids is not plausibly homologised with its opening to the preoral chamber in arachnids, and we have coded the state in pycnogonids as uncertain. In crustaceans, the esophagous has a posterior (Hutchinsoniella: Hessler and Elofsson, 1992, fig. 2) or posteroventral (e.g., Nebalia: Dahl and Wägele, 1994, fig. 365; Calanus: Razouls, 1994, fig. 189) orientation. Labrum: 0, absent; 1, present, originating from bilobed anlage. Partial covering of the mouth by a labrum is observed in all euarthropods except for the pycnogonids (Sharov, 1966; King, 1973). Scholtz (1998) recognised a labrum originating from bilobed anlage as a euarthropod synapomorphy. Fleshy labrum: 0, absent; 1, present. Walossek and Müller (1990) recognised an apomorphic character complex in the mouth region of crown-group Crustacea. This consists of a fleshy labrum which forms the cover of the atrium oris, with setulate, brush-like sides. A sternum with humped paragnaths is also part of the crustacean labral complex as defined by Walossek and Müller (1990). Entognathy (overgrowth of mandibles and maxillae by cranial folds): 0, absent; 1, present. Entognathy in the broad sense (mouthparts overgrown by cranial folds) occurs to varying degrees in onychophorans, pauropods and chilopods, as well as the hexapodan Entognatha (Manton, 1964). This character is scored to recognise more detailed similarities of the latter (Bitsch, 1994:114, and references therein; Kluge, 1999). The oral folds in Entognatha are joined together ventrally and are united behind with the postlabium. Admentum differentiated latero-ventrally on each side of head capsule, developed from posterior part of mouth fold: 0, absent; 1, present. Koch (1997) considered details of entognathy in Ellipura and Diplura to be fundamentally different, and disputed a single origin of the entognathous condition in Hexapoda (contra character 84). A unique sclerite, the admentum, is present in Diplura, intercalated between the prementum and oral fold (Koch, 1997; Bitsch and Bitsch, 2000). Ikeda and Machida (1998) described the origin of the admentum from the posterior part of the mouth fold (from the maxillary tergum?) as unique to Diplura. Coding is restricted to arthropods that can be meaningfully described as possessing a head capsule. Sclerotic sternum formed by antennal to maxillulary sternites: 0, absent; 1, present. Fusion of particular cephalic sternites into a sclerotic sternum is shared by the crown group of Crustacea (Walossek and Müller, 1998a), and is more generally shared with the fossil taxon Phosphatocopina according to Walossek (1999). Shultz (1990) regarded an unsegmented intercoxal plate in the prosomal sternum (the endostoma of Xiphosura) as the plesiomorphic state for arachnids. Neither the segmental composition nor morphology of this plate suggest homology with fusion of sternites in crustaceans. 87. Tritosternum: 0, absent; 1, present (Shultz, 1990: character 10). Coding is restricted to Chelicerata, wherein homologies of prosomal sternites can be identified. The tritosternum ("labium") is an anteroventrally projecting sternite in some arachnid orders. 88. Clypeolabrum and labium mobility: 0, free; 1, immobile. Kukalová-Peck (1991) identified immobility of the clypeolabrum and labium as a shared derived character of the Ellipura. 89. Hypopharynx: 0, absent or only a median lingua; 1, complete hypopharynx consisting of lingua and paired superlinguae. Coding is restricted to hexapods and myriapods, and follows Bitsch and Bitsch (2000: character 15). The possibility of homology between superlinguae and paragnaths in Crustacea (Crampton, 1921; Snodgrass, 1952; Bitsch and Bitsch, 2000) requires substantiation before coding is extended to crustaceans. Walossek and Müller (1998) indicate that paragnaths originate on the mandibular sternite, and Tiegs (1940) considered the superlinguae of Symphyla to likewise develop on the mandibular sternum, and to have mandibular innervation. The median apical lobes of the gnathochilarium of pauropods arise from the mandibular segment (Tiegs, 1947; Snodgrass, 1952), and are considered homologous with superlinguae. Ephemeroptera is coded based on the superlinguae of nymphs (Bitsch and Bitsch, 2000). 90. Fulturae: 0, absent or limited to a hypopharyngeal suspensorium; 1, present. The fulturae of entognathous hexapods and Archaeognatha are paired, cuticular thickenings in the lateral wall of the hypopharynx that extend from the bottom of the groove separating the mandibular and maxillary segments (Bitsch and Bitsch, 2000: character 19). Fulturae are represented in Myriapoda by paired hypopharyngeal processes that are fused with parts of the anterior tentorial apodemes (Kluge, 1999; Bitsch and Bitsch, 2000) (see character 91). Homologues of fulturae have not been documented in Crustacea, and absence is coded. 91. Hypopharynx supported by fultural sclerites that bear the head apodemes: 0, absent; 1, present. Snodgrass (1952) cited similarities of fultural sclerites of the hypopharynx as a "strong point in evidence of a relationship" between Diplopoda, Pauropoda and Chilopoda. In each case the fulturae support the apodemes that give rise to mandibular adductor muscles. We code the hypopharnygeal fulturae of Myriapoda as a character independent of the style of mandibular adbuction by movements of the tentorium (see character 126); one character involves the topological relationships of the hypopharynx, fulturae and apodemes, whilst the other involves movements of the apodemes relative to the mandible. Symphyla possess the head apodemes that serve as the attachments of the mandibular adductors, but lack fultural sclerites (Snodgrass, 1952). Snodgrass interpreted the fultural plates as the premandibular sternal sclerites of Myriapoda, and noted the absence of corresponding plates in Crustacea and Insecta. 92. Triradiate pharyngeal lumen: 0, absent; 1, present. Dewel and Dewel (1998) suggested that the triradiate pharynx of tardigrades may be plesiomorphic, being similarly developed in non-arthropod ecdysozoans. This argument has been elaborated by Schmidt-Rhaesa et al. (1998), who illustrated a triradiate lumen in Onychophora and nematodes. We code the "Introverted Y" pharynx of pycnogonids (Schmidt-Rhaesa et al., 1998) as triradiate. 93. Three-branched epistomal skeleton supporting the pharyngeal dilator muscles: 0, absent; 1, present. Scorpiones and Opiliones share attachments of the extrinsic pharyngeal muscles on three processes (one median and two lateral) on the epistome (Shultz, 2000). We code Opiliones based on Shultz’s data for Leiobunum, and code for Laniatores based on Gnidia. 94. Stomothecae: 0, absent; 1, present. Shultz (1990) defined stomothecae as expanded coxal endites that form the wall of the pre-oral chamber in some arachnids (here scorpions and opilionids). Jeram (1998) and Weygoldt (1998) interpreted Palaeozoic scorpions as lacking stomothecae, and questioned the homology of these endites, but Shultz (2000) provided considerable evidence for a unique structure of the coxapophyses in Opiliones and Scorpiones. Coding is restricted to arachnids. 95. Post-cephalic filter feeding apparatus with sternitic food groove: 0, absent; 1, present. Walossek (1993, 1995) emphasised a character complex associated with filter-feeding as an apomorphy of the Branchiopoda. 96. Antennal rami: 0, uniramous; 1, multiramous. Multiramy, defined as two or more rami attached distally to distinct basal podomeres (peduncle), is found in the Malacostraca, including the Leptostraca, in which the first antenna is biramous (developed as a scale), and the Stomatopoda, where it is triramous. The later condition is considered to be derived from the basal malacostracan biramous state (Kunze, 1983). Olesen and Walossek (2000:61) cited the development of the outer branch (scale) in Nebalia relatively late in ontogeny as evidence for biramy being an apomorphy in Malacostraca. The Remipedia have biramous first antennae, but lack defined basal podomeres, and their modification of the first head limb is not considered homologous with that of the malacostracan first antenna. 97. Size of the first antennae: 0, long, flagelliform; 1, only few segments. Branchiopoda have highly reduced first antennae, while most other crustaceans, myriapods and hexapods have long, flagelliform first antennae. 98. Antennal apical cone sensilla: 0, absent; 1, present. Unique to Diplopoda is a cluster of cone-shaped sensilla on the distal antennomere of the antenna (Enghoff, 1984). The cluster usually consists of four sensilla. 99. Intrinsic muscles of antennae: 0, present; 1, absent (flagellum). As recognised by Imms (1939), the flagelliform, unmusculated antenna unites Insecta (N.P. Kristensen, 1981, 1991, 1998), though Scutigeromorpha have the same structure. We have not coded flagellate biramous first antennae in Crustacea (e.g., Anaspides, many Reptantia) as homologous. 100. Scape and pedicel differentiated in antennae, with Johnson’s organ: 0, absent; 1, present. Presence of Johnson’s organ in the pedicel serves to distinguish insects from other arthropods that have an unmusculated antennal flagellum. 101. Sclerotised bridge between antennae: 0, present; 1, absent. Absence of a sclerotised bridge between the antennae is distinctive for Geophilomorpha within the Chilopoda (Dohle, 1990) (see Edgecombe et al., 1999: character 4 for discussion). 102. Antennal circulatory vessels: 0, antennal vessels joined with dorsal vessel; 1, antennal and dorsal vessels separate; 2, antennal vessels absent. Pass (1991) provided a review of antennal circulatory vessels in arthropods. Insecta are defined by the separation of the antennal vessels and the dorsal vessel, versus the connection of the dorsal vessel and antennal vessels in Diplura, Myriapoda and Crustacea. Collembola and Pauropoda lack antennal vessels. 103. Ampullo-ampullary dilator and ampullo-aortic dilator muscle: 0, absent; 1, present. Antennal pulsatile organs (ampullae) are present in most orders of pterygotes (see Pass, 2000, fig. 3 for summary). Of terminals scored here, Blattodea, Orthoptera and Diptera share pumping muscles for the pulsatile organs, in particular an ampullo-ampullary dilator and accessory dilators attached to the anterior end of the aorta. Ephemeroptera lack ampullae, either pulsatile or non-pulsatile (Pass, 1991). This and other taxa lacking pulsatile ampullae are scored as inapplicable. 104. Statocyst in basal segment of first antenna: 0, absent; 1, present. Richter and Scholtz (2001: character 56) followed Siewing (1956, 1963) in scoring for the presence of an antennal statocyst as a shared derived character in Anaspidacea and Decapoda (absent in other Malacostraca coded here). A statocyst on the same article is present in stomatopods (Tirmizi and Kazmi, 1984). 105. Appendage on third (tritocerebral) head segment: 0, unspecialised; 1, antenna; 2, intercalary appendage absent; 3, chelifore or chelicerae. Several major groups of arthropods are defined by conditions of the appendage of the third metamere. The plesiomorphic state is that observed in fossil groups such as trilobites, in which this post-antennal limb is undifferentiated from other cephalic limbs (or, for that matter, from trunk limbs). Crustaceans uniquely possess a second antenna, atelocerates have suppressed this somite (such that embryonic limb buds in some taxa are its maximal expression: Anderson, 1973), and chelicerates have the chelicera in this position. Certain pycnogonids (e.g. Colossendeis) lack chelifores, others (e.g. Endeis) lack chelifores in the adult stage although they are present in the juveniles, while others (e.g. Ammotheidae) have chelifores throughout ontogeny. Ammotheidae and Endeis have been coded for the presence of chelifores. 106. Cheliceral segmentation: 0, three segments, the last two forming a chela; 1, two segments, subchelate, "clasp-knife" type (Shear et al., 1987; Shultz, 1990; Dunlop, 1996). 107. Plagula ventralis: 0, absent; 1, present (Shear et al., 1987). Homann (1985) claimed that a small sclerite in the articular membrane between the fang and basis of the chelicera (the plagula ventralis) is unique to Tetrapulmonata. It is not present in all Araneae (Dunlop, 1996a). 108. Cheliceral tergo-deutomerite muscles: 0, absent; 1, present. A muscle from the carapace that inserts on the second cheliceral article is shared by Opiliones (Opilio coded based on Leiobunum: Shultz, 2000) and Scorpiones. Shultz (2000) cited its absence in Pycnogonida, Limulus and Eukoenenia (Palpigradi). Coding is restricted to chelicerates with three cheliceral articles. 109. Second antennal exopod: 0, present; 1, absent. Fossil arachnomorphs (e.g., trilobites) demonstrate that a biramous second cephalic limb is not restricted to Crustacea (as would appear to be the case based on extant taxa alone). In some crustacean taxa, the exopod is expressed only as a scale (Stomatopoda and Eumalacostraca; see character 110) while others have a more general flagellate exopod. The Notostraca and Anostraca have an antennal exopod in their larval stages, but not as adults. Other branchiopods, however, have biramous antennae as adults. 110. Single segmented antennal scale: 0, absent; 1, present. A single-segmented antennal scale (versus two-segmented in Hoplocarida) was used by Schram (1986) as a synapomorphy of Eumalacostraca. It is primitively present in Peracarida (e.g. in Mysidacaea: Dahl, 1983), though is absent in the Oniscidea coded here. Brusca and Wilson (1991) questioned homology between the scale-like exopods of asellote isopods and those of other malacostracans. Coding is restricted to taxa that possess an antennal exopod (see character 109). 