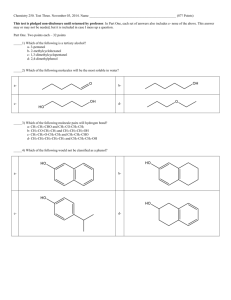
lecture notes ch 1 - Houston Community College Learning Web
advertisement

Chapter 1 Exploring Life Lecture Outline Overview Biology is the scientific study of life. You are starting your study of biology during its most exciting era. The largest and best-equipped community of scientists in history is beginning to solve problems that once seemed unsolvable. Biology is an ongoing inquiry about the nature of life. Biologists are moving closer to understanding: How a single cell develops into an adult animal or plant. How plants convert solar energy into the chemical energy of food. How the human mind works. How living things interact in biological communities. How the diversity of life evolved from the first microbes. Research breakthroughs in genetics and cell biology are transforming medicine and agriculture. Neuroscience and evolutionary biology are reshaping psychology and sociology. Molecular biology is providing new tools for anthropology and criminology. New models in ecology are helping society to evaluate environmental issues, such as the causes and biological consequences of global warming. Unifying themes pervade all of biology. A. Exploring Life on Its Many Levels Life’s basic characteristic is a high degree of order. Each level of biological organization has emergent properties. Biological organization is based on a hierarchy of structural levels, each building on the levels below. At the lowest level are atoms that are ordered into complex biological molecules. Biological molecules are organized into structures called organelles, the components of cells. Cells are the fundamental unit of structure and function of living things. Some organisms consist of a single cell; others are multicellular aggregates of specialized cells. Whether multicellular or unicellular, all organisms must accomplish the same functions: uptake and processing of nutrients, excretion of wastes, response to environmental stimuli, and reproduction. Multicellular organisms exhibit three major structural levels above the cell: similar cells are grouped into tissues, several tissues coordinate to form organs, and several organs form an organ system. Reference: Guide for Biology, Campbell, Reece and Mitchell 1 IG Lecture Outlines 1- For example, to coordinate locomotory movements, sensory information travels from sense organs to the brain, where nervous tissues composed of billions of interconnected neurons—supported by connective tissue—coordinate signals that travel via other neurons to the individual muscle cells. Organisms belong to populations, localized groups of organisms belonging to the same species. Populations of several species in the same area comprise a biological community. Populations interact with their physical environment to form an ecosystem. The biosphere consists of all the environments on Earth that are inhabited by life. 1. Organisms interact continuously with their environment. Each organism interacts with its environment, which includes other organisms as well as nonliving factors. Both organism and environment are affected by the interactions between them. The dynamics of any ecosystem include two major processes: the cycling of nutrients and the flow of energy from sunlight to producers to consumers. In most ecosystems, producers are plants and other photosynthetic organisms that convert light energy to chemical energy. Consumers are organisms that feed on producers and other consumers. All the activities of life require organisms to perform work, and work requires a source of energy. The exchange of energy between an organism and its environment often involves the transformation of energy from one form to another. In all energy transformations, some energy is lost to the surroundings as heat. In contrast to chemical nutrients, which recycle within an ecosystem, energy flows through an ecosystem, usually entering as light and exiting as heat. 2. Cells are an organism’s basic unit of structure and function. The cell is the lowest level of structure that is capable of performing all the activities of life. For example, the ability of cells to divide is the basis of all reproduction and the basis of growth and repair of multicellular organisms. Understanding how cells work is a major research focus of modern biology. At some point, all cells contain deoxyribonucleic acid, or DNA, the heritable material that directs the cell’s activities. DNA is the substance of genes, the units of inheritance that transmit information from parents to offspring. Each of us began life as a single cell stocked with DNA inherited from our parents. DNA in human cells is organized into chromosomes. Each chromosome has one very long DNA molecule, with hundreds or thousands of genes arranged along its length. The DNA of chromosomes replicates as a cell prepares to divide. Each of the two cellular offspring inherits a complete set of genes. In each cell, the genes along the length of DNA molecules encode the information for building the cell’s other molecules. DNA thus directs the development and maintenance of the entire organism. Most genes program the cell’s production of proteins. Each DNA molecule is made up of two long chains arranged in a double helix. Reference: Guide for Biology, Campbell, Reece and Mitchell 2 IG Lecture Outlines 1- Each link of a chain is one of four nucleotides, encoding the cell’s information in chemical letters. The sequence of nucleotides along each gene codes for a specific protein with a unique shape and function. Almost all cellular activities involve the action of one or more proteins. DNA provides the heritable blueprints, but proteins are the tools that actually build and maintain the cell. All forms of life employ essentially the same genetic code. Because the genetic code is universal, it is possible to engineer cells to produce proteins normally found only in some other organism. The library of genetic instructions that an organism inherits is called its genome. The chromosomes of each human cell contain about 3 billion nucleotides, including genes coding for more than 70,000 kinds of proteins, each with a specific function. Every cell is enclosed by a membrane that regulates the passage of material between a cell and its surroundings. Every cell uses DNA as its genetic material. There are two basic types of cells: prokaryotic cells and eukaryotic cells. The cells of the microorganisms called bacteria and archaea are prokaryotic. All other forms of life have more complex eukaryotic cells. Eukaryotic cells are subdivided by internal membranes into various organelles. In most eukaryotic cells, the largest organelle is the nucleus, which contains the cell’s DNA as chromosomes. The other organelles are located in the cytoplasm, the entire region between the nucleus and outer membrane of the cell. Prokaryotic cells are much simpler and smaller than eukaryotic cells. In a prokaryotic cell, DNA is not separated from the cytoplasm in a nucleus. There are no membrane-enclosed organelles in the cytoplasm. All cells, regardless of size, shape, or structural complexity, are highly ordered structures that carry out complicated processes necessary for life. 3. Biological systems are much more than the sum of their parts. “The whole is greater than the sum of its parts.” The combination of components can form a more complex organization called a system. Examples of biological systems are cells, organisms, and ecosystems. Consider the levels of life. With each step upward in the hierarchy of biological order, novel properties emerge that are not present at lower levels. These emergent properties result from the arrangements and interactions between components as complexity increases. A cell is much more than a bag of molecules. Our thoughts and memories are emergent properties of a complex network of neurons. This theme of emergent properties accents the importance of structural arrangement. The emergent properties of life are not supernatural or unique to life but simply reflect a hierarchy of structural organization. Reference: Guide for Biology, Campbell, Reece and Mitchell 3 IG Lecture Outlines 1- The emergent properties of life are particularly challenging because of the unparalleled complexity of living systems. The complex organization of life presents a dilemma to scientists seeking to understand biological processes. We cannot fully explain a higher level of organization by breaking it down into its component parts. At the same time, it is futile to try to analyze something as complex as an organism or cell without taking it apart. Reductionism, reducing complex systems to simpler components, is a powerful strategy in biology. The Human Genome Project—the sequencing of the genome of humans and many other species—is heralded as one of the greatest scientific achievements ever. Research is now moving on to investigate the function of genes and the coordination of the activity of gene products. Biologists are beginning to complement reductionism with new strategies for understanding the emergent properties of life—how all of the parts of biological systems are functionally integrated. The ultimate goal of systems biology is to model the dynamic behavior of whole biological systems. Accurate models allow biologists to predict how a change in one or more variables will impact other components and the whole system. Scientists investigating ecosystems pioneered this approach in the 1960s with elaborate models diagramming the interactions of species and nonliving components in ecosystems. Systems biology is now becoming increasingly important in cellular and molecular biology, driven in part by the deluge of data from the sequencing of genomes and our increased understanding of protein functions. In 2003, a large research team published a network of protein interactions within a cell of a fruit fly. Three key research developments have led to the increased importance of systems biology. 1. High-throughput technology. Systems biology depends on methods that can analyze biological materials very quickly and produce enormous amounts of data. An example is the automatic DNA-sequencing machines used by the Human Genome Project. 2. Bioinformatics. The huge databases from high-throughput methods require computing power, software, and mathematical models to process and integrate information. 3. Interdisciplinary research teams. Systems biology teams may include engineers, medical scientists, physicists, chemists, mathematicians, and computer scientists as well as biologists. 4. Regulatory mechanisms ensure a dynamic balance in living systems. Chemical processes within cells are accelerated, or catalyzed, by specialized protein molecules, called enzymes. Each type of enzyme catalyzes a specific chemical reaction. In many cases, reactions are linked into chemical pathways, each step with its own enzyme. How does a cell coordinate its various chemical pathways? Many biological processes are self-regulating: the output or product of a process regulates that very process. In negative feedback, or feedback inhibition, accumulation of an end product of a process slows or stops that process. Reference: Guide for Biology, Campbell, Reece and Mitchell 4 IG Lecture Outlines 1- Though less common, some biological processes are regulated by positive feedback, in which an end product speeds up its own production. Feedback is common to life at all levels, from the molecular level to the biosphere. Such regulation is an example of the integration that makes living systems much greater than the sum of their parts. B. Evolution, Unity, and Diversity Biology can be viewed as having two dimensions: a “vertical” dimension covering the size scale from atoms to the biosphere and a “horizontal” dimension that stretches across the diversity of life. The latter includes not only present-day organisms, but also those that have existed throughout life’s history. 1. The unity and diversity of life. Life is enormously diverse. Biologists have identified and named about 1.8 million species. This diversity includes 5,200 known species of prokaryotes, 100,000 fungi, 290,000 plants, 50,000 vertebrates, and 1,000,000 insects. Thousands of newly identified species are added each year. Estimates of the total species count range from 10 million to more than 200 million. In the face of this complexity, humans are inclined to categorize diverse items into a smaller number of groups. Taxonomy is the branch of biology that names and classifies species into a hierarchical order. Until the past decade, biologists divided the diversity of life into five kingdoms. New methods, including comparisons of DNA among organisms, have led to a reassessment of the number and boundaries of the kingdoms. Various classification schemes now include six, eight, or even dozens of kingdoms. Coming from this debate has been the recognition that there are three even higher levels of classifications, the domains. The three domains are Bacteria, Archaea, and Eukarya. The first two domains, domain Bacteria and domain Archaea, consist of prokaryotes. All the eukaryotes are now grouped into various kingdoms of the domain Eukarya. The recent taxonomic trend has been to split the single-celled eukaryotes and their close relatives into several kingdoms. Domain Eukarya also includes the three kingdoms of multicellular eukaryotes: the kingdoms Plantae, Fungi, and Animalia. These kingdoms are distinguished partly by their modes of nutrition. Most plants produce their own sugars and food by photosynthesis. Most fungi are decomposers that absorb nutrients by breaking down dead organisms and organic wastes. Animals obtain food by ingesting other organisms. Underlying the diversity of life is a striking unity, especially at the lower levels of organization. The universal genetic language of DNA unites prokaryotes and eukaryotes. Among eukaryotes, unity is evident in many details of cell structure. Above the cellular level, organisms are variously adapted to their ways of life. Reference: Guide for Biology, Campbell, Reece and Mitchell 5 IG Lecture Outlines 1- How do we account for life’s dual nature of unity and diversity? The process of evolution explains both the similarities and differences among living things. 