PLANT HORMONES AND TROPISMS LAB
advertisement

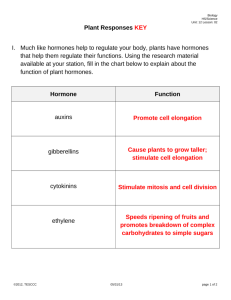
PLANT HORMONES AND TROPISMS LAB INTRODUCTION: Like all living organisms, plants sense and respond to their environment. However, the mechanisms differ substantially from those used by animals such as humans. Rooted to a particular location for most of their lives, plants respond to environmental cues by altering their patterns of growth and development. Most plants have the property of indeterminate growth; that is, they continue to grow and differentiate throughout their lives, and the program for development remains somewhat plastic. This plasticity allows a plant to adjust growth patterns to maximize reproductive success for a given set of environmental conditions. A plant’s morphology and physiology are tuned to their surroundings by an interaction of environmental cues and chemical signals. Among the most important internal signals are small chemical messengers known as plant hormones or growth factors. Similar to animal hormones, plant hormones are produced in small quantities in one part of the plant and are transported to other areas where they exert their effects. However, unlike animals, where hormones are produced in a limited number of places and have very specific target tissues, most plant hormones are produced in a variety of areas in the plant and can exert their effects on a variety of different tissues. In fact, each hormone has a multiplicity of effects, depending upon the site of action, developmental stage of the plant, and concentration of the hormone. Among the important processes affected by plant hormones are cell division, cell enlargement, cell differentiation, polarity of growth, pattern of branching, seed germination, flowering and senescence. Five classes of plant hormones have been identified. These are the auxins, the cytokinins, the gibberellins, abscisic acid, and the gas ethylene. Auxins are defined as any chemical substance that promotes the elongation of coleoptiles. Indole 3-acetic acid (IAA) is the naturally occurring auxin. Although IAA does exert other effects, the major function of IAA is to stimulate elongation of cells in young shoots. IAA is synthesized in the apical meristem of shoots and moves downward to the region of cell elongation where it stimulates cell growth by elongation. This simultaneously suppresses the growth of lateral buds, resulting in the phenomenon of apical dominance. IAA is also involved in the phototropic response of plants, which causes them to grow toward light. Cytokinins are a group of plant hormones which both stimulate cell division and regulate the pathway of differentiation. Cytokinins are chemically related to the nucleotide adenine. They are produced in actively growing tissues, particularly developing embryos, fruits and roots. Cytokinins synthesized in the roots move to their targets in the aerial part of the plant by way of the xylem sap. Cytokinins usually work in combination with auxins and other hormones to regulate growth patterns. For example, the ratio of cytokinin to auxin in a cell determines whether it will differentiate as a shoot or root. The cytokinin/auxin balance is also important for control of apical dominance. While auxin suppresses lateral bud growth, cytokinin stimulates it. Thus, there is more lateral bud growth closer to the roots where they cytokinin levels are higher than the auxin levels. The gibberellins (also known as gibberellic acids or GA) are a large class of naturally occurring plant hormones. They have a variety of effects on plant growth including stem elongation, fruit growth and seed germination. Acting in concert with auxin, the most conspicuous effect of gibberellins is the stimulation of cell division and cell elongation in stems. High doses of gibberellin cause plants to grow very tall and spindly, a phenomenon referred to as bolting. Abnormally low doses cause dwarfism. The fourth class of plant hormones, abscisic acid (ABA), is produced in mature leaves and fruits, and is important in inhibiting the growth of buds and vascular cambium. In so doing, ABA promotes dormancy. ABA (perhaps by stimulating ethylene production) also promotes senescence of leaves. Finally, ABA helps a plant cope with adverse environmental conditions such as drought. During periods of water stress, ABA promotes closure of the stomata, resulting in water conservation for the plant. Unique among the plant hormones, ethylene is a simple gaseous compound that diffuses through the air spaces between plant cells. Ethylene acts as a growth inhibitor. As such, it also is associated with the aging process (senescence), particularly the abscission of leaves, flowers, and fruits. Auxins counteract these aging effects and prolong the length of time that these structures stay on the plant. Thus, the natural balance of auxin and ethylene control the timing of these events in the plant life cycle. As a plant ages, auxin production naturally decreases in the leaves and fruits while ethylene levels increase. The result is the familiar dropping of fruits at the end of the growing season. As stated earlier, plant growth is regulated through interactions of environmental factors and hormonal signals. Many plant hormones act in concert to produce “balanced” growth of the plant. The exact effects of these hormonal signals are contingent upon the environmental cues that the plant receives. The growth responses of plants to environmental cues (or other external stimuli) are called tropisms. Phototropism is a response to a unidirectional source of light and is responsible for the growth of shoots towards light. Cells on the darker side of the plant elongate faster than those on the lighter side, causing the plant to bend toward the light. This is due to an uneven distribution of auxin in the shoot. Illumination of a shoot on one side causes auxin to migrate laterally from the bright side to the dark side. The phototropic response not only affects the morphology of the plant, but more importantly, the ability of the plant to capture the energy of sunlight for photosynthesis. Gravitropism is responsible for the upward growth of the aerial parts and the downward growth of the roots. In other words, shoots exhibit negative gravitropism while roots exhibit positive gravitropism. Gravitropism involves the settling of starch-containing plastids, the statoliths, to the low points of cells. The aggregation of cells at the low points causes the redistribution of calcium, which in turn causes lateral transport of auxins in the roots. Thigmotropism is directional growth in the response to touch and is responsible for the coiling of vines or other climbing plants. A more general form of this response is the stunting of growth in plants that grow in very windy environments. This decrease in stem length is associated with an increase in the thickness of the stem, enabling the plant to withstand strong gusts of wind. This response results from an increased production of ethylene in response to continuous mechanical stimulation. In this laboratory, you will investigate the effects of the hormones on plant growth. You will also examine the effects of light, gravity and touch on plant growth. Using your own experiments and the plants provided to you, distinguish between the four unknown hormones. The hormones you will be working with are listed below: Gibberellic acid (GA3) Indole Acetic Acid (IAA) Dicholorophenoxyacetic acid (2,4 – D) – A synthetic auxin N-dimethylaminosuccinamic acid (B-9) – A synthetic growth retardant Design an experiment to investigate the effect of external stimuli (light, gravity and/or touch) on plant growth.