111. Antennal naupliar protopod: 0, short; 1, long. Sanders (1963) contrasted the length of the antennal protopod in branchiopod nauplii with that in other crustaceans. The branchiopod condition (state 1) may be characterised as a protopod exceeding 50 percent of the length of the naupliar antenna (Schram and Hof, 1998). 112. Distal-less expressed in mandible: 0, present (including transient expression in embryo and in palp); 1, absent in all ontogenetic stages. Manton’s (1964) argument that atelocerate mandibles are the tips of ‘whole limbs’ whereas crustacean mandibles arise as coxal gnathobases has been recast (Panganiban et al., 1995; Popadić et al., 1996) and then rejected (Popadić et al., 1998; Scholtz et al., 1998) by work on Distal-less expression, which indicates that mandibles are uniformly gnathobasic. As in other characters involving gene expression, taxonomic sampling is limited. Glomeris is coded as a proxy for Epicyliosoma. Rather than coding inapplicability for Chelicerata, the high, continuous expression of Distal-less in all prosomal limbs of the spider Achaearanea (coded as a proxy for Mygalomorphae; Popadić et al., 1998) is used as evidence that state 0 pertains to non-mandibulates. The absence of Distal-less expression in all ontogenetic stages pertains only to Hexapoda [coded for Collembola (Arthropleona), Zygentoma (Lepismatidae) and Drosophila based on Popadić et al. (1998) and Scholtz et al. (1998); Locusta coded based on cricket data cited by Popadić et al. (1998) and Gryllus (Niwa et al., 1997)]. 113. Mandible (gnathobasic appendage of third limb-bearing metamere is main feeding limb of adult head): 0, absent; 1, present. Snodgrass (1950) and Manton (1964) assumed opposite positions on the significance of mandibles in arthropods. The former emphasised their fundamental similarity between crustaceans and atelocerates, whereas the latter regarded their differences to violate the possibility of a single origin. Defences of the homology of mandibles are offered by Weygoldt (1986) and Wägele (1993), the latter regarding the embedding of the mandible between the labrum and hypopharynx to form a "chewing chamber" as evidence for homology. The pattern of reduced Distalless expression through ontogeny that is observed in the mandibles of crustaceans and myriapods (expression completely suppressed in hexapods) reinforces the homology of mandibles (Popadić et al., 1998). 114. Mandibular base plate forming side of head: 0, absent; 1, present. Mandibular structure and function are similar in Symphyla and Diplopoda (Snodgrass, 1950). In each of these groups, the proximal part of the mandible, the base plate, is a prominent component in the side of the head capsule, such that muscular abduction of the mandible is abandoned (Manton, 1979a). Instead, the mandible abducts by the anterior tentorial apodeme (‘swinging tentorium’) pushing on the sides of the gnathal lobes. This style of abduction in symphylans and diplopods is associated with a greater degree of muscular independence of the gnathal lobe than in chilopods. This and other characters pertaining to mandibular structure are scored as inapplicable for Drosophila. 115. Telognathic mandible with musculated gnathal lobe, flexor (anterior dorsal muscle) arising dorsally on the cranium: 0, absent; 1, present. Staniczek (2000) critiqued the arguments of Kraus and Kraus (1996), Kraus (1998) and Kukalová-Peck (1998) that hexapods have segmented/telognathic mandibles, and concluded that gnathobasic mandibles are general for Mandibulata (see character 112). This implies "a secondary subdivision of the mandible in the myriapod lineages" (Staniczek, 2000:176), as concluded previously by Lauterbach (1972). Regardless of the status of "telognathy" in hexapods, the structural differentiation of myriapod mandibles can be characterised with apparently apomorphic details. Chilopoda (except Geophilomorpha) resemble Diplopoda and Symphyla in having the gnathal lobe of the mandible musculated by a large flexor that arises on the dorsal surface of the cranium (Snodgrass, 1950, 1952). The single-piece mandible of pauropods (except Hexamerocerata; Enghoff, 1990; Kraus and Kraus, 1994) lacks this differentiation of a gnathal lobe. The dorsal muscles of hexapods and crustaceans do not serve as gnathal lobe flexors. Kluge (1999) cited the division of the mandible into two movably jointed sclerites (i.e., gnathal lobe and base) with the anterior dorsal muscle serving as an adductor as a myriapod synapomorphy. 116. Second (anterior) mandibular articulation with the cranium, movement limited to transverse adduction around a horizontal axis of swing: 0, absent; 1, present. N.P. Kristensen (1975, 1981), among many others, noted that a dicondylic articulation of the mandible defined a clade uniting all insects to the exclusion of Archaeognatha. Observations by Fürst von Lieven (2000) and Staniczek (2000) strengthen the homology of the anterior articulation, e.g., the posterior part of the articulation complex is formed by a processus paratentorialis located ventral to the anterior tentorial pit. 117. Ball-and-socket mandibular articulation: 0, absent; 1, present, formed between clypeal condyle and mandibular ridges. Fürst von Lieven (2000) and Staniczek (2000) both defended the apomorphic status of the ball joint that forms the secondary articulation of the mandible in opthopteroid insects (putative synapomorphy for Odontata + Neoptera). Coding is restricted to Dicondylia (dependent on the presence of a second mandibular articulation). 118. Mandibular scutes: 0, absent; 1, present (mandible composed of 2-5 moveable scutes). The chilopod mandible is unique, even among to other myriapod taxa with segmented mandibles (symphylans and diplopods), in that it is fragmented into a series of scutes/laminae. This structure has been posited as a synapomorphy of Chilopoda (Boudreaux, 1979). 119. Mandibular palp: 0, present (appendage of third limb-bearing cephalic metamere with telopod); 1, absent throughout ontogeny; 2, present in larva, absent in adult. The lack of a mandibular palp is frequently evoked as a synapomorphy of Atelocerata by workers who have defended a monophyletic Mandibulata (e.g., Weygoldt, 1986; Kraus, 1998). The presence or absence of a mandibular palp is, however, variable within Crustacea. We recognise three states to account for ontogenetic change in the presence or absence of a mandibular palp in crustaceans. The coding for remipedes (palp absent in adult) is uncertain because of the lack of larval data, and thus has been coded as either 1 or 2 (but not 0). Machida (2000) demonstrated the absence of a palp throughout embryology in Archaeognatha. 120. Lacinia mobilis on adult mandible: 0, absent; 1, present. Moura and Christoffersen (1996) considered the presence of a lacinia mobilis to define a remipede-malacostracan-atelocerate clade. Edgecombe et al. (2000: character 67) critiqued this interpretation because the lacinia mobilis is not present in all malacostracans and hexapods. Bitsch and Bitsch (2000: character 14) recorded its presence in Symphyla, but documentation is needed. Homologues in Diplopoda have not, to our knowledge, been identified, and we have scored diplopods as lacking a lacinia mobilis. The internal and external tooth of Chilognatha (Enghoff, 1979; Köhler and Alberti, 1990) might be compared to incisor processes, but these teeth are lacking in polyxenids (Ishii, 1988; Ishii and Tamura, 1995). The basic similarity of the lacinia mobilis in remipedes, peracarids and hexapods (the latter being the prostheca: coded following Bisch and Bitsch, 2000: character 14) warrants coding. Ephemeroptera is scored for the nymph. 121. Posterior tentorial apodemes: 0, absent; 1, tentorial arms; 2, metatentorium. Posterior tentorial apodemes are lacking in myriapods. Manton (1964) regarded the anatomy, connections, and associated muscles of the posterior tentorial arms in Collembola and Diplura as indicating homology with the fused posterior tentorial bar (metatentorium) of Insecta. Koch (2000) defended this homology. An ordered coding recognises the fused state (metatentorium) as a modification of separate tentorial arms. 122. Pre- and metatentorium fused: 0, absent; 1, present. Fusion of tentorial components in Pterygota (N.P. Kristensen, 1991; character 11 of Koch, 2000) is codable for Insecta only. 123. Anterior tentorial arms: 0, absent; 1, cuticular tentorium developing as ectodermal invaginations; 2, complex connective endosternite; 3, cuticular fulcro-tentorium. The tentorial cephalic endoskeleton is restricted to myriapods and hexapods. Snodgrass (1950) regarded the head apodemes of Myriapoda as homologous with the anterior tentorial arms of Insecta, in which they likewise arise as cuticular (ectodermal) invaginations; Bitsch and Bitsch (2000), Koch (2000) and Klass and Kristensen (2001) noted the probable homology of these structures, though the morphological details of myriapods warrant further observations. Some workers (see Matsuda, 1965 for summary of early accounts) have considered homologous elements to be lacking in the Entognatha. Bitsch and Bitsch (2000: character 20) coded the fulcro-tentorium of Protura (here state 3) as non-homologous with the true tentorium of Insecta (state 1), and interpreted the endoskeletal formations of Collembola and Diplura to be a complex endosternite composed of connective fibers (state 2) rather than a cuticular tentorium. Koch (2000), however, endorsed homology between the anterior tentorial apodemes of Collembola, Diplura and Insecta, citing common points of origin, e.g., identical sclerotic connections with the labrum. 124. Posterior suspension of anterior apodemes to cranial wall: 0, absent; 1, present. Coding of this character in Diplura is dependent upon the homology of the anterior apodemes in that group and the anterior tentorium of Insecta (Koch, 2000: character 5) (see discussion of character 123). 125. Anterior tentorium: 0, absent (separate rod-like anterior tentorial apodemes); 1, anterior part of tentorial apodemes forms arched, hollow plates that approach each other mesially but remain separate; 2, anterior tentorium an unpaired roof. An anterior tentorium developed as a transverse roof (state 2) is present in Dicondylia (Koch, 2000: character 10). An ordered coding regards it as formed by coalescence of the similarly plate-like anterior part of the tentorial apodemes of Archaeognatha (state 1). 126. Swinging tentorium: 0, absent; 1, present. Abduction of the mandible in most Myriapoda is achieved by movements of the anterior apodemes/tentorial arms (Manton, 1964). This condition is unique to myriapods. Mandibular movements are, however, solely muscular in Geophilomorpha and Scutigeromorpha (Manton, 1965). 127. Suspensory bar from mandible: 0, absent; 1, present. An articular rod (Snodgrass, 1950) or suspensory bar (Boudreaux, 1979) is present in the posterior mandibular attachment in all orders of chilopods except the Geophilomorpha. A similar rod, a thickening of the lateral pouch wall, forms the posterior support of the mandible in Ellipura (Snodgrass, 1950). 128. Intergnathal connective lamina: 0, present; 1, absent. Codings for hexapods and for myriapod outgroups follow Bitsch and Bitsch (2000: character 21). Their "intergnathal connective lamina" refers to the transverse ‘tendon’ linking the mandibular and maxillar adductors in myriapods and basal hexapods (including Tricholepidion; Kristensen, 1998; Staniczek, 2000) but not in other Zygentoma or in Pterygota. Presence of the intergnathal lamina is plesiomorphic for myriapods and hexapods, as the obvious homologue is present in Crustacea ("intergnathal ligament" in Anaspides: Snodgrass, 1952, fig. 38H; Nebalia: Dahl and Wägele, 1994, fig. 354). The inferred loss of this tendon is associated with the origin of the ventral tentorial adductor muscle on the anterior tentorial arm (versus its origin on the tendon) (Staniczek, 2000). 129. Mandibulo-hypopharyngeal muscle: 0, absent; 1, present. Staniczek (2000) observed that mandibulo-hypopharyngeal muscle M8 of lepismatids, nicoletiids and pterygotes, which probably serves to protract the hypopharynx, is lacking in Tricholepidion. He considered absence to be plesiomorphic, based on the lack of a homologous muscle in Archaeognatha, myriapods and entognathous hexapods. We have coded myriapods and entognathans as a groundpattern (absence) based on these data. 130. Complete postoccipital ridge: 0, absent; 1, present. Snodgrass (1935) described a postoccipital ridge as the internal aspect of the postoccipital suture, which is often produced into apodemal plates on which are attached the dorsal prothoracic and neck muscles of the head. A complete postoccipital ridge (N.P. Kristensen, 1981) or dorsally-complete postoccipital suture (Kristensen, 1998) is cited as a synapomorphy of Dicondylia. Staniczek (2000) substantiated this homology. 131. Ovigers: 0, absent; 1, present (King, 1973; Arnaud and Bamber, 1987). A pair of "appendages" attached to the ventral surface of the body, posteriorly situated in relation to the palps, is present at least in the males of all the pycnogonid species here analyzed. 