2. Evolution is the core theme of biology. The history of life is a saga of a changing Earth billions of years old, inhabited by a changing cast of living forms. Charles Darwin brought evolution into focus in 1859 when he presented two main concepts in one of the most important and controversial books ever written, On the Origin of Species by Natural Selection. Darwin’s first point was that contemporary species arose from a succession of ancestors through “descent with modification.” This term captured the duality of life’s unity and diversity: unity in the kinship among species that descended from common ancestors and diversity in the modifications that evolved as species branched from their common ancestors. Darwin’s second point was his mechanism for descent with modification: natural selection. Darwin inferred natural selection by connecting two observations: Observation 1: Individual variation. Individuals in a population of any species vary in many heritable traits. Observation 2: Overpopulation and competition. Any population can potentially produce far more offspring than the environment can support. This creates a struggle for existence among variant members of a population. Inference: Unequal reproductive success. Darwin inferred that those individuals with traits best suited to the local environment would leave more healthy, fertile offspring. Inference: Evolutionary adaptation. Unequal reproductive success can lead to adaptation of a population to its environment. Over generations, heritable traits that enhance survival and reproductive success will tend to increase in frequency among a population’s individuals. The population evolves. Natural selection, by its cumulative effects over vast spans of time, can produce new species from ancestral species. For example, a population fragmented into several isolated populations in different environments may gradually diversify into many species as each population adapts over many generations to different environmental problems. Fourteen species of finches found on the Galápagos Islands diversified after an ancestral finch species reached the archipelago from the South American mainland. Each species is adapted to exploit different food sources on different islands. Biologists’ diagrams of evolutionary relationships generally take a treelike form. Just as individuals have a family tree, each species is one twig of a branching tree of life. Similar species like the Galápagos finches share a recent common ancestor. Finches share a more distant ancestor with all other birds. The common ancestor of all vertebrates is even more ancient. Trace life back far enough, and there is a shared ancestor of all living things. All of life is connected through its long evolutionary history. C. The Process of Science 1. Biologists use various forms of inquiry to explore life. Reference: Guide for Biology, Campbell, Reece and Mitchell 6 IG Lecture Outlines 1- The word science is derived from a Latin verb meaning “to know.” At the heart of science is inquiry, people asking questions about nature and focusing on specific questions that can be answered. The process of science blends two types of exploration: discovery science and hypothesis-based science. Discovery science is mostly about discovering nature. Hypothesis-based science is mostly about explaining nature. Most scientific inquiry combines the two approaches. Discovery science describes natural structures and processes as accurately as possible through careful observation and analysis of data. Discovery science built our understanding of cell structure and is expanding our databases of genomes of diverse species. Observation is the use of the senses to gather information, which is recorded as data. Data can be qualitative or quantitative. Quantitative data are numerical measurements. Qualitative data may be in the form of recorded descriptions. Jane Goodall has spent decades recording her observations of chimpanzee behavior during field research in Gambia. She has also collected volumes of quantitative data over that time. Discovery science can lead to important conclusions based on inductive reasoning. Through induction, we derive generalizations based on a large number of specific observations. In science, inquiry frequently involves the proposing and testing of hypotheses. A hypothesis is a tentative answer to a well-framed question. It is usually an educated postulate, based on past experience and the available data of discovery science. A scientific hypothesis makes predictions that can be tested by recording additional observations or by designing experiments. A type of logic called deduction is built into hypothesis-based science. In deductive reasoning, reasoning flows from the general to the specific. From general premises, we extrapolate to a specific result that we should expect if the premises are true. In hypothesis-based science, deduction usually takes the form of predictions about what we should expect if a particular hypothesis is correct. We test the hypothesis by performing the experiment to see whether or not the results are as predicted. Deductive logic takes the form of “If . . . then” logic. Scientific hypotheses must be testable. There must be some way to check the validity of the idea. Scientific hypotheses must be falsifiable. There must be some observation or experiment that could reveal if a hypothesis is actually not true. Reference: Guide for Biology, Campbell, Reece and Mitchell 7 IG Lecture Outlines 1- The ideal in hypothesis-based science is to frame two or more alternative hypotheses and design experiments to falsify them. No amount of experimental testing can prove a hypothesis. A hypothesis gains support by surviving various tests that could falsify it, while testing falsifies alternative hypotheses. Facts, in the form of verifiable observations and repeatable experimental results, are the prerequisites of science. 2. We can explore the scientific method. There is an idealized process of inquiry called the scientific method. Very few scientific inquiries adhere rigidly to the sequence of steps prescribed by the textbook scientific method. Discovery science has contributed a great deal to our understanding of nature without most of the steps of the so-called scientific method. We will consider a case study of scientific research. This case begins with a set of observations and generalizations from discovery science. Many poisonous animals have warning coloration that signals danger to potential predators. Imposter species mimic poisonous species, although they are harmless. An example is the bee fly, a nonstinging insect that mimics a honeybee. What is the function of such mimicry? What advantage does it give the mimic? In 1862, Henry Bates proposed that mimics benefit when predators mistake them for harmful species. This deception may lower the mimic’s risk of predation. In 2001, David and Karin Pfennig and William Harcombe of the University of North Carolina designed a set of field experiments to test Bates’s mimicry hypothesis. In North and South Carolina, a poisonous snake called the eastern coral snake has warning red, yellow, and black coloration. Predators avoid these snakes. It is unlikely that predators learn to avoid coral snakes, as a strike is usually lethal. Natural selection may have favored an instinctive recognition and avoidance of the warning coloration of the coral snake. The nonpoisonous scarlet king snake mimics the ringed coloration of the coral snake. Both king snakes and coral snake live in the Carolinas, but the king snake’s range also extends into areas without coral snakes. The distribution of these two species allowed the Pfennigs and Harcombe to test a key prediction of the mimicry hypothesis. Mimicry should protect the king snake from predators, but only in regions where coral snakes live. Predators in non–coral snake areas should attack king snakes more frequently than predators that live in areas where coral snakes are present. To test the mimicry hypothesis, Harcombe made hundreds of artificial snakes. The experimental group had the red, black, and yellow ring pattern of king snakes. The control group had plain, brown coloring. Equal numbers of both types were placed at field sites, including areas where coral snakes are absent. Reference: Guide for Biology, Campbell, Reece and Mitchell 8 IG Lecture Outlines 1- After four weeks, the scientists retrieved the fake snakes and counted bite or claw marks. Foxes, coyotes, raccoons, and black bears attacked snake models. The data fit the predictions of the mimicry hypothesis. The ringed snakes were attacked by predators less frequently than the brown snakes only within the geographic range of the coral snakes. The snake mimicry experiment provides an example of how scientists design experiments to test the effect of one variable by canceling out the effects of unwanted variables. The design is called a controlled experiment. An experimental group (artificial king snakes) is compared with a control group (artificial brown snakes). The experimental and control groups differ only in the one factor the experiment is designed to test—the effect of the snake’s coloration on the behavior of predators. The brown artificial snakes allowed the scientists to rule out such variables as predator density and temperature as possible determinants of number of predator attacks. Scientists do not control the experimental environment by keeping all variables constant. Researchers usually “control” unwanted variables, not by eliminating them but by canceling their effects using control groups. 3. Let’s look at the nature of science. There are limitations to the kinds of questions that science can address. These limits are set by science’s requirements that hypotheses are testable and falsifiable and that observations and experimental results be repeatable. The limitations of science are set by its naturalism. Science seeks natural causes for natural phenomena. Science cannot support or falsify supernatural explanations, which are outside the bounds of science. Everyday use of the term theory implies an untested speculation. The term theory has a very different meaning in science. A scientific theory is much broader in scope than a hypothesis. This is a hypothesis: “Mimicking poisonous snakes is an adaptation that protects nonpoisonous snakes from predators.” This is a theory: “Evolutionary adaptations evolve by natural selection.” A theory is general enough to generate many new, specific hypotheses that can be tested. Compared to any one hypothesis, a theory is generally supported by a much more massive body of evidence. The theories that become widely adopted in science (such as the theory of adaptation by natural selection) explain many observations and are supported by a great deal of evidence. In spite of the body of evidence supporting a widely accepted theory, scientists may have to modify or reject theories when new evidence is found. As an example, the five-kingdom theory of biological diversity eroded as new molecular methods made it possible to test some of the hypotheses about the relationships between living organisms. Scientists may construct models in the form of diagrams, graphs, computer programs, or mathematical equations. Models may range from lifelike representations to symbolic schematics. Reference: Guide for Biology, Campbell, Reece and Mitchell 9 IG Lecture Outlines 1- Science is an intensely social activity. Most scientists work in teams, which often include graduate and undergraduate students. Both cooperation and competition characterize scientific culture. Scientists attempt to confirm each other’s observations and may repeat experiments. They share information through publications, seminars, meetings, and personal communication. Scientists may be very competitive when converging on the same research question. Science as a whole is embedded in the culture of its times. For example, recent increases in the proportion of women in biology have had an impact on the research being performed. For instance, there has been a switch in focus in studies of the mating behavior of animals from competition among males for access to females to the role that females play in choosing mates. Recent research has revealed that females prefer bright coloration that “advertises” a male’s vigorous health, a behavior that enhances a female’s probability of having healthy offspring. Some philosophers of science argue that scientists are so influenced by cultural and political values that science is no more objective than other ways of “knowing nature.” At the other extreme are those who view scientific theories as though they were natural laws. The reality of science is somewhere in between. The cultural milieu affects scientific fashion, but need for repeatability in observation and hypothesis testing distinguishes science from other fields. If there is “truth” in science, it is based on a preponderance of the available evidence. 