132. Salivary glands: 0, absent; 1, arise as ectodermal invaginations on second maxilla/labium; 2, arise as mesodermal segmental organs of the first maxillae. Anderson (1973) recognised two patterns of development for salivary glands in hexapods and myriapods. In chilopods and hexapods, the salivary glands form as ectodermal invaginations of the second maxillary or labial segment. In Progoneata, salivary glands are mesodermal segmental organs of the maxillary segment. 133. Opening of maxillulary salivary glands: 0, pair of openings at the base of the second maxillae; 1, one median opening in the midventral groove of the labium; 2, one median opening in the salivarium, between the labium and hypopharynx. Bitsch and Bitsch (1998: character 8) identified distinctive positions of ducts of the salivary glands in Entognatha (state 1) and Insecta (state 2). Coding is restricted to those taxa with salivary glands of ectodermal origin on the second maxillae/labium (chilopods and hexapods; character 132, state 2). 134. Maxillae on fourth limb-bearing metamere: 0, absent; 1, present. All extant mandibulates have the appendage of the fourth metamere specialised as a mouthpart, a maxilla. This character is sometimes combined with maxillary development on the succeeding limbs as a mandibulate synapomorphy (e.g., Brusca and Brusca, 1990, among others). Discussion under character 143 indicates that first and second maxillae must be evaluated separately. 135. First maxillary precoxal segment: 0, absent; 1, present. Boxshall (1998) suggested that definition of a precoxa in the maxillulary protopod is restricted to the Remipedia and taxa traditionally grouped as Maxillopoda, including the Copepoda. 136. First maxilla with medially-directed lobate endites on the basal podomere, possibly consisting of a precoxa and coxa: 0, no lobate endites; 1, two endites; 2, one endite. Crustaceans are unique in the presence of medially-directed lobate endites on a number of appendages. Boxshall (1998) summarised information on endite configurations on the crustacean first maxilla (maxillule). Separate states are recognised for malacostracans (one basal endite) and for the cephalocarids, remipedes and mystacocarids (two basal endites). Extant branchiopods are coded as inapplicable (-) due to the reduction of the maxillulae, but fossil taxa such as Rehbachiella (Walossek, 1993) indicate that the plesiomorphic branchiopodan state is 2. Walossek (1993, 1999) recognised this similarity in maxillulary endite configurations as grounds for separating Entomostraca and Malacostraca. 137. Maxillary plate [basal parts of fifth limb-bearing metamere (second maxilla or labium) medially merged, bordering side of mouth cavity]: 0, absent; 1, present. Kraus and Kraus (1994) cited this morphology as a synapomorphy for Labiophora (Progoneata + Hexapoda = Labiata of Snodgrass, 1938). They contrasted it with the situation in chilopods, in which the first maxillae border the mouth. The maxillary plate corresponds to Snodgrass’ (1938) concept of a labium, which he also regarded as synapomorphic for Labiophora. Kraus and Kraus’ (1994, 1996) argument is dependent on their interpretation that the diplopod and pauropod gnathochilarium is composed of two pairs of appendages, first and second maxillae, a claim developed earlier by Verhoeff, and upheld by Kraus and Kraus based on external morphology. Dohle’s (1998) counterarguments, including the complete lack of limbs on the mx2 segment in embryos, innervation by a single pair of ganglia, and muscles being those of a single segment, are used in our codings of maxillary characters. Dohle (1998) concluded that the lower lip of Dignatha is composed of the appendages of the first maxillary segment and the intervening sternite alone. Scholtz et al. (1998) strengthened Dohle’s interpretation by demonstrating the lack of Distal-less expression on the postmaxillary segment in diplopods. 138. First maxillary palps: 0, present (telopod present on appendage of fourth metamere); 1, absent. The absence of maxillary palps has been cited as a synapomorphic character for Progoneata (Kraus and Kraus, 1994; Kraus, 1998). An ambiguity, however, is the interpretation of the minute lateral cone on the so-called stipes of symphylans, which has been cautiously homologised with a vestigial palp (e.g., Snodgrass, 1952). The evidence is not especially compelling, and we have coded the palp as absent in Symphyla. Presence of a palp in polyxenids follows Shear (1998). 139. Hypertrophied maxillary palp: 0, absent; 1, present. Kristensen (1998) regarded a maxillary palp larger than the thoracic locomotory limbs as a possible autapomorphy for the Archaeognatha. This character recognises the size of the palp rather than its pronounced segmentation. 140. First maxilla divided into cardo, stipes, lacinia, and galea, with similar musculation and function: 0, absent; 1, present. Manton (1964) observed the structure and function of the maxilla to be similar throughout the Hexapoda, and there is little question that details are apomorphic (see Kluge, 1999). While the same descriptive terminology is employed for the units of the first maxilla in symphylans, Manton noted significant differences in details of musculature and function from those in hexapods. Characteristic of the hexapod maxilla are the cardo bearing a strong point of articulation with the cranium, the principal retractor-adductor to the lacinia inserting on the cranium, and muscle XI (following the homology scheme of Manton, 1964) from the cardo being a principal protractor. Tuxen’s (1959) study of Diplura concluded that the structure of the maxilla in this group was most similar to that in Ellipura, but also noted that the entognathan maxilla closely resembled that of other hexapods. 141. Interlocking of galea and superligua: 0, absent; 1, present. Kristensen (1998, fig. 21.2D) documented a distinctive interlocking of the galea and superligua as a possible dipluran synapomorphy (also Bitsch and Bitsch, 2000: character 16). Koch (1997) likewise cited connection/partial fusion of the maxilla and hypopharynx as a unique aspect of entognathy in Diplura. The character is scored only for Hexapoda because homologues of the galea and superligua are lacking in other arthropods. 142. Gnathochilarium including intermaxillary plate: 0, absent; 1, present. Pauropods and diplopods have classically been united based on the maxillary structures, developed as a gnathochilarium. Kraus and Kraus (1996) have, however, disputed the view that pauropods and polyxenid diplopods possess a true gnathochilarium. Our coding recognises the traditional hypothesis. 143. Second maxillae on fifth metamere: 0, appendage developed as trunk limb; 1, well developed maxilla differentiated as mouthpart; 2, vestigial appendage; 3, appendage lacking. Second maxillae are lacking in some crown-group Crustacea (notably Cephalocarida), in which the corresponding appendage may retain the structure of a trunk limb. Suppression of the second maxilla, being largely a pedestal for the maxillary gland (state 2), is coded for branchiopods. The complete absence of limbs on the second maxillary segment is shared by pauropods and diplopods (following Dohle, 1998; see discussion of character 137). 144. Egg tooth on second maxilla: 0, absent (no embryonic egg tooth on cuticle of fifth limb-bearing metamere); 1, present. Dohle (1985) proposed that an egg tooth on the second maxilla is an autapomophy of Chilopoda. 145. Coxae of second maxilla medially fused: 0, absent (coxae of fifth metamere not fused); 1, present. Pleurostigmophoran chilopods share a medial fusion of the coxae of the second maxilla, a condition regarded as synapomorphic (Dohle, 1985; Shear and Bonamo, 1988). 146. Symphylan-type labium: 0, absent; 1, present. The medial fusion of the second maxillae to form a functional labium is cited as a symphylan synapomorphy (Kraus and Kraus, 1994, 1996). Features that distinguish the symphylan labium from the hexapod labium include the anterior plate with a row of papilla-bearing lobes distally and tapering proximal arms that extend back to a pair of cervical sclerites (Snodgrass, 1952). 147. Linea ventralis: 0, absent; 1, present. Kristensen (1991) and Kraus (1998) postulated that the median groove in the posterior/ventral surface of the head in Ellipura, the linea ventralis, was a synapomorphy. It extends from the openings of the labial glands to the neck membrane in Protura and back to the preabdominal ventral tube in Collembola. 148. Divided glossae and paraglossae: 0, undivided pair of glossae and paraglossae; 1, glossae and paraglossae bilobed. Kristensen (1991) noted that bilobed ligular elements were a "peculiarity" of the archaeognathan labium, but was uncertain whether or not they provided an autapomorphy for Archaeognatha. Diplura is coded as missing data because the ligulae are fused. Protura is coded with the assumption that the inner and outer labial lobes (Matsuda, 1976, fig. 40B) are the glossae and paraglossae, respectively. Ephemeroptera is scored for the state of the ligulae in nymphs (Matsuda, 1976, fig. 49A), the character being inapplicable for adults. 149. Rotation of labial Anlagen: 0, absent; 1, present. Ikeda and Machida (1998) described the labium of Campodeidae developing with a peculiar rotation of its Anlagen, and interpreted the posterior movement of the labial palp Anlagen in Japyx to indicate the same rotation. 150. Widened apical segment of labial palp: 0, absent; 1, present. Kristensen (1998) proposed that a marked widening of the apical segment of the labial palp is a synapomorphy for Zygentoma, though Bitsch and Bitsch (2000) and Staniczek (2000) questioned its value based on widening of this segment in some Archaeognatha and larval Ephemeroptera, respectively. 151. Collum: 0, absent; 1, present. Diplopods possess a limbless first post-cephalic metamere, the collum. Pauropods have, at most, minute vestiges of limbs on this segment (Kraus and Kraus, 1994). 152. Collum covering posterior part of head capsule and part of segment II: 0, absent; 1, present. An enlarged collum is present in Juliformia (Enghoff, 1984). Coding is restricted to Dignatha (dependent on presence of a collum). 153. Direct articulation between first and fourth articles of telopodite of maxilliped: 0, absent (first and fourth articles of telopodite of sixth metamere lack a common hinge); 1, present. Characters 153-160 pertain to various conditions of the maxilliped in Chilopoda. States in other arthropods are based on the appendage of the homologous (sixth limb-bearing) metamere. The common hinge between the articles of the maxilliped telopodite is a classic character for Epimorpha (Scolopendromorpha and Geophilomorpha), as recognised by Attems (1926) and maintained in more recent cladistic analyses (Kraus, 1998 and references therein). 154. Coxosternite of maxilliped sclerotised in midline: 0, coxae separated medially, with sternite present in adult; 1, coxosternal plates meeting medially, with flexible hinge; 2, coxosternal plates meeting medially, hinge sclerotised and non-functional. Shear and Bonamo (1988) coded the condition of the maxilliped coxosternum in Chilopoda as a multistate character. They (1988:9) regarded the medially sclerotised condition shared by Craterostigmus and Epimorpha as apomorphic based on serial homology (fusion makes the maxillipeds less like the following trunk legs). Dohle (1990) drew the same interpretation of this state as a synapomorphy for that group, whereas Ax (1999) considered the separate coxosternal plates of Scutigeromorpha to be an autapomorphy. Scutigeromorpha have a sternite present in adults (Manton, 1965, fig. 91a) and coxae are separated medially, as in outgroups (Symphyla and Hexapoda). In Scolopendromorpha, the sternal contribution to the coxosternite is expressed only in early ontogeny (Manton, 1965:324). Medial coalescence (state 1: Lithobiomorpha) and sclerotisation of the hinge (state 2: Craterostigmus + Epimorpha) are thus coded as states of an ordered character, as suggested by Shear and Bonamo (1988). 155. Maxilliped coxosternite deeply embedded into cuticle above second trunk segment: 0, not embedded; 1, embedded. Manton (1965) identified features of the maxillipeds in Craterostigmomorpha and Epimorpha associated with strengthening their proximal attachment. Among these is the embedding of the coxosternum into cuticle beneath the succeeding trunk segment. 156. Maxilliped segment with pleurite forming a girdle around coxosternite: 0, small lateral pleurite; 1, large girdling pleurite. This character has been coded only for myriapods and hexapods (other arthropod taxa lack pleurites on the first trunk segment), the apomorphic state being restricted to certain chilopods. In Craterostigmorpha and Epimorpha, the pleurite of the maxilliped segment envelopes the coxosternite (Manton, 1965; =‘Spange’ of Attems, 1926). 157. Sternal muscles truncated in maxilliped segment, not extending into head: 0, sternal muscles extended into head; 1, sternal muscles truncated. Manton (1965) observed that the truncation of the sternal muscles in the maxilliped segment distinguishes craterostigmomorph and epimorph chilopods from scutigeromorphs and lithobiomorphs. The latter condition, with the sternal muscles extending into the head, is shared by pauropods and symphylans (Manton, 1966) and collembolans and proturans (Manton, 1972), and is thus regarded as plesiomorphic. Because of uncertainties in drawing homologies with muscles in crustaceans and chelicerates, this character is coded for myriapods and hexapods only. 158. Maxilliped tooth plate (anteriorly-projecting, serrate coxal endite): 0, absent; 1, present. The toothplate is employed by scolopendromorph centipedes as a ‘can opener’ in stabbing prey. The same type of endite is present in the Craterostigmomorpha (Manton, 1965; Dohle, 1990) and the Devonian Devonobius (Shear and Bonamo, 1988). 159. Maxilliped poison gland: 0, absent; 1, present. This character describes the modification of the first pair of trunk limbs in chilopods into a fang with a poison gland. 160. Maxilliped distal segments fused as a tarsungulum: 0, separate tarsus and pretarsus; 1, tarsus and pretarsus fused as tarsungulum. Borucki (1996) recognised the fused tarsungulum in the maxilliped of pleurostigmorphoran chilopods as a synapomorphy for that group, in contrast to the articulated tarsus and pretarsus in scutigeromorphs. 161. Oblique muscle layer in body wall: 0, absent; 1, present. An oblique muscle layer, with fibres organised in a chevron pattern (Storch and Ruhberg, 1993), is a specialisation of Onychophora. 162. Longitudinal muscles: 0, united sternal and lateral longitudinal muscles; 1, separate sternal and lateral longitudinal muscles, with separate segmental tendons. The division of the longitudinal muscles into separate sternal and longitudinal bands serves to unite craterostigmomorph, scolopendromorph and geophilomorph chilopods (Manton, 1965). Taxa lacking lateral longitudinal muscles (e.g., Campodeidae; Manton, 1972) have been coded as inapplicable. Coding is restricted to myriapods and hexapods. 163. Superficial pleural muscles: 0, absent; 1, present. Within the Chilopoda, Epimorpha and Craterostigmomorpha share details of pleural musculature. These include the presence of the superficial pleural muscles pam.1, pam.2 and ptm of Manton (1965). The homology of these with muscles in other arthropod groups is questionable, so we have only coded this character for Symphyla, Pauropoda and Polyxenidae (based on Manton 1966). 164. Crossed, oblique dorsoventral muscles: 0, absent; 1, present. Boudreaux (1979) interpreted the crossed, oblique dorsoventral muscles in crustaceans, myriapods and hexapods as a mandibulate synapomorphy. Codings for Chilognatha and Pterygota are entered as uncertain because the trunk musculature is greatly modified. Serial dorsoventral muscles are lacking in pycnogonids and onychophorans (Firstman, 1973), rendering this character inapplicable for those taxa. The dorsoventral suspensors of the endosternum and the abdominal dorsoventral muscles of chelicerates lack the crossed, oblique arrangement of mandibulates. 165. Deep dorsoventral muscles in the trunk: 0, absent; 1, present. Manton (1965) identified a complex of dorsoventral muscles (dvc muscles in her terminology) that pass upwards to the trunk tergites in epimorphic chilopods. They are absent in ‘anamorphic’ chilopods. As for character 163, a limited range of taxa are coded due to uncertain homologies. Polarity can, however, be provided by Manton’s (1966) observation that deep dorsoventral trunk muscles are lacking in Symphyla. 166. Circular body muscle: 0, present; 1, suppressed. Lack of circular body wall muscle is shared by tardigrades and euarthropods. We code the similarity without assuming that the former is due to miniaturisation (Dewel and Dewel, 1998). 167. Discrete segmental cross-striated muscles attached to cuticular apodemes: 0, absent; 1, present. Brusca and Brusca (1990) and Nielsen (1995) cited cross-striated muscles as a synapomorphy to unite tardigrades with euarthropods. 168. Abdominal muscles: 0, straight; 1, twisted. This character describes one aspect of abdominal modifications associated with jumping in Archaeognatha. Manton’s (1972) description of abdominal skeletomusculature in Petrobius noted such modifications as twisted, rope-like dorsal muscles and deep oblique muscles in the abdomen, and strong development of the abdominal tendon system. Modifications of the endoskeleton and tergum, such as greatly overlapping abdominal tergites that slide over one another dorsally, are part of the same character complex (Kristensen, 1998). Kukalová-Peck (1991) claimed that the abdominal rope muscles of archaeognathans are a plesiomorphy because they are "shared with Crustacea". This pertains only to the so-called caridoid musculature of some malacostracans. 169. Proventriculus in the foregut: 0, absent; 1, present. Malacostracans (including Leptostraca) share an expanded anterodorsal chamber, divided into a cardiac and a pyloric region adjacent to anterior caecae of the midgut (Dahl, 1987). Klass (1998) documented detailed similarities in the proventriculus of Reptantia and Zygentoma, including similar sclerites, denticles and apodemes. Based on these similarities, the proventriculus of malacostracans and insects warrants a hypothesis of primary homology. Coding for the distribution of the proventriculus in Hexapoda is as by Bitsch and Bitsch (1998: character 7). 170. Lateralia and inferolateralia in the cardiac chamber: 0, absent; 1, present (Richter and Scholtz, 2001: character 68). This and the following four characters pertain to variation in the morphology of the foregut of Malacostraca, with states defined and coded as by Richter and Scholtz (2001). These characters are inapplicable to all non-malacostracan crustaceans (which lack a proventriculus), and all non-crustaceans except for some insects (see character 169). Precise homologies of these states in the insect proventriculus have not been determined, and we have scored these taxa as uncertain. 171. Unpaired superomedianum at transition from cardia to pyloric chamber: 0, absent; 1, present (Richter and Scholtz, 2001: character 68). 172. Inferomedianum anterius (midventral cardiac ridge): 0, absent; 1, present (Richter and Scholtz, 2001: character 69). 173. Inferomedianum posterius (midventral pyloric ridge): 0, absent; 1, present (Richter and Scholtz, 2001: character 70). 174. Atrium between the inferomediana connecting the cardiac primary filter grooves with the pyloric filter grooves: 0, absent; 1, present (Richter and Scholtz, 2001: character 71). 175. Gut caecae: 0, absent; 1, present along the midgut; 2, restricted to the anterior part of the midgut. Clarke (1979) summarised information on gut caecae in arthropods. The 16 so-called caecae of Campodea (Clarke, 1979), being positioned at the anterior end of the hindgut, are coded here as papillate Malpighian tubules. Codings for Hexapoda follow Bitsch and Bitsch (1998: character 9); Malacostraca is coded for the dorsal caeca following Richter and Scholtz (2001: character 75). 176. Proctodeal dilation: 0, posterior section of hindgut simple, lacking a dilation; 1, proctodeum having a rectal ampulla with differentiated papillae. Bitsch and Bitsch (1998: character 12) homologised the rectal ampulla in Campodea and the rectal ampullae of Insecta. 177. Peritrophic membrane: 0, absent; 1, present. Clarke (1979) documented the distribution of a peritrophic membrane in the gut, noting its presence in onychophorans, myriapods and hexapods [except for Protura (Bitsch and Bitsch, 1998); Snodgrass (1935) noted its presence in Collembola], as well as some crustaceans, including branchiopods (Martin, 1992) and stomatopods (Kunze, 1983). It is absent in tardigrades, chelicerates, pycnogonids and in most isopods (Wägele, 1992). 178. Radiating, tubular diverticula with intracellular final phase of digestion: 0, absent; 1, present. Snodgrass (1952) united euchelicerates (xiphosurids and arachnids) with pycnogonids on the shared presence of their radiating, tubular diverticula, and acknowledged Schlottke’s (1933) observation that the final phase of digestion is intracellular, in the walls of these diverticula. 179. Prosoma and opisthosoma: 0, absent; 1, present. Chelicerate tagmosis is uniquely defined by a prosoma bearing typically six pairs of locomotory and feeding appendages and an opisthosoma composed of, maximally, 12 somites (see Dunlop and Selden, 1998, for discussion of a putative 13 th opisthosomal somite in scorpions). Pycnogonids, however present variation in the presence/absence of the first pair (chelifores) and/or the second pair (palp), and can have up to six pairs of prosomal appendages. 180. Transverse furrows on prosomal carapace corresponding to margins of segmental tergites: 0, absent; 1, present. Shultz (1990) identified segmental furrows on the prosomal carapace as a synapomorphy for the arachnid clade Dromopoda, within which some taxa possess discrete segmental sclerites. Within the Opiliones this is only the case of most members of the Eupnoi and Dyspnoi, but not for Cyphophthalmi or Laniatores, which present a scutum. 181. Fusion of all (opisthosomal) tergites behind the opercular tergite into a thoracetron: 0, absent; 1, present. The fusion of opisthosomal tergites into a thoracetron is regarded as an apomorphy for Xiphosurida (Anderson and Selden, 1997), whereas the Palaeozoic synziphosurine grade have free opisthosomal tergites. 182. Opisthosoma greatly reduced, forming a slender tube emerging from between the posteriormost legs, with a terminal anus: 0, absent; 1, present (Snodgrass, 1952; Wheeler and Hayashi, 1998). 183. Lamellate respiratory organs derived from posterior wall of opisthosomal limb buds: 0, absent; 1, present (Dunlop 1998). 184. Position of lamellate respiratory organs: 0, on opisthosomal segments 3-7; 1, on opisthosomal segments 4-7; 2, on opisthosomal segments 2-3. This character codes for the position of book gills/lungs in Xiphosura (state 0), Scorpiones and Eurypterida (state 1: Braddy et al., 1999) and Tetrapulmonata (state 2). 185. Type of lamellate respiratory organs: 0, book gills; 1, book lungs. Dunlop and Webster (1999) followed Shultz (1990) in dismissing the homology of book lungs in scorpions and tetrapulmonates (other than their more general homology as lamellate respiratory organs of Chelicerata). We code the different segmental position of these organs as character 184. 186. Appendage on first opisthosomal segment: 0, appendage present on first opisthosomal segment in post-embryonic stages; 1, appendage absent. Shultz (1990) cited the lack of an appendage on the first opisthosomal segment as a derived character of arachnids. The appendages of the first abdominal (opisthosomal) segment in Xiphosura, the chilaria, are located between the coxae of leg IV and form a part of the feeding apparatus (character 187). The corresponding segment in Eurypterida bears a plate-like structure, the metastoma, that overlaps the medial portions of the coxae of leg IV. The sternal versus appendicular contributions to the metastoma are uncertain (Dunlop and Webster, 1999). Jeram (1998) considered the metastoma most likely homologous with the sternum of scorpions. Dunlop and Webster (1999) raised the question whether the scorpion sternum could be derived from limb buds observed in the embryo in certain taxa, the subsequent fate of which is ambiguous. Until this homology is determined we have coded an absence of paired appendages in post-embryonic stages in scorpions and arachnids (following Weygoldt and Paulus, 1979). Coding is here restricted to chelicerates (opisthosomal tagmosis is part of the character definition), though outgroup comparison with the homologous segment in other Arachnata (e.g., Trilobita) indicates presence of a limb to be plesiomorphic. 187. Limb VII as chilaria: 0, absent; 1, present. Reduced appendages on somite VII of similar structure are shared by extant Xiphosura. 188. First opisthosomal segment: 0, broad; 1, narrow, developed as pedicel (Shultz, 1990). Shultz (1990: character 40) coded a pedicel as a narrow first opisthosomal segment. Previous workers (Weygoldt and Paulus, 1979) had applied a stricter definition of a pedicel (e.g., restricted to Araneae and Amblypygi), though even this homology has been disputed (Shear et al., 1987). Selden et al. (1991) considered the narrowing of segment 7 to adequately code for an apomorphy of tetrapulmonates. Dunlop (1996: character 7) coded a pedicel as present in Ricinulei, Palpigradi, and Tetrapulmonata, a usage similar to that of Shultz (1990), and the one that we follow here. 189. Abdomen (limb-free somites between the terminal segment and limb-bearing trunk segments, posterior to expression domain of Ubx, abdA and abdB): 0, absent; 1, present. Evidence from Hox genes provides insights into the homologies of tagmata, and forms the basis for not coding hexapod and crustacean "thoraxes" (characters 190, 191) as homologous. Expression of the Hox genes Antp, Ubx, abdA and AbdB has led to the proposal that the crustacean "thorax" or pereion is homologous to the hexapod thorax and abdomen (Averof and Akam, 1995; Deutsch, 1997). Grenier et al. (1997) found this Hox cluster throughout the Panarthropoda, including Onychophora. Crustacea uniquely possess a limbless abdomen, which does not express Ubx and abdA (Grenier et al., 1997, and references therein). The crustaceans coded here all have a morphologically defined limbless abdomen, except for the Malacostraca, Conchostraca and the Remipedia. While a series of limbless abdominal somites have been cited as an autapomorphy for Entomostraca (Walossek, 1999), Scholtz (1995) showed that Engrailed expression extends to additional embryonic segments within the telson of malacostracans. We interpret these somites as the homologue of the abdomen of entomostracans. 