4. Science and technology are functions of society. Although science and technology may employ similar inquiry patterns, their basic goals differ. The goal of science is to understand natural phenomena. Technology applies scientific knowledge for some specific purpose. Technology results from scientific discoveries applied to the development of goods and services. Scientists put new technology to work in their research. Science and technology are interdependent. The discovery of the structure of DNA by Watson and Crick sparked an explosion of scientific activity. These discoveries made it possible to manipulate DNA, enabling genetic technologists to transplant foreign genes into microorganisms and mass-produce valuable products. DNA technology and biotechnology have revolutionized the pharmaceutical industry. They have had an important impact on agriculture and the legal profession. The direction that technology takes depends less on science than it does on the needs of humans and the values of society. Debates about technology center more on “should we do it” than “can we do it.” With advances in technology come difficult choices, informed as much by politics, economics, and cultural values as by science. Scientists should educate politicians, bureaucrats, corporate leaders, and voters about how science works and about the potential benefits and hazards of specific technologies. D. Review: Using Themes to Connect the Concepts of Biology Reference: Guide for Biology, Campbell, Reece and Mitchell 10 IG Lecture Outlines 1- In some ways, biology is the most demanding of all sciences, partly because living systems are so complex and partly because biology is a multidisciplinary science that requires knowledge of chemistry, physics, and mathematics. Biology is also the science most connected to the humanities and social sciences. Reference: Guide for Biology, Campbell, Reece and Mitchell Reference: Guide for Biology, Campbell, Reece and Mitchell 11 IG Lecture Outlines 1- Chapter 2 The Chemical Context of Life Lecture Outline Overview Living organisms and the world they live in are subject to the basic laws of physics and chemistry. Biology is a multidisciplinary science, drawing on insights from other sciences. Life can be organized into a hierarchy of structural levels. At each successive level, additional emergent properties appear. A. Elements and Compounds 1. Matter consists of chemical elements in pure form and in combinations called compounds. Organisms are composed of matter. Matter is anything that takes up space and has mass. Matter is made up of elements. An element is a substance that cannot be broken down into other substances by chemical reactions. There are 92 naturally occurring elements. Each element has a unique symbol, usually the first one or two letters of the name. Some of the symbols are derived from Latin or German names. A compound is a substance consisting of two or more elements in a fixed ratio. Table salt (sodium chloride or NaCl) is a compound with equal numbers of atoms of the elements chlorine and sodium. While pure sodium is a metal and chlorine is a gas, they combine to form an edible compound. This change in characteristics when elements combine to form a compound is an example of an emergent property. 2. 25 chemical elements are essential to life. About 25 of the 92 natural elements are known to be essential for life. Four elements—carbon (C), oxygen (O), hydrogen (H), and nitrogen (N)—make up 96% of living matter. Most of the remaining 4% of an organism’s weight consists of phosphorus (P), sulfur (S), calcium (Ca), and potassium (K). Trace elements are required by an organism but only in minute quantities. Some trace elements, like iron (Fe), are required by all organisms. Other trace elements are required by only some species. For example, a daily intake of 0.15 milligrams of iodine is required for normal activity of the human thyroid gland. Reference: Guide for Biology, Campbell, Reece and Mitchell 12 IG Lecture Outlines 1- B. Atoms and Molecules 1. Atomic structure determines the behavior of an element. Each element consists of unique atoms. An atom is the smallest unit of matter that still retains the properties of an element. Atoms are composed of even smaller parts, called subatomic particles. Two of these, neutrons and protons, are packed together to form a dense core, the atomic nucleus, at the center of an atom. Electrons can be visualized as forming a cloud of negative charge around the nucleus. Each electron has one unit of negative charge. Each proton has one unit of positive charge. Neutrons are electrically neutral. The attractions between the positive charges in the nucleus and the negative charges of the electrons keep the electrons in the vicinity of the nucleus. A neutron and a proton are almost identical in mass, about 1.7 × 10−24 gram per particle. For convenience, a smaller unit of measure, the dalton, is used to measure the mass of subatomic particles, atoms, or molecules. The mass of a neutron or a proton is close to 1 dalton. The mass of an electron is about 1/2000 that of a neutron or proton. Therefore, we typically ignore the contribution of electrons when determining the total mass of an atom. All atoms of a particular element have the same number of protons in their nuclei. This number of protons is the element’s unique atomic number. The atomic number is written as a subscript before the symbol for the element. For example, 2He means that an atom of helium has 2 protons in its nucleus. Unless otherwise indicated, atoms have equal numbers of protons and electrons and, therefore, no net charge. Therefore, the atomic number tells us the number of protons and the number of electrons that are found in a neutral atom of a specific element. The mass number is the sum of the number of protons and neutrons in the nucleus of an atom. Therefore, we can determine the number of neutrons in an atom by subtracting the number of protons (the atomic number) from the mass number. The mass number is written as a superscript before an element’s symbol (for example, 4He). The atomic weight of an atom, a measure of its mass, can be approximated by the mass number. For example, 4He has a mass number of 4 and an estimated atomic weight of 4 daltons. More precisely, its atomic weight is 4.003 daltons. While all atoms of a given element have the same number of protons, they may differ in the number of neutrons. Two atoms of the same element that differ in the number of neutrons are called isotopes. In nature, an element occurs as a mixture of isotopes. For example, 99% of carbon atoms have 6 neutrons (12C). Most of the remaining 1% of carbon atoms have 7 neutrons (13C) while the rarest carbon isotope, with 8 neutrons, is 14C. Reference: Guide for Biology, Campbell, Reece and Mitchell 13 IG Lecture Outlines 1- Most isotopes are stable; they do not tend to lose particles. Both 12C and 13C are stable isotopes. The nuclei of some isotopes are unstable and decay spontaneously, emitting particles and energy. 14C is one of these unstable isotopes, or radioactive isotopes. When 14C decays, one of its neutrons is converted to a proton and an electron. This converts 14C to 14N, transforming the atom to a different element. Radioactive isotopes have many applications in biological research. Radioactive decay rates can be used to date fossils. Radioactive isotopes can be used to trace atoms through metabolic processes. Radioactive isotopes are also used to diagnose medical disorders. For example, a known quantity of a substance labeled with a radioactive isotope can be injected into the blood, and its rate of excretion in the urine can be measured. Also, radioactive tracers can be used with imaging instruments to monitor chemical processes in the body. While useful in research and medicine, the energy emitted in radioactive decay is hazardous to life. This energy can destroy molecules within living cells. The severity of damage depends on the type and amount of radiation that the organism absorbs. 2. Electron configuration influences the chemical behavior of an atom. Simplified models of the atom greatly distort the atom’s relative dimensions. To gain an accurate perspective of the relative proportions of an atom, if the nucleus was the size of a golf ball, the electrons would be moving about 1 kilometer from the nucleus. Atoms are mostly empty space. When two elements interact during a chemical reaction, it is actually their electrons that are involved. The nuclei do not come close enough to interact. The electrons of an atom vary in the amount of energy they possess. Energy is the ability to do work. Potential energy is the energy that matter stores because of its position or location. Water stored behind a dam has potential energy that can be used to do work turning electric generators. Because potential energy has been expended, the water stores less energy at the bottom of the dam than it did in the reservoir. Electrons have potential energy because of their position relative to the nucleus. The negatively charged electrons are attracted to the positively charged nucleus. The farther electrons are from the nucleus, the more potential energy they have. Changes in an electron’s potential energy can only occur in steps of a fixed amount, moving the electron to a fixed location relative to the nucleus. An electron cannot exist between these fixed locations. The different states of potential energy that the electrons of an atom can have are called energy levels or electron shells. The first shell, closest to the nucleus, has the lowest potential energy. Electrons in outer shells have more potential energy. Reference: Guide for Biology, Campbell, Reece and Mitchell 14 IG Lecture Outlines 1- Electrons can change their position only if they absorb or release a quantity of energy that matches the difference in potential energy between the two levels. The chemical behavior of an atom is determined by its electron configuration—the distribution of electrons in its electron shells. The first 18 elements, including those most important in biological processes, can be arranged in 8 columns and 3 rows. Elements in the same row fill the same shells with electrons. Moving from left to right, each element adds one electron (and proton) from the element before. The first electron shell can hold only 2 electrons. The two electrons of helium fill the first shell. Atoms with more than two electrons must place the extra electrons in higher shells. For example, lithium, with three electrons, has two in the first shell and one in the second shell. The second shell can hold up to 8 electrons. Neon, with 10 total electrons, has two in the first shell and eight in the second, filling both shells. The chemical behavior of an atom depends mostly on the number of electrons in its outermost shell, the valence shell. Electrons in the valence shell are known as valence electrons. Lithium has one valence electron; neon has eight. Atoms with the same number of valence electrons have similar chemical behaviors. An atom with a completed valence shell, like neon, is nonreactive. All other atoms are chemically reactive because they have incomplete valence shells. The paths of electrons are often portrayed as concentric paths, like planets orbiting the sun. In reality, an electron occupies a more complex three-dimensional space, an orbital. The orbital represents the space in which the electron is found 90% of the time. Each orbital can hold a maximum of two electrons. The first shell has room for a single spherical 1s orbital for its pair of electrons. The second shell can pack pairs of electrons into a spherical 2s orbital and three dumbbell-shaped 2p orbitals. The reactivity of atoms arises from the presence of unpaired electrons in one or more orbitals of their valence shells. Electrons occupy separate orbitals within the valence shell until forced to share orbitals. The four valence electrons of carbon each occupy separate orbitals, but the five valence electrons of nitrogen are distributed into three unshared orbitals and one shared orbital. When atoms interact to complete their valence shells, it is the unpaired electrons that are involved. 3. Atoms combine by chemical bonding to form molecules. Atoms with incomplete valence shells can interact with each other by sharing or transferring valence electrons. These interactions typically result in the atoms remaining close together, held by attractions called chemical bonds. The strongest chemical bonds are covalent bonds and ionic bonds. Reference: Guide for Biology, Campbell, Reece and Mitchell 15 IG Lecture Outlines 1- A covalent bond is formed by the sharing of a pair of valence electrons by two atoms. If two atoms come close enough that their unshared orbitals overlap, they will share their newly paired electrons. Each atom can count both electrons toward its goal of filling the valence shell. For example, if two hydrogen atoms come close enough that their 1s orbitals overlap, then they can share a pair of electrons, with each atom contributing one. Two or more atoms held together by covalent bonds constitute a molecule. We can abbreviate the structure of the molecule by substituting a line for each pair of shared electrons, drawing the structural formula. H—H is the structural formula for the covalent bond between two hydrogen atoms. The molecular formula indicates the number and types of atoms present in a single molecule. H2 is the molecular formula for hydrogen gas. Oxygen needs to add 2 electrons to the 6 already present to complete its valence shell. Two oxygen atoms can form a molecule by sharing two pairs of valence electrons. These atoms have formed a double covalent bond. Every atom has a characteristic total number of covalent bonds that it can form, equal to the number of unpaired electrons in the outermost shell. This bonding capacity is called the atom’s valence. The valence of hydrogen is 1. Oxygen is 2. Nitrogen is 3. Carbon is 4. Phosphorus should have a valence of 3, based on its three unpaired electrons, but in biological molecules it generally has a valence of 5, forming three single covalent bonds and one double bond. Covalent bonds can form between atoms of the same element or atoms of different elements. While both types are molecules, the latter are also compounds. Water, H2O, is a compound in which two hydrogen atoms form single covalent bonds with an oxygen atom. This satisfies the valences of both elements. Methane, CH4, satisfies the valences of both C and H. The attraction of an atom for the shared electrons of a covalent bond is called its electronegativity. Strongly electronegative atoms attempt to pull the shared electrons toward themselves. If electrons in a covalent bond are shared equally, then this is a nonpolar covalent bond. A covalent bond between two atoms of the same element is always nonpolar. A covalent bond between atoms that have similar electronegativities is also nonpolar. Because carbon and hydrogen do not differ greatly in electronegativities, the bonds of CH4 are nonpolar. When two atoms that differ in electronegativity bond, they do not share the electron pair equally and form a polar covalent bond. The bonds between oxygen and hydrogen in water are polar covalent because oxygen has a much higher electronegativity than does hydrogen. Compounds with a polar covalent bond have regions of partial negative charge near the strongly electronegative atom and regions of partial positive charge near the weakly electronegative atom. Reference: Guide for Biology, Campbell, Reece and Mitchell 16 IG Lecture Outlines 1- An ionic bond can form if two atoms are so unequal in their attraction for valence electrons that one atom strips an electron completely from the other. For example, sodium, with one valence electron in its third shell, transfers this electron to chlorine, with 7 valence electrons in its third shell. Now, sodium has a full valence shell (the second) and chlorine has a full valence shell (the third). After the transfer, both atoms are no longer neutral, but have charges and are called ions. Sodium has one more proton than electrons and has a net positive charge. Atoms with positive charges are cations. Chlorine has one more electron than protons and has a net negative charge. Atoms with negative charges are anions. Because of differences in charge, cations and anions are attracted to each other to form an ionic bond. Atoms in an ionic bond need not have acquired their charges by transferring electrons with each other. Compounds formed by ionic bonds are ionic compounds, or salts. An example is NaCl, or table salt. The formula for an ionic compound indicates the ratio of elements in a crystal of that salt. NaCl is not a molecule, but a salt crystal with equal numbers of Na+ and Cl− ions. Ionic compounds can have ratios of elements different from 1:1. For example, the ionic compound magnesium chloride (MgCl2) has 2 chloride atoms per magnesium atom. Magnesium needs to lose 2 electrons to drop to a full outer shell; each chlorine atom needs to gain 1. Entire molecules that have full electrical charges are also called ions. In the salt ammonium chloride (NH4Cl), the anion is Cl− and the cation is NH4+. The strength of ionic bonds depends on environmental conditions, such as moisture. Water can dissolve salts by reducing the attraction between the salt’s anions and cations. 4. Weak chemical bonds play important roles in the chemistry of life. Within a cell, weak, brief bonds between molecules are important to a variety of processes. For example, signal molecules from one neuron use weak bonds to bind briefly to receptor molecules on the surface of a receiving neuron. This triggers a response by the recipient. Weak interactions include ionic bonds (weak in water), hydrogen bonds, and van der Waals interactions. Hydrogen bonds form when a hydrogen atom already covalently bonded to a strongly electronegative atom is attracted to another strongly electronegative atom. These strongly electronegative atoms are typically nitrogen or oxygen. These bonds form because a polar covalent bond leaves the hydrogen atom with a partial positive charge and the other atom with a partial negative charge. The partially positive–charged hydrogen atom is attracted to regions of full or partial negative charge on molecules, atoms, or even regions of the same large molecule. For example, ammonia molecules and water molecules interact with weak hydrogen bonds. In the ammonia molecule, the hydrogen atoms have partial positive charges, and the more electronegative nitrogen atom has a partial negative charge. Reference: Guide for Biology, Campbell, Reece and Mitchell 17 IG Lecture Outlines 1- In the water molecule, the hydrogen atoms also have partial positive charges, and the oxygen atom has a partial negative charge. Areas with opposite charges are attracted. Even molecules with nonpolar covalent bonds can have temporary regions of partial negative and positive charge. Because electrons are constantly in motion, there can be periods when they accumulate by chance in one area of a molecule. This creates ever-changing regions of partial negative and positive charge within a molecule. Molecules or atoms in close proximity can be attracted by these fleeting charge differences, creating van der Waals interactions. While individual bonds (ionic, hydrogen, van der Waals) are weak and temporary, collectively they are strong and play important biological roles. 5. A molecule’s biological function is related to its shape. The three-dimensional shape of a molecule is an important determinant of its function in a cell. A molecule with two atoms is always linear. However, a molecule with more than two atoms has a more complex shape. The shape of a molecule is determined by the positions of the electron orbitals that are shared by the atoms involved in the bond. When covalent bonds form, the orbitals in the valence shell of each atom rearrange. For atoms with electrons in both s and p orbitals, the formation of a covalent bonds leads to hybridization of the orbitals to four new orbitals in a tetrahedral shape. In a water molecule, two of oxygen’s four hybrid orbitals are shared with hydrogen atoms. The water molecule is shaped like a V, with its two covalent bonds spread apart at an angle of 104.5°. In a methane molecule (CH4), the carbon atom shares all four of its hybrid orbitals with H atoms. The carbon nucleus is at the center of the tetrahedron, with hydrogen nuclei at the four corners. Large organic molecules contain many carbon atoms. In these molecules, the tetrahedral shape of carbon bonded to four other atoms is often a repeating motif. Biological molecules recognize and interact with one another with a specificity based on molecular shape. For example, signal molecules from a transmitting cell have specific shapes that bind to complementary receptor molecules on the surface of the receiving cell. The temporary attachment of the receptor and signal molecule stimulates activity in the receptor cell. Molecules with similar shapes can have similar biological effects. For example, morphine, heroin, and other opiate drugs are similar enough in shape that they can bind to the same receptors as natural signal molecules called endorphins. Binding of endorphins to receptors on brain cells produces euphoria and relieves pain. Opiates mimic these natural endorphin effects. 6. Chemical reactions form and break chemical bonds. In chemical reactions, chemical bonds are broken and reformed, leading to new arrangements of atoms. Reference: Guide for Biology, Campbell, Reece and Mitchell 18 IG Lecture Outlines 1- The starting molecules in the process are called reactants, and the final molecules are called products. In a chemical reaction, all of the atoms in the reactants must be present in the products. The reactions must be “balanced.” Matter is conserved in a chemical reaction. Chemical reactions rearrange matter; they do not create or destroy matter. For example, we can recombine the covalent bonds of H2 and O2 to form the new bonds of H2O. In this reaction, two molecules of H2 combine with one molecule of O2 to form two molecules of H2O. Photosynthesis is an important chemical reaction. Humans and other animals ultimately depend on photosynthesis for food and oxygen. Green plants combine carbon dioxide (CO2) from the air and water (H2O) from the soil to create sugar molecules and release molecular oxygen (O2) as a by-product. This chemical reaction is powered by sunlight. The overall process of photosynthesis is 6CO2 + 6H2O -> C6H12O6 + 6O2. This process occurs in a sequence of individual chemical reactions that rearrange the atoms of the reactants to form the products. Some chemical reactions go to completion; that is, all the reactants are converted to products. Most chemical reactions are reversible, with the products in the forward reaction becoming the reactants for the reverse reaction. For example in this reaction: 3H2 + N2 <=> 2NH3 hydrogen and nitrogen molecules combine to form ammonia, but ammonia can decompose to hydrogen and nitrogen molecules. Initially, when reactant concentrations are high, they frequently collide to create products. As products accumulate, they collide to reform reactants. Eventually, the rate of formation of products is the same as the rate of breakdown of products (formation of reactants), and the system is at chemical equilibrium. At equilibrium, products and reactants are continually being formed, but there is no net change in the concentrations of reactants and products. At equilibrium, the concentrations of reactants and products are typically not equal, but their concentrations have stabilized at a particular ratio. Reference: Guide for Biology, Campbell, Reece and Mitchell Reference: Guide for Biology, Campbell, Reece and Mitchell 19 IG Lecture Outlines 1- Chapter 3 Water and the Fitness of the Environment Lecture Outline Overview Because water is the substance that makes life possible on Earth, astronomers hope to find evidence of water on newly discovered planets orbiting distant stars. Life on Earth began in water and evolved there for 3 billion years before colonizing the land. Even terrestrial organisms are tied to water. Most cells are surrounded by water. Cells are about 70–95% water. Water is a reactant in many of the chemical reactions of life. Water is the only common substance that exists in the natural world in all three physical states of matter: solid ice, liquid water, and water vapor. A. The Effects of Water’s Polarity 1. The polarity of water molecules results in hydrogen bonding. In a water molecule, two hydrogen atoms form single polar covalent bonds with an oxygen atom. Because oxygen is more electronegative than hydrogen, the region around the oxygen atom has a partial negative charge. The regions near the two hydrogen atoms have a partial positive charge. A water molecule is a polar molecule in which opposite ends of the molecule have opposite charges. Water has a variety of unusual properties because of the attraction between polar water molecules. The slightly negative regions of one water molecule are attracted to the slightly positive regions of nearby water molecules, forming hydrogen bonds. Each water molecule can form hydrogen bonds with up to four neighbors. 2. Organisms depend on the cohesion of water molecules. The hydrogen bonds joining water molecules are weak, about 1/20 as strong as covalent bonds. They form, break, and reform with great frequency. Each hydrogen bond lasts only a few trillionths of a second. At any instant, a substantial percentage of all water molecules are bonded to their neighbors, creating a high level of structure. Collectively, hydrogen bonds hold water together, a phenomenon called cohesion. Cohesion among water molecules plays a key role in the transport of water and dissolved nutrients against gravity in plants. Water molecules move from the roots to the leaves of a plant through water-conducting vessels. Reference: Guide for Biology, Campbell, Reece and Mitchell 20 IG Lecture Outlines 1- As water molecules evaporate from a leaf, other water molecules from vessels in the leaf replace them. Hydrogen bonds cause water molecules leaving the vessels to tug on molecules farther down. This upward pull is transmitted down to the roots. Adhesion, clinging of one substance to another, contributes too, as water adheres to the wall of the vessels. Surface tension, a measure of the force necessary to stretch or break the surface of a liquid, is related to cohesion. Water has a greater surface tension than most other liquids because hydrogen bonds among surface water molecules resist stretching or breaking the surface. Water behaves as if covered by an invisible film. Some animals can stand, walk, or run on water without breaking the surface. 