190. Pereion tagmosis: 0, one locomotory tagma; 1, two locomotory tagmata. Following from the identity of the crustacean pereion in character 189, the malacostracan pleon is identified as a second set of thoracic segments (Walossek, 1999). Number of somites in the pereion of different crustacean taxa (see Walossek and Muller, 1998, fig. 12.13) is expressed in the coding of variable gonopore positions (character 264) rather than as a separate character (e.g., we have not coded seven pairs of thoracopods and the male gonopore on the seventh pereion segment as independent evidence for a group of the maxillopodans analysed here). Malacostracan trunk tagmosis appears to be distinct in its pattern of Hox gene expression, with well-defined, discrete expression domains of Antp, Ubx and abd-A marking the limits of tagmata (Porcellio: Abzhanov and Kaufman, 2000). 191. Thorax with three limb-bearing segments: 0, absent; 1, present. 192. Meso- and metathorax in mature stages bearing wings: 0, absent; 1, present. 193. Wing flexion: 0, absent; 1, present. A character complex involved in flexion of the wings over the back unites Neoptera (Hennig, 1981), e.g., a pleural muscle on the third axillary sclerite. 194. Segmentation of pleon: 0, seven segments; 1, six segments; 2, five segments (sixth pleomere fused with telson to fom pleotelson). Burnett and Hessler (1973) and Kunze (1983) interpreted the six segmented abdomen in Hoplocarida and Eumalacostraca to be the result of fusion of segments 6 and 7, whereas Leptostraca have a seven-segmented pleon. Coding is restricted to Malacostraca (dependent on presence of pleon; see character 190). The pleotelson of Isopoda is scored following Richter and Scholtz (2001). 195. Diplosegments: 0, absent; 1, present. The fusion of trunk segments into diplosegments is considered a diplopod synapomorphy (Dohle, 1980; Enghoff, 1984; Kraus and Kraus, 1994). 196. Endosternum (ventral tendons fused into prosomal endosternum): 0, absent; 1, present. Euchelicerates are distinct in the modification of the intersegmental tendon system. In the prosoma, the ventral tendons are consolidated into a plate, the endosternum, which is suspended by the dorsoventral muscles (Boudreaux, 1979). Firstman (1973) additionally ascribed a role for fusion of perineural vascular membrane in the endosternum. 197. Dorsal endosternal suspensor of fourth postoral segment with anterolateral carapacal insertion: 0, absent; 1, present (Shultz, 1990). Applicability is limited to Euchelicerata (dependent on presence of the endosternum). 198. Tergal scutes extend laterally into paratergal folds: 0, absent; 1, present. Paratergal folds (paranotae) have been upheld as a basal synapomorphy for Euarthropoda (Boudreaux, 1979; Wägele, 1993). They are lacking or reduced in myriapods, a condition that has been interpreted as an apomorphic reversal (Boudreaux, 1979). Wilson and Shear (2000) indicated the presence of paraterga in diplopods, in contrast to their absence in chilopods, symphylans and pauropods, though they considered diplopod paraterga to be non-homologous with those of non-myriapod arthropods. The "paraterga" of Diplopoda can be accounted for by the medial positioning of the leg bases (character 220). 199. Paramedian sutures: 0, absent; 1, present. Paramedian sutures (Manton, 1965) are a pair of lineations along the tergum and sternum in epimorphic chilopods. Attems (1926) cited this character ("Längsnähten") as a defining character of Epimorpha. 200. Intercalary sclerites: 0, absent; 1, developed as small rings; 2, developed as pretergite and presternite. Well sclerotised intercalary tergites and sternites are present in Craterostigmorpha and Epimorpha (Chilopoda). Weaker sclerotisations occur in the corresponding positions in Lithobiomorpha. A correlated character (dependent on the presence of intercalary sternites) is the anchoring of the tendon of the sternal longitudinal muscles on the intercalary sternite in these chilopods (Manton, 1965). Dohle (1985) indicated that division of the tergites and sternites into preand metatergites and pre- and metasternites was autapomorphic for the Geophilomorpha. Manton (1965) demonstrated that the muscles of the pretergites are more independent of those of the metatergites than are those of other chilopods with intercalary sclerites, and related this to the burrowing habits of the geophilomorphs. The intercalary sclerites of Symphyla (Manton, 1966) are tergal only, and not coded as homologous with those of chilopods. 201. Trunk heterotergy: 0, absent; 1, present (alternating long and short tergites, with reversal of lengths between seventh and eighth walking leg-bearing segments). Borucki (1996) recognised special heterotergy as a synapomorphy of the Chilopoda, with a homologous alternation in long and short tergites between post-maxillipedal segments 7 and 8 in all ingroup taxa except the Geophilomorpha. Heterotergy in non-chilopods (such as Symphyla) does not share this precise segmental homology. 202. Trunk sternites: 0, each segment with large sternum; 1, sternal area divided into two hemisternites by linea ventralis; 2, sternal area mostly membranous, with pair of small sternites; 3, sternal plate (at least that of thoracic segments II and III) bears a Y-shaped ridge/apodeme; 4, sternites extended rearwards to form substernal laminae; 5, thoracic sternal areas reduced and partly invaginated along the median line; 6, sternites lacking. Definition and coding of character states for the sternites in myriapods and hexapods follow Bitsch and Bitsch (2000: character 23) except to add a state for the absence of sternal plates in Pauropoda. State 0 (a large sternite on each segment) is certainly plesiomorphic, its presence recorded in exemplar Crustacea. 203. Trunk endoskeleton in each segment: 0, a pair of lateral connective plates; 1, a pair of sternocoxal rods; 2, complex connective endosternite; 3, endoskeleton mainly cuticular, composed of two intrasegmental furcal arms and an intersegmental spinal process. States for the trunk endoskeleton in myriapods and hexapods are as defined by Bitsch and Bitsch (2000: character 25) except that Japygidae is coded for the presence of the connective endosternite (state 2) as in other Diplura, whereas Bitsch and Bitsch coded an autapomophic state for the additional presence of cuticular spinae. 204. Pleural part of trunk segments: 0, pleurites absent; 1, supracoxal arches (catapleural and anapleural arches) on each segment; 2, pleural part of thoracic segments II and III consisting of a single sclerite with a large pleural process; 3, pleuron in each thoracic segment consists of a single sclerite divided into anterior and posterior parts by pleural suture, from which a pleural apophysis is invaginated, its internal end connected to the furcal arm. State definitions and codings for pleural sclerites in myriapods and hexapods are from Bitsch and Bitsch (2000: character 24), but an absence state is required to code for taxa that lack ‘subcoxal’ pleurites (e.g., Symphyla, Pauropoda). 205. Procoxal and metacoxal pleurites surround coxa: 0, pleurites absent or incompletely surrounding coxa; 1, procoxa and metacoxa surround coxa. The chilopod orders Scolopendromorpha and Geophilomorpha are united by the pronounced development of pleurites around the leg base (Edgecombe et al. 2000, fig. 2D). The metacoxa is a large sclerotisation in the scolopendromorphs (see Manton, 1965, figs. 48, 49), unlike Craterostigmus (Manton, 1965, fig. 74) or the "anamorphic" chilopods. 206. Elongate coxopleurites on anal legs: 0, absent; 1, present. Formation of coxopleurites on the last legbearing segment was cited by Kraus (1998) as a synapomorphy for Craterostigmus and Epimorpha s.s. Defined as such, this character is actually present in all chilopods, which invariably have pleurites fused to the coxa to form a single basal leg sclerite on the anal leg segment. The apomorphy that Kraus (1998) may have been describing as "coxopleurites" is their enhanced differentiation in Craterostigmus and Epimorpha. In scutigeromorphs and lithobiomorphs, the anal leg coxopleurite is shorter than that of Epimorpha, with the former condition more closely resembling the bases of preceding legs. 207. Pleuron filled with small pleurites: 0, absent; 1, present. Geophilomorphs (Edgecombe et al. 2000, fig. 2D) have an elaboration of pleurites (including the so-called scutellum, katopleure, stigmatopleurite and a few more small pleurites) that fill the pleuron except for the pleural furrow (sensu Manton, 1965). 208. Pleurites fused to tergites: 0, pleurites free; 1, pleurites fused. Fused pleurites were considered by Enghoff (1984) to be autapomorphic for Eugnatha. 209. Trochantin: 0, absent; 1, present. Separation of a trochantin at the anterior part of the thoracic coxopleurite is present only in Neoptera (N.P. Kristensen, 1981; 1991; character 43 of Whiting et al., 1997). 210. Longitudinal muscles attach to intersegmental tendons: 0, absent; 1, present. The intersegmental tendon system of euarthropods was reviewed by Boudreaux (1979). Absence of such tendons in pycnogonids is scored following Boudreaux. 211. Lobopods with pads and claws: 0, absent; 1, present. 212. Articulated limbs with intrinsic muscles: 0, absent; 1, present. Nielsen (1995) considered the presence of articulated limbs with intrinsic muscles to be a synapomorphy uniting tardigardes and euarthropods to the exclusion of onychophorans. Monge-Nájera (1995) and Schmidt-Rhaesa et al. (1998) questioned the homology of sclerotised limbs between tardigrades and euarthropods, distinguishing the former as telescopic and the latter as jointed, whereas Dewel and Dewel (1998) noted that telescopic limbs are present only in arthrotardigrades. Brusca and Brusca (1990) stated that tardigrade limb musculature is entirely extrinsic as in onychophorans, but our observations confirm Nielsen’s (1995) coding of intrinsic muscles. 213. Fundamentally biramous post-antennal limbs (endopod and exopod): 0, absent; 1, present. The homology of the limb rami (endopod and exopod) and the basis in stem-group crustaceans and stem-group chelicerates (e.g., trilobites) has been argued by Walossek (1995), and extended to xiphosurans (Walossek and Müller, 1998a, b). The alleged homology of minute thoracic styli in hexapods such as Archaeognatha and the exopod of Crustacea or fossil arachnates (e.g., Delle Cave and Simonetta, 1991; Bitsch, 1994) is unconvincing, and we see no compelling evidence for an exopod in Hexapoda. Rather than coding "strictly uniramous walking legs" in Oniscidea (Brusca, 2000) for the absence of exopods on the thoracopods, we record the presence of the endopod and exopod on the pleopods. 214. Coxopodite with gnathobasic endite lobes medially: 0, absent; 1, present. Gnathobasic feeding was identified as a common feature of crustaceans and chelicerates (Manton, 1964), and was used to support the TCC group (Cisne, 1974). As discussed under character 112, gnathobasic endites/gnathobasic feeding is also shown in the mandible of myriapods and hexapods (Scholtz et al., 1998). Endites have been posited as synapomorphic for Euarthropoda (Wägele and Stanjek, 1995). Coding for chelicerates follows Shultz (1990: character 23) to recognise their absence in many arachnids. 215. Protopod composed of basis and coxa: 0, absent; 1, present. Although variably interpreted, the proximal limb region of Crustacea possesses unique structure, as is most generally distributed on the antenna and mandible. Our coding accommodates Walossek and Müller’s (1990, 1992, 1998a) hypothesis that that the coxa of Crustacea is a novel element, developed from a proximal endite that is more generally shared by Cambrian fossils. The coxa and basis are distinguishable on some limbs (e.g., the maxilliped) in Oniscidea. 216. Paddle-like epipods: 0, absent; 1, present. Hessler (1992) suggested that epipods on cephalocarid, branchiopod and malacostracan (leptostracan) trunk limbs were an apomorphy uniting these crustacean taxa into a clade, Thoracopoda 217. Trunk limbs with lobate endites formed by folds in limb bud: 0, absent; 1, present. Morphogenesis of branchiopod trunk limbs indicates that "phyllopodous" limbs in that group arise from early, radical repatterning compared to Leptostraca, in which the developing limbs preserve fundamentally biramous structure (Williams, 1999). 218. Coxal swing: 0, coxa mobile, promotor-remotor swing between coxa and body; 1, coxa with limited mobility or immobile, promotor-remotor swing between coxa and trochanter. Many arachnids differ from xiphosurids (and other arthropods) in the anchoring of the coxa on the prosoma, with promotorremotor swing shifted distally to the coxa-trochanter joint rather than the coxa-body joint. 