3. Water moderates temperatures on Earth. Water stabilizes air temperatures by absorbing heat from warmer air and releasing heat to cooler air. Water can absorb or release relatively large amounts of heat with only a slight change in its own temperature. Atoms and molecules have kinetic energy, the energy of motion, because they are always moving. The faster a molecule moves, the more kinetic energy it has. Heat is a measure of the total quantity of kinetic energy due to molecular motion in a body of matter. Temperature measures the intensity of heat in a body of matter due to the average kinetic energy of molecules. As the average speed of molecules increases, a thermometer will record an increase in temperature. Heat and temperature are related, but not identical. When two objects of different temperatures come together, heat passes from the warmer object to the cooler object until the two are the same temperature. Molecules in the cooler object speed up at the expense of kinetic energy of the warmer object. Ice cubes cool a glass of pop by absorbing heat from the pop as the ice melts. In most biological settings, temperature is measured on the Celsius scale (°C). At sea level, water freezes at 0°C and boils at 100°C. Human body temperature is typically 37°C. While there are several ways to measure heat energy, one convenient unit is the calorie (cal). One calorie is the amount of heat energy necessary to raise the temperature of one g of water by 1°C. A calorie is released when 1 g of water cools by 1°C. In many biological processes, the kilocalorie (kcal) is more convenient. A kilocalorie is the amount of heat energy necessary to raise the temperature of 1000 g of water by 1°C. Another common energy unit, the joule (J), is equivalent to 0.239 cal. Water stabilizes temperature because it has a high specific heat. The specific heat of a substance is the amount of heat that must be absorbed or lost for 1 g of that substance to change its temperature by 1°C. Reference: Guide for Biology, Campbell, Reece and Mitchell 21 IG Lecture Outlines 1- By definition, the specific heat of water is 1 cal per gram per degree Celsius or 1 cal/g/°C. Water has a high specific heat compared to other substances. For example, ethyl alcohol has a specific heat of 0.6 cal/g/°C. The specific heat of iron is 1/10 that of water. Water resists changes in temperature because of its high specific heat. In other words, water absorbs or releases a relatively large quantity of heat for each degree of temperature change. Water’s high specific heat is due to hydrogen bonding. Heat must be absorbed to break hydrogen bonds, and heat is released when hydrogen bonds form. Investment of one calorie of heat causes relatively little change to the temperature of water because much of the energy is used to disrupt hydrogen bonds, not speed up the movement of water molecules. Water’s high specific heat has effects that range from the level of the whole Earth to the level of individual organisms. A large body of water can absorb a large amount of heat from the sun in daytime during the summer and yet warm only a few degrees. At night and during the winter, the warm water will warm cooler air. Therefore, ocean temperatures and coastal land areas have more stable temperatures than inland areas. Living things are made primarily of water. Consequently, they resist changes in temperature better than they would if composed of a liquid with a lower specific heat. The transformation of a molecule from a liquid to a gas is called vaporization or evaporation. This occurs when the molecule moves fast enough to overcome the attraction of other molecules in the liquid. Even in a low-temperature liquid (with low average kinetic energy), some molecules are moving fast enough to evaporate. Heating a liquid increases the average kinetic energy and increases the rate of evaporation. Heat of vaporization is the quantity of heat that a liquid must absorb for 1 g of it to be converted from liquid to gas. Water has a relatively high heat of vaporization, requiring about 580 cal of heat to evaporate 1 g of water at room temperature. This is double the heat required to vaporize the same quantity of alcohol or ammonia. This is because hydrogen bonds must be broken before a water molecule can evaporate from the liquid. Water’s high heat of vaporization moderates climate. Much of the sun’s heat absorbed by tropical oceans is used for evaporation of surface water. As moist tropical air moves to the poles, water vapor condenses to form rain, releasing heat. As a liquid evaporates, the surface of the liquid that remains behind cools, a phenomenon called evaporative cooling. This occurs because the most energetic molecules are the most likely to evaporate, leaving the lower–kinetic energy molecules behind. Evaporative cooling moderates temperature in lakes and ponds. Evaporation of sweat in mammals or evaporation of water from the leaves of plants prevents terrestrial organisms from overheating. Reference: Guide for Biology, Campbell, Reece and Mitchell 22 IG Lecture Outlines 1- Evaporation of water from the leaves of plants or the skin of humans removes excess heat. 4. Oceans and lakes don’t freeze solid because ice floats. Water is unusual because it is less dense as a solid than as a cold liquid. Most materials contract as they solidify, but water expands. At temperatures above 4°C, water behaves like other liquids, expanding as it warms and contracting as it cools. Water begins to freeze when its molecules are no longer moving vigorously enough to break their hydrogen bonds. When water reaches 0°C, water becomes locked into a crystalline lattice, with each water molecule bonded to a maximum of four partners. As ice starts to melt, some of the hydrogen bonds break, and water molecules can slip closer together than they can while in the ice state. Ice is about 10% less dense than water at 4°C. Therefore, ice floats on the cool water below. This oddity has important consequences for life. If ice sank, eventually all ponds, lakes, and even the ocean would freeze solid. During the summer, only the upper few centimeters of the ocean would thaw. Instead, the surface layer of ice insulates liquid water below, preventing it from freezing and allowing life to exist under the frozen surface. 5. Water is the solvent of life. A liquid that is a completely homogeneous mixture of two or more substances is called a solution. A sugar cube in a glass of water will eventually dissolve to form a uniform solution of sugar and water. The dissolving agent is the solvent, and the substance that is dissolved is the solute. In our example, water is the solvent and sugar the solute. In an aqueous solution, water is the solvent. Water is not a universal solvent, but it is very versatile because of the polarity of water molecules. Water is an effective solvent because it readily forms hydrogen bonds with charged and polar covalent molecules. For example, when a crystal of salt (NaCl) is placed in water, the Na+ cations interact with the partial negative charges of the oxygen regions of water molecules. The Cl− anions interact with the partial positive charges of the hydrogen regions of water molecules. Each dissolved ion is surrounded by a sphere of water molecules, a hydration shell. Eventually, water dissolves all the ions, resulting in a solution with two solutes: sodium and chloride ions. Polar molecules are also soluble in water because they form hydrogen bonds with water. Even large molecules, like proteins, can dissolve in water if they have ionic and polar regions. Any substance that has an affinity for water is hydrophilic (water-loving). These substances are dominated by ionic or polar bonds. Some hydrophilic substances do not dissolve because their molecules are too large. Reference: Guide for Biology, Campbell, Reece and Mitchell 23 IG Lecture Outlines 1- For example, cotton is hydrophilic because cellulose, its major constituent, has numerous polar covalent bonds. However, its giant cellulose molecules are too large to dissolve in water. Water molecules form hydrogen bonds with the cellulose fibers of cotton, allowing you to dry yourself with your cotton towel as the water is pulled into the towel. Substances that have no affinity for water are hydrophobic (water-fearing). These substances are nonionic and have nonpolar covalent bonds. Because there are no consistent regions with partial or full charges, water molecules cannot form hydrogen bonds with hydrophobic molecules. Oils such as vegetable oil are hydrophobic because the dominant bonds, carbon-carbon and carbon-hydrogen, share electrons equally. Hydrophobic molecules are major ingredients of cell membranes. Biological chemistry is “wet” chemistry with most reactions involving solutes dissolved in water. Chemical reactions depend on collisions of molecules and therefore on the concentrations of solutes in aqueous solution. We measure the number of molecules in units called moles. The actual number of molecules in a mole is called Avogadro’s number, 6.02 × 1023. A mole is equal to the molecular weight of a substance but scaled up from daltons to grams. To illustrate, how could we measure out a mole of table sugar—sucrose (C12H22O11)? A carbon atom weighs 12 daltons, hydrogen 1 dalton, and oxygen 16 daltons. One molecule of sucrose would weigh 342 daltons, the sum of weights of all the atoms in sucrose, or the molecular weight of sucrose. To get one mole of sucrose, we would weigh out 342 g. The advantage of using moles as a measurement is that a mole of one substance has the same number of molecules as a mole of any other substance. If substance A has a molecular weight of 10 daltons and substance B has a molecular weight of 100 daltons, then we know that 10 g of substance A has the same number of molecules as 100 g of substance B. A mole of sucrose contains 6.02 × 1023 molecules and weighs 342 g, while a mole of ethyl alcohol (C2H6O) also contains 6.02 × 1023 molecules but weighs only 46 g because the molecules are smaller. Measuring in moles allows scientists to combine substances in fixed ratios of molecules. In “wet” chemistry, we are typically combining solutions or measuring the quantities of materials in aqueous solutions. The concentration of a material in solution is called its molarity. A one molar solution has one mole of a substance dissolved in one liter of solvent, typically water. To make a 1 molar (1M) solution of sucrose, we would slowly add water to 342 g of sucrose until the total volume was 1 liter and all the sugar was dissolved. B. The Dissociation of Water Molecules Occasionally, a hydrogen atom participating in a hydrogen bond between two water molecules shifts from one molecule to the other. The hydrogen atom leaves its electron behind and is transferred as a single proton—a hydrogen ion (H+). The water molecule that lost the proton is now a hydroxide ion (OH−). Reference: Guide for Biology, Campbell, Reece and Mitchell 24 IG Lecture Outlines 1- The water molecule with the extra proton is now a hydronium ion (H3O+). A simplified way to view this process is to say that a water molecule dissociates into a hydrogen ion and a hydroxide ion: H2O <=> H+ + OH− This reaction is reversible. At equilibrium, the concentration of water molecules greatly exceeds that of H+ and OH−. In pure water, only one water molecule in every 554 million is dissociated. At equilibrium, the concentration of H+ or OH− is 10−7M (at 25°C). Although the dissociation of water is reversible and statistically rare, it is very important in the chemistry of life. Because hydrogen and hydroxide ions are very reactive, changes in their concentrations can drastically affect the chemistry of a cell. Adding certain solutes, called acids and bases, disrupts the equilibrium and modifies the concentrations of hydrogen and hydroxide ions. The pH scale is used to describe how acidic or basic a solution is. 1. Organisms are sensitive to changes in pH. An acid is a substance that increases the hydrogen ion concentration in a solution. When hydrochloric acid is added to water, hydrogen ions dissociate from chloride ions: HCl -> H+ + Cl− Addition of an acid makes a solution more acidic. Any substance that reduces the hydrogen ion concentration in a solution is a base. Some bases reduce the H+ concentration directly by accepting hydrogen ions. Ammonia (NH3) acts as a base when the nitrogen’s unshared electron pair attracts a hydrogen ion from the solution, creating an ammonium ion (NH4+). NH3 + H+ <=> NH4+ Other bases reduce H+ indirectly by dissociating to OH−, which then combines with H+ to form water. NaOH -> Na+ + OH− OH− + H+ -> H2O Solutions with more OH− than H+ are basic solutions. Solutions with more H+ than OH− are acidic solutions. Solutions in which concentrations of OH− and H+ are equal are neutral solutions. Some acids and bases (HCl and NaOH) are strong acids or bases. These molecules dissociate completely in water. Other acids and bases (NH3) are weak acids or bases. For these molecules, the binding and release of hydrogen ions are reversible. At equilibrium, there will be a fixed ratio of products to reactants. Carbonic acid (H2CO3) is a weak acid: H2CO3 <=> HCO3− + H+ At equilibrium, 1% of the H2CO3 molecules will be dissociated. In any solution, the product of the H+ and OH− concentrations is constant at 10−14. Brackets ([H+] and [OH−]) indicate the molar concentration of the enclosed substance. Reference: Guide for Biology, Campbell, Reece and Mitchell 25 IG Lecture Outlines 1- [H+] [OH−] = 10−14 In a neutral solution, [H+] = 10−7 M and [OH−] = 10−7 M Adding acid to a solution shifts the balance between H+ and OH− toward H+ and leads to a decline in OH−. If [H+] = 10−5 M, then [OH−] = 10−9 M Hydroxide concentrations decline because some of the additional acid combines with hydroxide to form water. Adding a base does the opposite, increasing OH− concentration and lowering H+ concentration. The H+ and OH− concentrations of solutions can vary by a factor of 100 trillion or more. To express this variation more conveniently, the H+ and OH− concentrations are typically expressed via the pH scale. The pH scale, ranging from 1 to 14, compresses the range of concentrations by employing logarithms. pH = − log [H+] or [H+] = 10−pH In a neutral solution, [H+] = 10−7 M, and the pH = 7. Values for pH decline as [H+] increase. While the pH scale is based on [H+], values for [OH−] can be easily calculated from the product relationship. The pH of a neutral solution is 7. Acidic solutions have pH values less than 7, and basic solutions have pH values greater than 7. Most biological fluids have pH values in the range of 6 to 8. However, the human stomach has strongly acidic digestive juice with a pH of about 2. Each pH unit represents a tenfold difference in H+ and OH− concentrations. A small change in pH actually indicates a substantial change in H+ and OH− concentrations. The chemical processes in the cell can be disrupted by changes to the H+ and OH− concentrations away from their normal values, usually near pH 7. To maintain cellular pH values at a constant level, biological fluids have buffers. Buffers resist changes to the pH of a solution when H+ or OH− is added to the solution. Buffers accept hydrogen ions from the solution when they are in excess and donate hydrogen ions when they have been depleted. Buffers typically consist of a weak acid and its corresponding base. One important buffer in human blood and other biological solutions is carbonic acid, which dissociates to yield a bicarbonate ion and a hydrogen ion. The chemical equilibrium between carbonic acid and bicarbonate acts as a pH regulator. The equilibrium shifts left or right as other metabolic processes add or remove H+ from the solution. 