219. Coxopodite articulation: 0, arthrodial membrane; 1, pleural condyle; 2, sternal condyle; 3, sternal and pleural condyles; 4, internal plate. Manton (e.g., 1972) attributed much importance to the nature of coxal articulation within the Atelocerata. She contrasted the myriapod condition (coxa with a condylic articulation ventrally, on the sternum) with that observed in most insects (coxa articulating dorsally, on a pleurite). Contrary to Manton’s and Boudreaux’s (1979) characterisations of Myriapoda, Wilson and Shear (2000) indicated that a true coxa-sternite articulation among the Progoneata is confined to Chilognatha, whereas pauropods and polyxenids have the coxae inserted into the body wall, coxalsternal overlap not involving an articulation. In Chilopoda, articulation between the coxa and sternum is pronounced in Craterostigmus (Manton, 1965, fig. 73 accurately illustrates the articulation), whereas geophilomorphs have the procoxal and metacoxal sclerites overlapping the sternite. Unique conditions are recognised for Collembola, in which an internal suspensory system is developed (Manton, 1972), and Protura, which possess sternal as well as pleural condyles. In primitive crustacean taxa as well as early fossil arachnomorphs (Chen et al., 1997), the coxopodite (=basis) joins the body in arthrodial membrane rather than a condylic joint. 220. Separation of coxae of trunk legs: 0, coxae separated laterally; 1, coxae in close approximation mid ventrally. The close medial setting of the trunk coxae serves as a synapomorphy for Diplopoda (cf. Manton, 1956:155). 221. Coxal vesicles: 0, absent; 1, present on numerous trunk segments; 2, restricted to first abdominal segment (modified as Ventraltubus). Dohle (1980) reviewed the distribution of coxal vesicles (or eversible sacs) in Atelocerata. He noted their variable positioning in different progoneate and hexapod taxa, and did not conclude that they provided sound evidence for a monophyletic group. Kraus and Kraus (1994), however, listed coxal vesicles together with styli as a synapomorphy uniting progoneates and hexapods, whereas Moura and Christoffersen (1996) cited a stylus and eversible vesicles as an atelocerate synapomorphy (but did not acknowledge their absence in Chilopoda). Matsuda (1976) distinguished between eversible sacs of appendicular nature (e.g., the single pair of sacs at the end of the Ventraltubus on the first abdominal segment in Collembola and Protura; the vesicles of campodeids, which arise from the appendicular Anlagen of the abdominal segments: Ikeda and Machida, 1998) and those that appear to have extra-appendicular origins. The latter include the vesicles of Symphyla, which arise on the ‘ventral organs’ associated with ganglion formation (Tiegs, 1940), these being segmental thickenings of the embryonic ventral ectoderm. It can thus be validly questioned whether ‘coxal vesicles’ should be regarded as broadly homologous. Although Tiegs (1947) regarded a pair of organs of the collum of pauropods (Edgecombe et al. 2000, fig. 2F) as vesicles, Kraus and Kraus (1994) suggested that they are vestigial appendages. Vesicles are present on numerous trunk segments in symphylans and some groups of diplopods (not the representatives considered here) and on numerous abdominal segments in Diplura, Archaeognatha and Tricholepidion (but absent in Lepismatidae). In contrast, the vesicles are confined to the first abdominal segment in Ellipura (state 2). 222. Styli: 0, absent; 1, present. Styli have a close association with coxal vesicles/eversible sacs in some atelocerate taxa, for example Symphyla and Diplura (see discussion under character 221). However, styli and vesicles do not covary phylogenetically; Ellipura possess vesicles but lack styli. As such we treat these as separate characters (cf. Dohle, 1980) rather than a single feature (Kraus and Kraus, 1994). Evidence for styli in chilopods is contentious, the only evidence being Heymons’ (1901) description of a coxal spur on embryonic appendages of Scolopendra, which has been upheld as being in a position comparable to the coxal stylus of machiloids (Matsuda, 1976). A further distinction could be made between taxa having styli on numerous abdominal/trunk segments in both sexes and those that have more restricted distributions of styli (e.g., on the ninth segment of the adult males only in Ephemeroptera, and some pterygotes, e.g., Blattidae). 223. Musculi laterales: 0, absent; 1, present (Shultz, 1990). The musculi laterales is a term used by spider anatomists to define a lateral tergocoxal muscle from the walking legs that inserts on the pleural membrane adjacent to the coxa rather than on the coxa itself. This type of insertion is found in Araneae and Uropygi (Shultz, 1990). 224. Coxotrochanteral joint: 0, simple; 1, complex (Shultz, 1989, 1990). Coding is restricted to Chelicerata. 225. Trochanteronotal muscle: 0, absent; 1, present. Bitsch and Bitsch (2000: character 31) coded a tergal muscle attached to the trochanter segment in Machilidae, Zygentoma and Pterygota. 226. Trochanter distal joint: 0, mobile; 1, short, ring-like trochanter lacking mobility at joint with prefemur. The very short trochanter in chilopods is part of a proximal region of the leg specialised to facilitate a rapid backstroke (Manton, 1965). Associated with the shortening of the trochanter in chilopods is immobility at its distal joint with the prefemur. Arthropods lacking stenopodial legs, such as most Crustacea, are scored as missing data for this character because the homologues of the myriapodan trochanter and prefemur cannot be identified with reasonable confidence. 227. Trochanterofemoral joint of walking legs: 0, transverse bicondylar; 1, vertical bicondylar (Shultz, 1989, 1990). 228. Unique trochanteral femur-twisting muscle: 0, absent; 1, present. N.P. Kristensen (1998, fig. 21.2E) alleged that a twisting muscle shared by Campodea and Heterojapyx is a possible autapomorphy for Diplura. Coding for this and the following character outside Hexapoda is scored as uncertain. 229. Unique monocondylic femur-tibia pivot joint: 0, absent; 1, present. Manton (1972) described a modified (monocondylic) femur-tibia pivot joint as a diagnostic feature of Diplura, and it has been accepted as a potential dipluran autapomorphy by N.P. Kristensen (1998). 230. Patella/tibia joint: 0, free; 1, fused. Due to uncertain identity of the hexapod patella/tibia joint in other arthropods (and its likely absence in taxa presumed to lack a patella), coding for this character is restricted to hexapods and myriapods. Kristensen (1991) cited "tight union of the patella and the tibia" as a hexapod autapomorphy. This is one expression of the six-segmented leg that is considered a novelty for Hexapoda (Kristensen, 1998; Willmann, 1998). 231. Patellotibial joint of walking legs: 0, dorsal monocondylar; 1, simple bicondylar; 2, vertical bicondylar (Shultz, 1989, 1990). A bicondylar articulation of the patella and tibia defines a subset of Arachnida (Shultz, 1989, 1990). Because homology of the chelicerate patella in other arthropod taxa is uncertain (the patella being widely regarded as lost in extant arthropods other than chelicerates; e.g., Boudreaux, 1979), we have restricted coding of this and other patellar characters (232-233) to the Chelicerata. 232. Femoropatellar joint: 0, transverse dorsal hinge; 1, bicondylar articulation. Shultz (1989, 1990) recognised a bicondylar articulation between the femur and patella as a synapomorphy for the arachnid taxon Dromopoda. 233. Origin of posterior transpatellar muscle: 0, arises on distodorsal surface of femur, traverses femoropatellar joint ventral to axis of rotation, receives fibres from wall of patella; 1, arises on distal process of femur, traverses femoropatellar joint dorsal to axis of rotation, does not receive fibres from patella. The transpatellar muscle corresponds to muscle 7 of Shultz (1989), who noted that its origin in opilionids, scorpions, pseudoscorpions and solifuges was distinctive within Arachnida, although the transpatellar muscles are absent in Cyphophthalmi (Shultz, 1989). Coding is restricted to taxa with a posterior transpatellar muscle (chelicerates). 234. Elastic arthrodial sclerites spanning the tibia-tarsus joints: 0, absent; 1, present (Shultz, 2000). 235. Tarsus segmentation: 0, not subsegmented; 1, subsegmented. Segmentation of the tarsus into tarsomeres has been cited as an apomorphy for several clades within the Arthropoda (e.g., Chilopoda and Arachnida fide Boudreaux, 1979; Insecta fide N.P. Kristensen, 1981, 1991). Shear et al. (1998) conclude that diplopods have a unitary tarsus except for instances of secondary subdivision. We have scored the tarsus as subdivided in Penicillata (Edgecombe et al. 2000, fig. 3B) following the podomere homologies of Manton (1956, text-fig. 3). 236. Tarsal organ: 0, absent; 1, present (Forster, 1980; Selden et al., 1991). Selden et al. (1991) claimed that the tarsal organ of spiders (Blumenthal, 1935) is present in Amblypygi, Uropygi and Schizomida, and disputed homology with similar tarsal structures in scorpions and ticks based on ultrastructural differences. 237. Origin of pretarsal depressor muscle: 0, pretarsal depressor originates on tarsus; 1, pretarsal depressor originates on patella. Shultz (1990) cited a patellar point of origination of the pretarsal depressor as an apomorphic character for arachnids, versus a tarsal origination in pycnogonids and in xiphosurids. Edgecombe et al. (2000: character 158) attempted to code a tibial origination of the pretarsal depressor in myriapods and hexapods within the scope of this character, but we consider the homologies too uncertain to justify that stance. Coding is restricted to Chelicerata. 238. Pretarsal levator muscle: 0, present; 1, absent (depressor is sole pretarsal muscle). Snodgrass (1952) recognised a single pretarsal muscle, a depressor, as a synapomorphy uniting myriapods and hexapods. The crustacean dactylopodite has two muscles, a levator and a depressor (Snodgrass, 1935:198). 239. Pretarsal claws: 0, paired; 1, unpaired. Unpaired pretarsal claws have been upheld as a synapomorphy for Protura and Collembola (N.P. Kristensen, 1981), although it has alternatively been speculated that a single median claw could be the basal state for Hexapoda (Bitsch, 1994) and paired claws a synapomorphy of Insecta (Kraus, 1998). We have scored all myriapods except Symphyla (Edgecombe et al. 2000, fig. 2C) as having unpaired claws based on the condition of the median claw, although accessory claws are commonly paired (chilopods) or a lateral accessory claw may combine with the median claw to simulate pairing (pauropods; Kraus and Kraus, 1994, fig. 7b). Although comparisons have been made with the malacostracan dactylos in an attempt to determine the basal state for this character in atelocerates (e.g., Bitsch, 1994) pretarsal claws are lacking in most Crustacea, and this character is scored as uncertain. Edgecombe et al. (2000) coded claw arrangements in chelicerates in their appraisal of claw pairing in Hexapoda. We consider the codings employed in that work to be erroneous (Limulus) or misleading. Fossil taxa, notably trilobites, eurypterids and basal scorpions, indicate that paired claws in arachnids are derived from telotarsal spurs (Jeram, 1998: character 19). Coding is now restricted to myriapods and hexapods. 240. Pretarsal claw(s) articulation: 0, on pretarsal base; 1, on distal tarsomere. The articulation of the pretarsal claws on the distal tarsomere has been proposed as an insect apomorphy (Boudreaux, 1979; Kristensen, 1998). 241. Plantulae: 0, absent; 1, present. Minet and Bougoin (1986) advocated tarsal plantulae as a synapomorphy for basal neopteran orders (Whiting et al., 1997: character 73). Applicability of this character is scored as dependent upon the presence of a hexapod-type pretarsus (i.e., unguitractor plate to which the plantulae lie distally). 242. Tracheae/spiracles: 0, absent; 1, pleural spiracles; 2, spiracles at bases of walking legs, opening into tracheal pouches; 3, single pair of spiracles on head; 4, dorsal spiracle opening to tracheal lungs; 5, open-ended tracheae with spiracle on second opisthosomal segment; 6, many spiracles scattered on body. Defined as ectodermal tubes with a chitinous intima and respiratory function (Dohle, 1997), tracheae are present in arachnids and onychophorans as well as hexapods and myriapods. Dohle (1997), Kraus (1998) and Hilken (1998) take the diversity in tracheal position and structure in Atelocerata to imply four to seven independent originations of tracheae in that group alone. Given that all of these lineages are sister groups in Kraus’ (1998, fig. 22.3) and Hilken’s (1998, fig. 37) cladograms, the idea that their shared ancestors lacked tracheae is unparsimonious. We have not, however, forced a broad homology of tracheae, acknowledging the weakness of primary homology (Hilken, 1998). Codings of states correspond to Dohle (1997) and Hilken’s (1998) hypotheses of tracheal origins except for coding the pleural tracheae of insects and chilopods as a shared state based on similarities in position, branching, and helical taenidia (Kaufman, 1967). Klass and Kristensen (2001) also support the homology of pleural spiracles in chilopods and hexapods based on similarities in musculature and innervation. Studied representatives of Collembola and Protura lack tracheae and are thus coded for absence; we have not attempted to code for the peculiarities of tracheae in other collembolans and proturans (Xué et al., 1994). Within the Pauropoda, Hexamerocerata share a peculiar spiracle position with diplopods, opening into tracheal pockets that function as apodemes (Kraus and Kraus, 1994). Dohle (1998) emphasised that these similarities provide strong evidence for a common origin. Our coding for Pauropoda is based on an exemplar approach, Pauropodidae lacking tracheae. 