2. Acid precipitation threatens the fitness of the environment. Acid precipitation is a serious assault on water quality in some industrialized areas. Uncontaminated rain has a slightly acidic pH of 5.6. The acid is a product of the formation of carbonic acid from carbon dioxide and water. Acid precipitation occurs when rain, snow, or fog has a pH that is more acidic than 5.6. Acid precipitation is caused primarily by sulfur oxides and nitrogen oxides in the atmosphere. Reference: Guide for Biology, Campbell, Reece and Mitchell 26 IG Lecture Outlines 1- These molecules react with water to form strong acids that fall to the surface with rain or snow. The major source of these oxides is the burning of fossil fuels (coal, oil, and gas) in factories and automobiles. The presence of tall smokestacks allows this pollution to spread from its site of origin to contaminate relatively pristine areas thousands of kilometers away. In 2001, rain in the Adirondack Mountains of upstate New York had an average pH of 4.3. The effects of acids in lakes and streams are more pronounced in the spring during snowmelt. As the surface snows melt and drain down through the snowfield, the meltwater accumulates acid and brings it into lakes and streams all at once. The pH of early meltwater may be as low as 3. Acid precipitation has a great impact on the eggs and the early developmental stages of aquatic organisms that are abundant in the spring. Thus, strong acidity can alter the structure of molecules and impact ecological communities. Direct impacts of acid precipitation on forests and terrestrial life are more controversial. However, acid precipitation can impact soils by affecting the solubility of soil minerals. Acid precipitation can wash away key soil buffers and plant nutrients such as calcium and magnesium ions. It can also increase the concentrations of compounds such as aluminum to toxic levels. This has done major damage to forests in Europe and substantial damage of forests in North America. Progress has been made in reducing acid precipitation. Reference: Guide for Biology, Campbell, Reece and Mitchell Reference: Guide for Biology, Campbell, Reece and Mitchell 27 IG Lecture Outlines 1- Chapter 4 Carbon and the Molecular Diversity of Life Lecture Outline Overview A. Carbon—The Backbone of Biological Molecules Although cells are 70–95% water, the rest consists mostly of carbon-based compounds. Carbon is unparalleled in its ability to form large, complex, and diverse molecules. Carbon accounts for the diversity of biological molecules and has made possible the great diversity of living things. Proteins, DNA, carbohydrates, and other molecules that distinguish living matter from inorganic material are all composed of carbon atoms bonded to each other and to atoms of other elements. These other elements commonly include hydrogen (H), oxygen (O), nitrogen (N), sulfur (S), and phosphorus (P). 1. Organic chemistry is the study of carbon compounds. The study of carbon compounds, organic chemistry, deals with any compound with carbon (organic compounds). Organic compounds can range from simple molecules, such as CO2 or CH4, to complex molecules such as proteins, which may weigh more than 100,000 daltons. The overall percentages of the major elements of life (C, H, O, N, S, and P) are quite uniform from one organism to another. However, because of carbon’s versatility, these few elements can be combined to build an inexhaustible variety of organic molecules. Variations in organic molecules can distinguish even between individuals of a single species. The science of organic chemistry began in attempts to purify and improve the yield of products obtained from other organisms. Initially, chemists learned to synthesize simple compounds in the laboratory, but had no success with more complex compounds. The Swedish chemist Jons Jacob Berzelius was the first to make a distinction between organic compounds that seemed to arise only in living organisms and inorganic compounds that were found in the nonliving world. This led early organic chemists to propose vitalism, the belief that physical and chemical laws did not apply to living things. Support for vitalism began to wane as organic chemists learned to synthesize complex organic compounds in the laboratory. In the early 1800s, the German chemist Friedrich Wöhler and his students were able to synthesize urea from totally inorganic materials. Reference: Guide for Biology, Campbell, Reece and Mitchell 28 IG Lecture Outlines 1- In 1953, Stanley Miller at the University of Chicago set up a laboratory simulation of chemical conditions on the primitive Earth and demonstrated the spontaneous synthesis of organic compounds. Such spontaneous synthesis of organic compounds may have been an early stage in the origin of life. Organic chemists finally rejected vitalism and embraced mechanism, accepting that the same physical and chemical laws govern all natural phenomena including the processes of life. Organic chemistry was redefined as the study of carbon compounds regardless of their origin. Organisms do produce the majority of organic compounds. The laws of chemistry apply to inorganic and organic compounds alike. 2. Carbon atoms can form diverse molecules by bonding to four other atoms. With a total of 6 electrons, a carbon atom has 2 in the first electron shell and 4 in the second shell. Carbon has little tendency to form ionic bonds by losing or gaining 4 electrons to complete its valence shell. Instead, carbon usually completes its valence shell by sharing electrons with other atoms in four covalent bonds. This tetravalence by carbon makes large, complex molecules possible. When carbon forms covalent bonds with four other atoms, they are arranged at the corners of an imaginary tetrahedron with bond angles of 109.5°. In molecules with multiple carbons, every carbon bonded to four other atoms has a tetrahedral shape. However, when two carbon atoms are joined by a double bond, all bonds around those carbons are in the same plane and have a flat, three-dimensional structure. The three-dimensional shape of an organic molecule determines its function. The electron configuration of carbon makes it capable of forming covalent bonds with many different elements. The valences of carbon and its partners can be viewed as the building code that governs the architecture of organic molecules. In carbon dioxide, one carbon atom forms two double bonds with two different oxygen atoms. In the structural formula, O=C=O, each line represents a pair of shared electrons. This arrangement completes the valence shells of all atoms in the molecule. While CO2 can be classified as either organic or inorganic, its importance to the living world is clear. CO2 is the source of carbon for all organic molecules found in organisms. It is usually fixed into organic molecules by the process of photosynthesis. Urea, CO(NH2)2, is another simple organic molecule in which each atom forms covalent bonds to complete its valence shell. 3. Variation in carbon skeletons contributes to the diversity of organic molecules. Carbon chains form the skeletons of most organic molecules. The skeletons vary in length and may be straight, branched, or arranged in closed rings. The carbon skeletons may include double bonds. Atoms of other elements can be bonded to the atoms of the carbon skeleton. Hydrocarbons are organic molecules that consist of only carbon and hydrogen atoms. Reference: Guide for Biology, Campbell, Reece and Mitchell 29 IG Lecture Outlines 1- Hydrocarbons are the major component of petroleum, a fossil fuel that consists of the partially decomposed remains of organisms that lived millions of years ago. Fats are biological molecules that have long hydrocarbon tails attached to a nonhydrocarbon component. Petroleum and fat are hydrophobic compounds that cannot dissolve in water because of their many nonpolar carbon-to-hydrogen bonds. Isomers are compounds that have the same molecular formula but different structures and, therefore, different chemical properties. For example, butane and isobutane have the same molecular formula, C4H10, but butane has a straight skeleton and isobutane has a branched skeleton. The two butanes are structural isomers, molecules that have the same molecular formula but differ in the covalent arrangement of atoms. Geometric isomers are compounds with the same covalent partnerships that differ in the spatial arrangement of atoms around a carbon–carbon double bond. The double bond does not allow atoms to rotate freely around the bond axis. The biochemistry of vision involves a light-induced change in the structure of rhodopsin in the retina from one geometric isomer to another. Enantiomers are molecules that are mirror images of each other. Enantiomers are possible when four different atoms or groups of atoms are bonded to a carbon. In this case, the four groups can be arranged in space in two different ways that are mirror images. They are like left-handed and right-handed versions of the molecule. Usually one is biologically active, while the other is inactive. Even subtle structural differences in two enantiomers have important functional significance because of emergent properties from specific arrangements of atoms. One enantiomer of the drug thalidomide reduced morning sickness, the desired effect, but the other isomer caused severe birth defects. The L-dopa isomer is an effective treatment of Parkinson’s disease, but the D-dopa isomer is inactive. B. Functional Groups 1. Functional groups are involved in chemical reactions and contribute to the molecular diversity of life. The components of organic molecules that are most commonly involved in chemical reactions are known as functional groups. If we consider hydrocarbons to be the simplest organic molecules, we can view functional groups as attachments that replace one or more of the hydrogen atoms bonded to the carbon skeleton of the hydrocarbon. Each functional group behaves consistently from one organic molecule to another. The number and arrangement of functional groups help give each molecule its unique properties. As an example, the basic structure of testosterone (a male sex hormone) and estradiol (a female sex hormone) is the same. Both are steroids with four fused carbon rings, but they differ in the functional groups attached to the rings. Reference: Guide for Biology, Campbell, Reece and Mitchell 30 IG Lecture Outlines 1- These functional groups interact with different targets in the body. There are six functional groups that are most important to the chemistry of life: hydroxyl, carbonyl, carboxyl, amino, sulfhydryl, and phosphate groups. All are hydrophilic and increase the solubility of organic compounds in water. In a hydroxyl group (—OH), a hydrogen atom forms a polar covalent bond with an oxygen atom, which forms a polar covalent bond to the carbon skeleton. Because of these polar covalent bonds, hydroxyl groups increase the solubility of organic molecules. Organic compounds with hydroxyl groups are alcohols, and their names typically end in -ol. A carbonyl group (>CO) consists of an oxygen atom joined to the carbon skeleton by a double bond. If the carbonyl group is on the end of the skeleton, the compound is an aldehyde. If the carbonyl group is within the carbon skeleton, then the compound is a ketone. Isomers with aldehydes versus ketones have different properties. A carboxyl group (—COOH) consists of a carbon atom with a double bond to an oxygen atom and a single bond to the oxygen of a hydroxyl group. Compounds with carboxyl groups are carboxylic acids. A carboxyl group acts as an acid because the combined electronegativities of the two adjacent oxygen atoms increase the dissociation of hydrogen as an ion (H+). An amino group (—NH2) consists of a nitrogen atom bonded to two hydrogen atoms and the carbon skeleton. Organic compounds with amino groups are amines. The amino group acts as a base because the amino group can pick up a hydrogen ion (H+) from the solution. Amino acids, the building blocks of proteins, have amino and carboxyl groups. A sulfhydryl group (—SH) consists of a sulfur atom bonded to a hydrogen atom and to the backbone. This group resembles a hydroxyl group in shape. Organic molecules with sulfhydryl groups are thiols. Two sulfhydryl groups can interact to help stabilize the structure of proteins. A phosphate group (—OPO32−) consists of a phosphorus atom bound to four oxygen atoms (three with single bonds and one with a double bond). A phosphate group connects to the carbon backbone via one of its oxygen atoms. Phosphate groups are anions with two negative charges, as two protons have dissociated from the oxygen atoms. One function of phosphate groups is to transfer energy between organic molecules. Adenosine triphosphate, or ATP, is the primary energy-transferring molecule in living cells. 