243. Longitudinal and transverse connections between segmental tracheal branches: 0, tracheae not connected; 1, tracheae connected. Tracheal commissures and connectives have been recognised as a synapomorphy for Epimorpha within Chilopoda (Manton, 1965) and for Dicondylia within Insecta (N. P. Kristensen, 1981). Hennig (1981) stated that tracheal connectives are more general across Insecta, also being present in Archaeognatha. Our coding follows Bitsch and Bitsch (1998: their character 15), not accepting that the variably developed connections of Archaeognatha are reliably homologous. Coding for characters 244-245 is restricted to chilopods and hexapods with pleural spiracles. 244. Pericardial tracheal system with chiasmata: 0, dendritic tracheae; 1, long, regular pipe-like tracheae with specialised moulting rings. Manton (1965) documented numerous modifications of the tracheal system in Geophilomorpha. These include distinctive pericardial tracheae and a median dorsal atrium, as well as chiasmata between the anastomoses (Hilken, 1997). 245. Abdominal spiracles: 0, present (pleural spiracles on posterior part of trunk); 1, absent on first abdominal segment; 2, absent on all abdominal segments. Štys and Bilinski (1990) stated that the absence of abdominal spiracles is a synapomorphy for campodeids and Ellipura (versus a primitive presence of abdominal spiracles in japygids and insects). To evaluate this character at a more general level it is necessary to homologise the hexapod abdomen with the posterior region of the trunk in myriapods. Evidence from chilopods conforms to Štys and Bilinski’s (1990) polarity (presence of posterior trunk spiracles plesiomorphic). Archaeognatha lack a spiracle on the first abdominal segment alone, and this has been regarded as an autapomorphy (Hennig, 1981; Kristensen, 1991, 1998; Larink, 1997). 246. Abdominal segmentation: 0, six segments; 1, 10 segments; 2, 11 segments; 3, 12 segments. The segmental composition of the hexapod abdomen has received extensive debate (see Matsuda, 1976). We follow Kristensen (1998) and Ikeda and Machida (1998) in defining the insect abdomen as being composed of 11 true segments and a telson (state 2 above). Additional states are employed for Collembola (state 0), Diplura (state 1: Koch, 1997; Ikeda and Machida, 1998) and Protura (state 3). Coding is limited to Hexapoda. 247. Annulated caudal filament: 0, absent; 1, present. Abdominal segment XI (or XII; see character 246) is modified as an annulated caudal filament in Archaeognatha, Zygentoma and primitive pterygotes (Ephemeroptera and Odonata), and has accordingly been cited as an insect synapomorphy (Kukalová-Peck, 1991; Kristensen, 1998; Bitsch and Bitsch, 2000). 248. Abdominal segment XI modified as cerci: 0, absent; 1, present. Cerci are absent in Ellipura (Kristensen, 1991), although Kukalová-Peck (1991:150) referred to their presence in Protura. In addition to segmental homology, cerci in Diplura and Insecta have a modified, fused condylic base; this has been cited as evidence in favour of a dipluran sister group to insects (Kukalová-Peck, 1991). 249. Articulate furcal rami: 0, absent; 1, present. Walossek and Müller (1992) recognised a pair of articulated furcal rami as a shared derived character for the crown-group level of Crustacea. 250. Uropods: 0, absent; 1, present. Uropods are uniquely present in Hoplocarida and Eumalacostraca (McLaughlin, 1982), except for vestigial presence/loss in Brachyura (Scholtz and Richter, 1995). 251. Styliform post-anal telson: 0, absent; 1, present (Bergström et al., 1980). 252. Paired terminal spinnerets: 0, absent; 1, present. Spinnerets serve as a diagnostic character of Symphyla (Kraus and Kraus, 1994; Kraus, 1998). Bitsch and Bitsch (2000: character 35) reviewed evidence for and against an appendicular (versus tergal organ) origin for symphylan spinnerets. 253. Anal segment with a pair of large sense calicles, each with a long sensory seta: 0, absent; 1, present. Flagelliform sensory calicles on the anal segment are unique to Symphyla (Scheller, 1982). 254. Egg cluster guarded until hatching, female coiling around egg cluster: 0, absent; 1, females coils ventrally around cluster; 2, female coils dorsally around egg cluster. A distinctive style of maternal care is shared by Craterostigmomorpha and Epimorpha (Manton, 1965; Dohle, 1985). Dohle (1985) and Borucki (1996) upheld geophilomorph monophyly based on the habit of the female to coil with the dorsum against the eggs, versus the sternum against the eggs in Craterostigmus and Scolopendromorpha. 255. Peripatoid and foetoid stages protected by mother: 0, absent; 1, present. Brood care (character 254) in Epimorpha is extended to the first two postembryonic stadia (Dohle, 1985). 256. Female gonopod used to manipulate single eggs: 0, absent; 1, present. Ax (1999) treated the usage of the female gonopod in egg manipulation and the laying of single eggs as two independent autapomorphies of his taxon Gonopodophora (=Lithobiomorpha + Scutigeromorpha), whereas Edgecombe et al. (1999: character 101) combined this information as a single character. Singlesegmented gonopods are identified in Geophilomorpha, but are lacking in Scolopendromorpha and Craterostigmus (Prunescu, 1996). Absence of gonopods renders the character broadly inapplicable; most instances of gonopods outside Chilopoda (e.g., Diplopoda) cannot be homologised with those of chilopods. 257. Female abdomen with ovipositor formed by gonapophyses of segments VIII and IX: 0, absent; 1, present. The ovipositor is cited as an insect synapomorphy (Kristensen, 1998). Among insects coded here, it is absent in Ephemeroptera and Drosophila (Bitsch and Bitsch, 2000: character 40). 258. Gonangulum sclerite fully developed as ovipositor base, articulating with tergum IX and attached to 1st valvula/valvifer. 0, absent; 1, present. A putative synapomorphy for Dicondylia (N. Kristensen, 1981, 1998), this character is applicable only to those taxa with an ovipositor (character 257). 259. Ovipositor opening at anteroventral part of opisthosoma: 0, absent; 1, present. An ovipositor organ is present in Opiliones and many Acari. 260. Legs of anterior trunk segment(s) transformed into gonopods: 0, absent; 1, present. A copulatory organ (gonopod) on the seventh trunk segment defines Helminthomorpha according to Enghoff (1984). W.A. Shear (pers. comm., 1999) disputes the homology of gonopods, based on their different segmental positions. 261. Dignathan-type penes: 0, absent; 1, present. "Penes" refers to a pair of narrow appendages behind the second trunk leg pair in diplopods and pauropods, bearing the male gonopore at their tips (Dohle, 1980, 1998; Kraus and Kraus, 1994). Edgecombe et al. (2000) coded penes as an unreversed synapomorphy for Dignatha. Presence of penes is restricted to Penicillata and Pauropoda. 262. Penis (spermatopositor) opening on anteroventral part of opisthosoma: 0, absent; 1, present. A penis is present in all the Opiliones, and certain Acari, but absent in the remaining chelicerate taxa. 263. Male parameres: 0, undifferentiated; 1, pair of ‘lateral plates’ on segment XI; 2, pair of parameres on segment IX (second pair variably present on segment VIII); 3, incorporated into the phallic apparatus as sclerites. Homologies and character state definitions for parameres in Hexapoda follow Bitsch and Bitsch (2000: character 42). 264. Penis on abdominal segment IX: 0, absent; 1, present. Bitsch and Bitsch (2000: character 43) coded for homology of median or paired (Ephemeroptera) penes in Ectognatha; we accept this homology based on the segmental correspondence of the copulatory apparatus. A penis is lacking in Collembola and Diplura, whilst that of Protura has a different segmental position (opening between segments XI and XII). 265. Male gonopore location: 0, posterior end (opisthogoneate); 1, somite 11 (sixth pereion segment); 2, somite 12 (seventh pereion segment); 3, somite 8 (first opithosomal segment); 4, behind legs of somite 8 (second pair of trunk legs); 5, somite 13 (eighth pereion segment); 6, somite 17 (twelfth pereion segment); 7, somite 16; 8, on multiple leg bases; 9, between segments VIII and IX, more or less hidden by hind border of sternum VIII; 10, somite 19 (fourteenth trunk segment). This character is coded to recognise varied states of ‘progoneaty’ in chelicerates, crustaceans and myriapods. We acknowledge that state 0 includes additional variation (e.g., male gonopore behind the eleventh abdominal segment in Protura; behind the ninth abdominal segment in insects; coded as character 264). 266. Female gonopore position: 0, on same somite as male; 1, two segments anterior to male; 2, six segments anterior to male; 3, seven segments anterior to male. States 1-3 recognise the separation of male and female gonopores in Malacostraca, Cirripedia, and Remipedia, respectively. Within Hexapoda, Entognatha have male and female gonopores on the same segment (Matsuda, 1976) whereas in Insecta the female gonopore is generally located behind the seventh abdominal sternite, and the male on the ninth segment. 267. Genital operculum divided, incorporated into pedicel: 0, absent; 1, present (Shultz, 1990). 268. Genital operculum overlapping third opisthosomal sternite: 0, absent; 1, present (Shultz, 1990; Dunlop, 1999). 269. Postgenital appendages: 0, opercular and/or lamellar; 1, poorly sclerotized or eversible; 2, absent (Shultz, 1990). Coding is restricted to Chelicerata (i.e., specific to taxa with the gonopore position of chelicerates). 270. Embryonic gonoduct origin: 0, gonoduct arising as a mesodermal coelomoduct; 1, gonoduct arising as a secondary ectodermal ingrowth; 2, gonoduct arising in association with splanchnic mesoderm. The developmental origin of the gonoducts was traced by Anderson (1973). Specialised conditions were described for progoneate myriapods, in which the gonoduct is a secondary ectodermal ingrowth, and hexapods, in which it arises in association with splanchnic mesoderm. Additional apomorphic states can likely be defined within state 0. Tardigrades are scored as unknown; although the gonads have been described as arising from the posterior pair of coelomic pouches, Dewel et al. (1993:171) considered this inadequately established. Glomeris is coded as a proxy for Sphaerotheriida. 271. Genital atrium with looped deferens duct: 0, absent; 1, present. The deferens duct in Chilopoda is looped near its opening (Prunescu, 1996, fig. 5). This character is scored only for those taxa with an unpaired, opisthogoneate deferens duct. 272. Lateral testicular vesicles linked by a central, posteriorly-extended deferens duct: 0, absent; 1, present. Prunescu (1996) described an apomorphic testicular system in Craterostigmus and epimorphic chilopods, in which the vasa efferentia emanate from both ends of the testes. Ordinal groundpatterns have been employed for this and the following character. 273. Testicular follicles with pectinate arrangement: 0, absent (elongated testicular sac or sacs); 1, several pectinate follicles present. State 1 corresponds to a basal apomorphy for Insecta in the analysis of Bitsch and Bitsch (1998: character 24). 274. Spermatophore web produced by ‘Spingriffel’ structure: 0, absent; 1, present. Dohle (1985) indicated that lithobiomorph, scolopendromorph and geophilomorph chilopods spin a web for the deposition of the spermatophore. While the web has been documented in few chilopod taxa, web spinning can be coded based on the ‘Spinngriffel’ structure (so-called penis). Dohle (1990: 77) identified this structure in Craterostigmus. The web material is probably produced by accessory glands of which Pleurostigmophora have two pairs and Scutigeromorpha (which do not produce a web) have a single, rudimentary pair (Brunhuber and Hall, 1970). We code for these shared morphological details associated with web spinning in Chilopoda, rather than the simple presence of a spermatophore web (also observed in Pauropoda, some Diplopoda, Lepismatidae and Tricholepidion: Schaller, 1979; Sturm, 1997). 275. By-passing" foreplay, spermatophore transfer on web, "waiting" ritual by female: 0, absent; 1, present. Rituals associated with indirect sperm transfer in Tricholepidion were described by Sturm (1997), and display detailed similarity with the behavior of lepismatids. Hexapods with copulation/internal fertilisation are scored as inapplicable. 276. Sperm dimorphism: 0, absent; 1, present (microsperm and macrosperm). Although ultrastructural evidence for sperm dimorphism is best known for Scolopendromorpha (Jamieson, 1986), it is consistent with sperm of two sizes in all other chilopod orders except Geophilomorpha (Jamieson, 1987; Carcupino et al., 1999). Such dimorphism is elsewhere known in Symphyla (Dallai and Afzelius, 2000). Sperm dimorphism occurs within Drosophila (Lachaise et al., 1991), but not in the D. melanogaster group. 277. Acrosomal complex in sperm: 0, filamentous actin perforatorium present; 1, monolayered (perforatorium absent); 2, acrosome absent. Codings for the presence of a central rod (performatium) in the sperm are based on Baccetti and Dallai (1978), Baccetti et al. (1979), Jamieson (1987, 1991), Alberti (1995) and Jamieson et al. (1999). Baccetti et al. (1979) particularly regarded the loss of the perforatium to be a shared derived character of Myriapoda. Dallai and Afzelius (2000) described the acrosome of Symphyla as "bilayered", but this involves the presence of an acrosomal vesicle; the perforatorium is not present. Certain groups of Opiliones and Acari have also lost the acrosomal filament, or completely lack an acrosomal complex, even within the same genus, e.g., Siro rubens (with all the acrosomal components) (Juberthie et al., 1976), and Siro duricorius without an acrosomal complex (Alberti, 1995). Nemastoma lugubre presents a peculiar situation, where subacrosomal material spreads between the nuclear envelope and plasmalemma (Alberti, 1995). The presence of the perforatorium in leptophlebiids may be plesiomorphic for Ephemeroptera, but we have not made a groundplan coding. 278. Perforatorium bypasses nucleus: 0, absent (perforatorium penetrates nucleus); 1, present. The sperm of Anaspides resembles that of peracarids (isopods, amphipods, cumaceans) in that the perforatorium bypasses the nucleus, rather than penetrating it as in stomatopods (Jamieson, 1991). The latter condition is inferred to be plesiomorphic [e.g., present in the cephalocarid Hutchinsoniella and remipede Speleonectes (Jamieson, 1991) and in Xiphosura (Alberti, 1995); represented by the endonuclear rod in the collembolan Anurida (Dallai, 1970) and Orthoptera (Jamieson et al., 1999)]. In Aranaeae and in Mastigoproctus, the acrosomal filament (perforatorium) spirals around the periphery of the nucleus (Alberti, 1985). 279. Periacrosomal material: 0, absent; 1, present. Jamieson (1987) cited the presence of periacrosomal material as an insectan apomorphy; this is dependent upon the cytoplasm and microtubules on the acrosomal vesicle in Archaeognatha and Zygentoma being homologous with the periacrosomal material of Pterygota. 280. Striated core in subacrosomal space: 0, absent; 1, present. Baccetti (1979) interpreted the pseudoperforatorium of Juliformia, crystalline subacrosomal material lacking actin, to be a novel organelle. Dallai and Afzelius (2000) described subacrosomal material in Symphyla with an array of tubules (striated core) as possibly homologous with the subacrosomal organelle in diplopods. 281. Centrioles in sperm: 0, proximal and distal centrioles present, not coaxial; 1, coaxial centrioles; 2, single centriole; 3, centrioles absent; 4, doublet centrioles, each with a radial ‘foot’. Wirth (1984) identified the presence of two coaxial centrioles in all flagellate sperm (state 1 above) as an autapomorphy of the Chelicerata, although most Opiliones have aflagellate sperm, and the centrioles have only been described for Siro. The doublet centrioles of malacostracans (state 4: Stomatopoda, Peracarida, Decapoda; Jamieson, 1991) are not regarded as homologous with state 0. Within the tardigrades, members of the family Hypsibiidae present variation in the presence/absence of a midpiece, and thus, characters regarding the midpiece cannot be scored for supraspecific terminals (characters 282-284). 282. Centriole adjunct: 0, absent; 1, present. A wide zone of dense material around the connecting piece of the sperm, the centriole adjunct, was regarded by Jamieson (1987) as an autapomorphy of atelocerates, but lost in Entognatha. However, Dallai et al. (1992) recorded the presence of a centriole adjunct in the proturan Acerentulus. Coding for Lepismatidae interprets the single postcentriolar body (Wingstrand, 1973) as the centriole adjunct (Jamieson, 1987). In those taxa lacking a centriole (character 281, state 3), this character is coded as inapplicable. 283. Sperm ‘accessory bodies’ developed from the centriole: 0, absent; 1, present. Kristensen (1991) followed Jamieson (1987) in regarding 1-3 crystalline ‘accessory bodies’ flanking the axoneme as a synapomorphy of Insecta. 284. Cristate, non-crystalline mitochondrial derivatives in sperm: 0, absent; 1, present. Jamieson (1987) identified two elongate mitochondrial derivatives as a groundplan synapomorphy of Hexapoda, interpreting their absence in Protura as a reversal. 285. Supernumary axonemal tubules (peripheral singlets): 0, absent; 1, present, formed from the manchette; 2, present, formed from axonemal doublets. A 9 + 2 arrangement of axonemal tubules was regarded by Baccetti (1979) as plesiomorphic for arthropods, and this condition is widespread. Insects and campodeids share a 9 + 9 + 2 pattern, a state also found in onychophorans. Dallai and Afzelius (1993) revealed different origins for the hexapod and onychophoran states, which we accordingly code separately. A 9 + 2 axoneme is present in all the studied eutardigrades (e.g., Rebecchi and Bertolani, 1999). 286. Axonemal endpiece ‘plume’: 0, endpiece not extended; 1, endpiece extended, plume-like. Jamieson (1986) postulated that chilopods and pauropods shared derived characters in sperm structure. In particular, he proposed that an expanded endpiece of the axoneme, the so-called ‘plume’, unites these taxa. Data reviewed by Jamieson (1987) confirm the presence of the "plume" in Scolopendromorpha, Geophilomorpha and Lithobiomorpha and Mazzini et al. (1991) indicate that it is present in Scutigeromorpha (Scutigera) as well. 287. Sperm flagellum: 0, present; 1, absent. Štys and Bilinski (1990) proposed that immotile sperm are a synapomorphy for Ellipura/Parainsecta. This condition is also observed within Arachnida, and throughout the Diplopoda, where it has been regarded as a synapomorphy (Enghoff, 1984). Many Crustacea also have aflagellate, immotile sperm, including the Branchiopoda, Cephalocarida, Copepoda, and Malacostraca. The flagellum is absent in Collembola, though the axoneme is present in an encysted state. 288. Nucleus of sperm forms spiral ridge: 0, absent; 1, present. Chilopod sperm possess several modifications, including a spiral ridge on the nucleus (various references cited by Dohle, 1985). A similar corkscrew-appearance of the nucleus is developed within Arachnida (Uropygi and most Araneae: Alberti, 1995), and in eutardigrades (Rebecchi and Bertolani, 1999). Edgecombe et al. (2000: character 202) coded the apomorphy only in Chilopoda. 289. Nucleus of sperm with a manchette of microtubules: 0, absent; 1, present (Alberti and PalaciosVargas, 1987; Shultz, 1990; Alberti, 1995). A manchette of microtubules (or microtubule array, according to Shultz, 1990) is observed in the flagellate spermatozoa of several arachnid orders (Uropygi, Schizomida, Amblypygi, Araneae and Ricinulei). No manchette is found in Scorpiones, Pseudoscorpiones, Palpigradi, Acari, and Xiphosura (Alberti and Palacios-Vargas, 1987). Laniatores is the only group of Opiliones that has a manchette of microtubules (Juberthie and Manier, 1978; Jones and Cokendolpher, 1985). The condition in pycnogonids is polymorphic (van Deurs, 1974a, b), and the character is coded as uncertain. 290. Coiling of spermatozoan flagellum: 0, absent (filiform sperm); 1, present. Coiled-flagellate spermatozoa are found in Pseudoscorpiones, Uropygi, Schizomida, Amblypygi, Araneae and Ricinulei (Alberti, 1995). Coding is necessarily restricted to taxa possessing an axoneme. 291. Medial microtubules in spermatozoan axoneme: 0, two (9+2); 1, three (9 + 3); 2, none (9 + 0) (Baccetti, 1979; Shultz, 1990; Alberti, 1995). This varies between the two xiphosurids: Limulus has 9 + 2, and the Asian horseshoe crabs have 9 + 0 (Alberti, 1995). Araneae, Uropygi, Schizomida and Amblypygi have a 9 + 3 axoneme. However, certain spiders (Lyniphiidae) have a 9 + 0 axoneme, as in Carcinoscorpius, Siro rubens (but not S. duricorius), and many scorpions. Within the variable scorpions, the 9 + 2 pattern is found in Euscorpius, while other genera such as Hadrurus or Centruroides have a 9 + 1 axoneme, and others such as Tityus, Vaejovis, Anuroctonus and Uroctonus have a 9 + 0 pattern (references in Baccetti, 1979). Buthidae has been coded with a 9 + 0 pattern, according to familial groundplan. The condition in pycnogonids is also variable, and does not correspond to any of the described states, so we have coded them as ‘?’. 292. Sperm conjugation: 0, absent; 1, present. Tricholepidion and Lepismatidae share sperm conjugation (Wygodzinsky, 1961; Wingstrand, 1972; Kristensen, 1998). Pairing of the sperm also occurs in several orders of helminthomorph diplopods, including Spirobolida (Baccetti and Dallai, 1978; coded for Narceus). 293. Female spermathecae formed by paired lateral pockets in mouth cavity: 0, absent; 1, present. Kraus (1998) cited the unique oral spermathecae as an autapomorphy of Symphyla. 294. Ovary shape: 0, sac- or tube-shaped, entire; 1, divided into ovarioles; 2, ovarian network. Štys and Bilinski (1990) observed the lack of subdivision of the ovary into ovarioles as a distinctive state in campodeids, proturans and collembolans. Broader comparison (Štys et al., 1993), however, indicates that the lack of metameric ovarioles in these taxa, in contrast to their development in Japygina and Insecta, is certainly a plesiomorphic state. Makioka (1988) regarded the looped ovary of ticks as approximating the basal state for Euchelicerata. Xiphosura (Carcinoscorpius coded from Tachypleus: Makioka and Koike, 1985) and Scorpiones share a complex, network-like ovary (Makioka, 1988, fig. 4). 295. Location of ovary germarium: 0, germarium forms elongate zone in the ventral or lateral ovarian wall; 1, germarium in the terminal part of each egg tube; 2, single, median mound-shaped germarium on the ovarian floor. Bitsch and Bitsch (1998: character 21) contrasted an allegedly myriapod-like position of the gonial tissue in Collembola with its apical position in the ovariole in all other hexapods. State 0 is known for euchelicerates, pycnogonids, onychophorans, chilopods (Lithobius) and some crustaceans (Anostraca); state 1 is present in Notostraca, the conchostracan Limnadia, and Armadillidium (data summarised by Makioka, 1988). An apical germarium is also in Cephalocarida (Hessler and Elofsson, 1996:278), Calanoidea (Boxshall, 1992:369) and Tardigrada (Dewel et al., 1993:171). Progoneate myriapods share a distinctive median, mound-shaped germarium (state 2 above), which is observed in symphylans, pauropods and polyxenid diplopods (Yahata and Makioka, 1994, 1997). We follow Yahata and Makioka’s (1994) interpretation that the germarium is lost in chilognathan diplopods (which instead have paired germ zones on the ovarian wall as the sites of oogonial proliferation and oocyte growth), and code this character as inapplicable for that group. 296. Site for oocyte growth: 0, in ovarian lumen; 1, on outer surface of ovary, in hemocoel, connected by egg stalk. Mandibulate-type (state 0) and chelicerate-type (state 1) oocyte growth patterns follow descriptions by Makioka (1988) and Ikuta and Makioka (1999). The mandibulate pattern was identified in pycnogonids, but a reinterpretation of the pedal space containing the growing oocytes suggests that the oocytes protrude from the ovarian surface into the hemocoel and are stalked as in chelicerates (Miyazaki and Makioka, 1991: coding used for Endeis). The chelicerate pattern is shared by Onychophora according to Makioka (1988), coded as a groundpattern. 297. Coxal organs: 0, absent; 1, present. Rosenberg (1982, 1983a, b) investigated the histology of organs associated with the coxal pores in pleurostigmophoran chilopods. Dohle (1985) and Shear and Bonamo (1988) accepted the homology of these coxal organs and we concur, based on detailed ultrastructural similarity. From an ecological scenario, Prunescu (1996) interpreted the lack of coxal organs as a secondary loss in scutigeromorphs, but this is unparsimonious. 298. Crural glands: 0, absent; 1, present. Monge-Nájera (1995) cited crural glands as a synapomorphy for onychophorans. 299. Pair of repugnatorial glands in the cephalon: 0, absent; 1, present. A pair of odoriferous or repugnatorial glands (opening to ozopores) is found in a more or less lateral position on the carapace of all Opiliones, but not in any other group of chelicerates. 300. Pleural defense glands with benzoquinones: 0, absent; 1, present. Defense glands are unique to Chilognatha (Enghoff, 1984), though they are absent in numerous orders within that group (e.g., Sphaerotheriida). Benzoquinones are the dominant component of the secretions in Juliformia. 301. Coxal gland orifice, leg I: 0, absent; 1, present (Shultz, 1990). Shultz (1990) indicated that the coxal glands of chelicerates may be homologous with paired excretory organs of other arthropods and onychophorans. Homologies with non-chelicerates are left uncertain. 302. Relative position of COI and COII: 0, COI/COII;1, COI/tRNAL(UUR)/COII. Boore et al. (1995, 1998) cited a relocation of tRNAL(UUR) as a shared derived character of hexapods and crustaceans (branchiopods and a eumalacostracan), lacking in diplopods, Lithobius, Limulus, an onychophoran, and a tardigrade. Codings for characters 302-303 are as follow: Peripatopsidae based on Euperipatoides leuckarti (Boore et al. 1998); Eutardigrada based on Thulinia sp. (Boore et al. 1998); Limulus polyphemus (Lavrov et al. 2000); Buthidae based on the Scorpionidae Pandinus imperator (Boore et al. 1998); Lithobius based on Lithobius forficatus (Boore et al. 1998); Narceus based on Narceus sp. (Boore et al. 1998); Locusta migratoria (Flook et al. 1995); Drosophila melanogaster (Lewis et al. 1995); Remipedia based on Speleonectes tulumensis (Boore et al. 1998); Anostraca based on Artemia franciscana (Valverde et al. 1994); Reptantia based on Homarus americanus (Boore et al. 1998). 303. Relative position of tRNAL(CUN): 0, l-rRNA/ tRNAL(CUN)/ND1; 1, l-rRNA/ tRNAL(CUN) / tRNAL(UUR)/ND1 (Boore et al., 1995, 1998).