2. These are the chemical elements of life. Living matter consists mainly of carbon, oxygen, hydrogen, and nitrogen, with smaller amounts of sulfur and phosphorus. These elements are linked by strong covalent bonds. Carbon, with its four covalent bonds, is the basic building block in molecular architecture. Reference: Guide for Biology, Campbell, Reece and Mitchell 31 IG Lecture Outlines 1- The great diversity of organic molecules with their special properties emerges from the unique arrangement of the carbon skeleton and the functional groups attached to the skeleton. Reference: Guide for Biology, Campbell, Reece and Mitchell Reference: Guide for Biology, Campbell, Reece and Mitchell 32 IG Lecture Outlines 1- Chapter 5 The Structure and Function of Macromolecules Lecture Outline Overview Within cells, small organic molecules are joined together to form larger molecules. These large macromolecules may consist of thousands of covalently bonded atoms and weigh more than 100,000 daltons. The four major classes of macromolecules are carbohydrates, lipids, proteins, and nucleic acids. A. The Principles of Polymers 1. Most macromolecules are polymers. Three of the four classes of macromolecules—carbohydrates, proteins, and nucleic acids—form chainlike molecules called polymers. A polymer is a long molecule consisting of many similar or identical building blocks linked by covalent bonds. The repeated units are small molecules called monomers. Some of the molecules that serve as monomers have other functions of their own. The chemical mechanisms that cells use to make and break polymers are similar for all classes of macromolecules. Monomers are connected by covalent bonds that form through the loss of a water molecule. This reaction is called a condensation reaction or dehydration reaction. When a bond forms between two monomers, each monomer contributes part of the water molecule that is lost. One monomer provides a hydroxyl group (—OH), while the other provides a hydrogen (—H). Cells invest energy to carry out dehydration reactions. The process is aided by enzymes. The covalent bonds connecting monomers in a polymer are disassembled by hydrolysis, a reaction that is effectively the reverse of dehydration. In hydrolysis, bonds are broken by the addition of water molecules. A hydrogen atom attaches to one monomer, and a hydroxyl group attaches to the adjacent monomer. Our food is taken in as organic polymers that are too large for our cells to absorb. Within the digestive tract, various enzymes direct hydrolysis of specific polymers. The resulting monomers are absorbed by the cells lining the gut and transported to the bloodstream for distribution to body cells. The body cells then use dehydration reaction to assemble the monomers into new polymers that carry out functions specific to the particular cell type. 2. An immense variety of polymers can be built from a small set of monomers. Reference: Guide for Biology, Campbell, Reece and Mitchell 33 IG Lecture Outlines 1- Each cell has thousands of different kinds of macromolecules. These molecules vary among cells of the same individual. They vary more among unrelated individuals of a species, and even more between species. This diversity comes from various combinations of the 40–50 common monomers and some others that occur rarely. These monomers can be connected in a great many combinations, just as the 26 letters in the alphabet can be used to create a great diversity of words. B. Carbohydrates Serve as Fuel and Building Material Carbohydrates include sugars and their polymers. The simplest carbohydrates are monosaccharides, or simple sugars. Disaccharides, or double sugars, consist of two monosaccharides joined by a condensation reaction. Polysaccharides are polymers of many monosaccharides. 1. Sugars, the smallest carbohydrates, serve as fuel and a source of carbon. Monosaccharides generally have molecular formulas that are some multiple of the unit CH2O. For example, glucose has the formula C6H12O6. Monosaccharides have a carbonyl group (>C=O) and multiple hydroxyl groups (—OH). Depending on the location of the carbonyl group, the sugar is an aldose or a ketose. Most names for sugars end in -ose. Glucose, an aldose, and fructose, a ketose, are structural isomers. Monosaccharides are also classified by the number of carbons in the carbon skeleton. Glucose and other six-carbon sugars are hexoses. Five-carbon backbones are pentoses; three-carbon sugars are trioses. Monosaccharides may also exist as enantiomers. For example, glucose and galactose, both six-carbon aldoses, differ in the spatial arrangement of their parts around asymmetrical carbons. Monosaccharides, particularly glucose, are a major fuel for cellular work. They also function as the raw material for the synthesis of other monomers, such as amino acids and fatty acids. While often drawn as a linear skeleton, monosaccharides in aqueous solutions form rings. Two monosaccharides can join with a glycosidic linkage to form a disaccharide via dehydration. Maltose, malt sugar, is formed by joining two glucose molecules. Sucrose, table sugar, is formed by joining glucose and fructose. Sucrose is the major transport form of sugars in plants. Lactose, milk sugar, is formed by joining glucose and galactose. 2. Polysaccharides, the polymers of sugars, have storage and structural roles. Polysaccharides are polymers of hundreds to thousands of monosaccharides joined by glycosidic linkages. Some polysaccharides serve for storage and are hydrolyzed as sugars are needed. Other polysaccharides serve as building materials for the cell or the whole organism. Starch is a storage polysaccharide composed entirely of glucose monomers. Reference: Guide for Biology, Campbell, Reece and Mitchell 34 IG Lecture Outlines 1- Most of these monomers are joined by 1–4 linkages (number 1 carbon to number 4 carbon) between the glucose molecules. The simplest form of starch, amylose, is unbranched and forms a helix. Branched forms such as amylopectin are more complex. Plants store surplus glucose as starch granules within plastids, including chloroplasts, and withdraw it as needed for energy or carbon. Animals that feed on plants, especially parts rich in starch, have digestive enzymes that can hydrolyze starch to glucose. Animals store glucose in a polysaccharide called glycogen. Glycogen is highly branched like amylopectin. Humans and other vertebrates store a day’s supply of glycogen in the liver and muscles. Cellulose is a major component of the tough wall of plant cells. Plants produce almost one hundred billion tons of cellulose per year. It is the most abundant organic compound on Earth. Like starch, cellulose is a polymer of glucose. However, the glycosidic linkages in these two polymers differ. The difference is based on the fact that there are actually two slightly different ring structures for glucose. These two ring forms differ in whether the hydroxyl group attached to the number 1 carbon is fixed above (beta glucose) or below (alpha glucose) the plane of the ring. Starch is a polysaccharide of alpha glucose monomers. Cellulose is a polysaccharide of beta glucose monomers, making every other glucose monomer upside down with respect to its neighbors. The differing glycosidic links in starch and cellulose give the two molecules distinct threedimensional shapes. While polymers built with alpha glucose form helical structures, polymers built with beta glucose form straight structures. The straight structures built with beta glucose allow H atoms on one strand to form hydrogen bonds with OH groups on other strands. In plant cell walls, parallel cellulose molecules held together in this way are grouped into units called microfibrils, which form strong building materials for plants (and for humans, as lumber). The enzymes that digest starch by hydrolyzing its alpha linkages cannot hydrolyze the beta linkages in cellulose. Cellulose in human food passes through the digestive tract and is eliminated in feces as “insoluble fiber.” As it travels through the digestive tract, cellulose abrades the intestinal walls and stimulates the secretion of mucus, aiding in the passage of food. Some microbes can digest cellulose to its glucose monomers through the use of cellulase enzymes. Many eukaryotic herbivores, from cows to termites, have symbiotic relationships with cellulolytic microbes, providing the microbe and the host animal access to a rich source of energy. Some fungi can also digest cellulose. Another important structural polysaccharide is chitin, used in the exoskeletons of arthropods (including insects, spiders, and crustaceans). Reference: Guide for Biology, Campbell, Reece and Mitchell 35 IG Lecture Outlines 1- Chitin is similar to cellulose, except that it contains a nitrogen-containing appendage on each glucose monomer. Pure chitin is leathery but can be hardened by the addition of calcium carbonate. Chitin also provides structural support for the cell walls of many fungi. C. Lipids Are a Diverse Group of Hydrophobic Molecules Unlike other macromolecules, lipids do not form polymers. The unifying feature of lipids is that they all have little or no affinity for water. This is because they consist mostly of hydrocarbons, which form nonpolar covalent bonds. Lipids are highly diverse in form and function. 1. Fats store large amounts of energy. Although fats are not strictly polymers, they are large molecules assembled from smaller molecules by dehydration reactions. A fat is constructed from two kinds of smaller molecules: glycerol and fatty acids. Glycerol is a three-carbon alcohol with a hydroxyl group attached to each carbon. A fatty acid consists of a carboxyl group attached to a long carbon skeleton, often 16 to 18 carbons long. The many nonpolar C—H bonds in the long hydrocarbon skeleton make fats hydrophobic. Fats separate from water because the water molecules hydrogen bond to one another and exclude the fats. In a fat, three fatty acids are joined to glycerol by an ester linkage, creating a triacylglycerol, or triglyceride. The three fatty acids in a fat can be the same or different. Fatty acids may vary in length (number of carbons) and in the number and locations of double bonds. If the fatty acid has no carbon-carbon double bonds, then the molecule is a saturated fatty acid, saturated with hydrogens at every possible position. If the fatty acid has one or more carbon-carbon double bonds formed by the removal of hydrogen atoms from the carbon skeleton, then the molecule is an unsaturated fatty acid. A saturated fatty acid is a straight chain, but an unsaturated fatty acid has a kink wherever there is a double bond. Fats made from saturated fatty acids are saturated fats. Most animal fats are saturated. Saturated fats are solid at room temperature. Fats made from unsaturated fatty acids are unsaturated fats. Plant and fish fats are liquid at room temperature and are known as oils. The kinks caused by the double bonds prevent the molecules from packing tightly enough to solidify at room temperature. The phrase “hydrogenated vegetable oils” on food labels means that unsaturated fats have been synthetically converted to saturated fats by the addition of hydrogen. Peanut butter and margarine are hydrogenated to prevent lipids from separating out as oil. A diet rich in saturated fats may contribute to cardiovascular disease (atherosclerosis) through plaque deposits. Reference: Guide for Biology, Campbell, Reece and Mitchell 36 IG Lecture Outlines 1- The process of hydrogenating vegetable oils produces saturated fats and also unsaturated fats with trans double bonds. These trans fat molecules contribute more than saturated fats to atherosclerosis. The major function of fats is energy storage. A gram of fat stores more than twice as much energy as a gram of a polysaccharide such as starch. Because plants are immobile, they can function with bulky energy storage in the form of starch. Plants use oils when dispersal and compact storage is important, as in seeds. Animals must carry their energy stores with them and benefit from having a more compact fuel reservoir of fat. Humans and other mammals store fats as long-term energy reserves in adipose cells that swell and shrink as fat is deposited or withdrawn from storage. Adipose tissue also functions to cushion vital organs, such as the kidneys. A layer of fat can also function as insulation. This subcutaneous layer is especially thick in whales, seals, and most other marine mammals. 2. Phospholipids are major components of cell membranes. Phospholipids have two fatty acids attached to glycerol and a phosphate group at the third position. The phosphate group carries a negative charge. Additional smaller groups may be attached to the phosphate group to form a variety of phospholipids. The interaction of phospholipids with water is complex. The fatty acid tails are hydrophobic, but the phosphate group and its attachments form a hydrophilic head. When phospholipids are added to water, they self-assemble into assemblages with the hydrophobic tails pointing toward the interior. This type of structure is called a micelle. Phospholipids are arranged as a bilayer at the surface of a cell. Again, the hydrophilic heads are on the outside of the bilayer, in contact with the aqueous solution, and the hydrophobic tails point toward the interior of the bilayer. The phospholipid bilayer forms a barrier between the cell and the external environment. Phospholipids are the major component of all cell membranes. 3. Steroids include cholesterol and certain hormones. Steroids are lipids with a carbon skeleton consisting of four fused rings. Different steroids are created by varying functional groups attached to the rings. Cholesterol, an important steroid, is a component in animal cell membranes. Cholesterol is also the precursor from which all other steroids are synthesized. Many of these other steroids are hormones, including the vertebrate sex hormones. While cholesterol is an essential molecule in animals, high levels of cholesterol in the blood may contribute to cardiovascular disease. Both saturated fats and trans fats exert their negative impact on health by affecting cholesterol levels. D. Proteins Have Many Structures and Many Functions Reference: Guide for Biology, Campbell, Reece and Mitchell 37 IG Lecture Outlines 1- Proteins account for more than 50% of the dry mass of most cells. They are instrumental in almost everything that an organism does. Protein functions include structural support, storage, transport, cellular signaling, movement, and defense against foreign substances. Most important, protein enzymes function as catalysts in cells, regulating metabolism by selectively accelerating chemical reactions without being consumed. Humans have tens of thousands of different proteins, each with a specific structure and function. Proteins are the most structurally complex molecules known. Each type of protein has a complex three-dimensional shape or conformation. All protein polymers are constructed from the same set of 20 amino acid monomers. Polymers of proteins are called polypeptides. A protein consists of one or more polypeptides folded and coiled into a specific conformation. 1. Amino acids are the monomers from which proteins are constructed. Amino acids are organic molecules with both carboxyl and amino groups. At the center of an amino acid is an asymmetric carbon atom called the alpha carbon. Four components are attached to the alpha carbon: a hydrogen atom, a carboxyl group, an amino group, and a variable R group (or side chain). Different R groups characterize the 20 different amino acids. R groups may be as simple as a hydrogen atom (as in the amino acid glycine), or it may be a carbon skeleton with various functional groups attached (as in glutamine). The physical and chemical properties of the R group determine the unique characteristics of a particular amino acid. One group of amino acids has hydrophobic R groups. Another group of amino acids has polar R groups that are hydrophilic. A third group of amino acids includes those with functional groups that are charged (ionized) at cellular pH. Some acidic R groups are negative in charge due to the presence of a carboxyl group. Basic R groups have amino groups that are positive in charge. Note that all amino acids have carboxyl and amino groups. The terms acidic and basic in this context refer only to these groups in the R groups. Amino acids are joined together when a dehydration reaction removes a hydroxyl group from the carboxyl end of one amino acid and a hydrogen from the amino group of another. The resulting covalent bond is called a peptide bond. Repeating the process over and over creates a polypeptide chain. At one end is an amino acid with a free amino group (the N-terminus) and at the other is an amino acid with a free carboxyl group (the C-terminus). Polypeptides range in size from a few monomers to thousands. Each polypeptide has a unique linear sequence of amino acids. 2. The amino acid sequence of a polypeptide can be determined. Frederick Sanger and his colleagues at Cambridge University determined the amino acid sequence of insulin in the 1950s. Reference: Guide for Biology, Campbell, Reece and Mitchell 38 IG Lecture Outlines 1- Sanger used protein-digesting enzymes and other catalysts to hydrolyze the insulin at specific places. The fragments were then separated by a technique called chromatography. Hydrolysis by another agent broke the polypeptide at different sites, yielding a second group of fragments. Sanger used chemical methods to determine the sequence of amino acids in the small fragments. He then searched for overlapping regions among the pieces obtained by hydrolyzing with the different agents. After years of effort, Sanger was able to reconstruct the complete primary structure of insulin. Most of the steps in sequencing a polypeptide have since been automated. 3. Protein conformation determines protein function. A functional protein consists of one or more polypeptides that have been twisted, folded, and coiled into a unique shape. It is the order of amino acids that determines what the three-dimensional conformation of the protein will be. A protein’s specific conformation determines its function. When a cell synthesizes a polypeptide, the chain generally folds spontaneously to assume the functional conformation for that protein. The folding is reinforced by a variety of bonds between parts of the chain, which in turn depend on the sequence of amino acids. Many proteins are globular, while others are fibrous in shape. In almost every case, the function of a protein depends on its ability to recognize and bind to some other molecule. For example, an antibody binds to a particular foreign substance. An enzyme recognizes and binds to a specific substrate, facilitating a chemical reaction. Natural signal molecules called endorphins bind to specific receptor proteins on the surface of brain cells in humans, producing euphoria and relieving pain. Morphine, heroin, and other opiate drugs mimic endorphins because they are similar in shape and can bind to the brain’s endorphin receptors. The function of a protein is an emergent property resulting from its specific molecular order. Three levels of structure—primary, secondary, and tertiary structures—organize the folding within a single polypeptide. Quaternary structure arises when two or more polypeptides join to form a protein. The primary structure of a protein is its unique sequence of amino acids. Lysozyme, an enzyme that attacks bacteria, consists of 129 amino acids. The precise primary structure of a protein is determined by inherited genetic information. Even a slight change in primary structure can affect a protein’s conformation and ability to function. The substitution of one amino acid (valine) for the normal one (glutamic acid) at a particular position in the primary structure of hemoglobin, the protein that carries oxygen in red blood cells, can cause sickle-cell disease, an inherited blood disorder. The abnormal hemoglobins crystallize, deforming the red blood cells into a sickle shape and clogging capillaries. Most proteins have segments of their polypeptide chains repeatedly coiled or folded. Reference: Guide for Biology, Campbell, Reece and Mitchell 39 IG Lecture Outlines 1- These coils and folds are referred to as secondary structure and result from hydrogen bonds between the repeating constituents of the polypeptide backbone. The weakly positive hydrogen atom attached to the nitrogen atom has an affinity for the oxygen atom of a nearby peptide bond. Each hydrogen bond is weak, but the sum of many hydrogen bonds stabilizes the structure of part of the protein. Typical secondary structures are coils (an alpha helix) or folds (beta pleated sheets). The structural properties of silk are due to beta pleated sheets. The presence of so many hydrogen bonds makes each silk fiber stronger than a steel strand of the same weight. Tertiary structure is determined by interactions among various R groups. These interactions include hydrogen bonds between polar and/or charged areas, ionic bonds between charged R groups, and hydrophobic interactions and van der Waals interactions among hydrophobic R groups. While these three interactions are relatively weak, strong covalent bonds called disulfide bridges that form between the sulfhydryl groups (SH) of two cysteine monomers act to rivet parts of the protein together. Quaternary structure results from the aggregation of two or more polypeptide subunits. Collagen is a fibrous protein of three polypeptides that are supercoiled like a rope. This provides structural strength for collagen’s role in connective tissue. Hemoglobin is a globular protein with quaternary structure. It consists of four polypeptide subunits: two alpha and two beta chains. Both types of subunits consist primarily of alpha-helical secondary structure. Each subunit has a nonpeptide heme component with an iron atom that binds oxygen. What are the key factors determining protein conformation? A polypeptide chain of a given amino acid sequence can spontaneously arrange itself into a 3D shape determined and maintained by the interactions responsible for secondary and tertiary structure. The folding occurs as the protein is being synthesized within the cell. However, protein conformation also depends on the physical and chemical conditions of the protein’s environment. Alterations in pH, salt concentration, temperature, or other factors can unravel or denature a protein. These forces disrupt the hydrogen bonds, ionic bonds, and disulfide bridges that maintain the protein’s shape. Most proteins become denatured if the are transferred to an organic solvent. The polypeptide chain refolds so that its hydrophobic regions face outward, toward the solvent. Denaturation can also be caused by heat, which disrupts the weak interactions that stabilize conformation. This explains why extremely high fevers can be fatal. Proteins in the blood become denatured by the high body temperatures. Some proteins can return to their functional shape after denaturation, but others cannot, especially in the crowded environment of the cell. Biochemists now know the amino acid sequences of more than 875,000 proteins and the 3D shapes of about 7,000. Reference: Guide for Biology, Campbell, Reece and Mitchell 40 IG Lecture Outlines 1- Nevertheless, it is still difficult to predict the conformation of a protein from its primary structure alone. Most proteins appear to undergo several intermediate stages before reaching their “mature” configuration. The folding of many proteins is assisted by chaperonins or chaperone proteins. Chaperonins do not specify the final structure of a polypeptide but rather work to segregate and protect the polypeptide while it folds spontaneously. At present, scientists use X-ray crystallography to determine protein conformation. This technique requires the formation of a crystal of the protein being studied. The pattern of diffraction of an X-ray by the atoms of the crystal can be used to determine the location of the atoms and to build a computer model of its structure. Nuclear magnetic resonance (NMR) spectroscopy has recently been applied to this problem. This method does not require protein crystallization. E. Nucleic Acids Store and Transmit Hereditary Information The amino acid sequence of a polypeptide is programmed by a unit of inheritance known as a gene. A gene consists of DNA, a polymer known as a nucleic acid. 1. There are two types of nucleic acids: RNA and DNA. There are two types of nucleic acids: ribonucleic acid (RNA) and deoxyribonucleic acid (DNA). These are the molecules that allow living organisms to reproduce their complex components from generation to generation. DNA provides directions for its own replication. DNA also directs RNA synthesis and, through RNA, controls protein synthesis. Organisms inherit DNA from their parents. Each DNA molecule is very long, consisting of hundreds to thousands of genes. Before a cell reproduces itself by dividing, its DNA is copied. The copies are then passed to the next generation of cells. While DNA encodes the information that programs all the cell’s activities, it is not directly involved in the day-to-day operations of the cell. Proteins are responsible for implementing the instructions contained in DNA. Each gene along a DNA molecule directs the synthesis of a specific type of messenger RNA molecule (mRNA). The mRNA molecule interacts with the cell’s protein-synthesizing machinery to direct the ordering of amino acids in a polypeptide. The flow of genetic information is from DNA -> RNA -> protein. Protein synthesis occurs on cellular structures called ribosomes. In eukaryotes, DNA is located in the nucleus, but most ribosomes are in the cytoplasm. mRNA functions as an intermediary, moving information and directions from the nucleus to the cytoplasm. Prokaryotes lack nuclei but still use RNA as an intermediary to carry a message from DNA to the ribosomes. 2. A nucleic acid strand is a polymer of nucleotides. Reference: Guide for Biology, Campbell, Reece and Mitchell 41 IG Lecture Outlines 1- Nucleic acids are polymers made of nucleotide monomers. Each nucleotide consists of three parts: a nitrogenous base, a pentose sugar, and a phosphate group. The nitrogen bases are rings of carbon and nitrogen that come in two types: purines and pyrimidines. Pyrimidines have a single six-membered ring. There are three different pyrimidines: cytosine (C), thymine (T), and uracil (U). Purines have a six-membered ring joined to a five-membered ring. The two purines are adenine (A) and guanine (G). The pentose joined to the nitrogen base is ribose in nucleotides of RNA and deoxyribose in DNA. The only difference between the sugars is the lack of an oxygen atom on carbon two in deoxyribose. Because the atoms in both the nitrogenous base and the sugar are numbered, the sugar atoms have a prime after the number to distinguish them. Thus, the second carbon in the sugar ring is the 2’ (2 prime) carbon and the carbon that sticks up from the ring is the 5’ carbon. The combination of a pentose and a nitrogenous base is a nucleoside. The addition of a phosphate group creates a nucleoside monophosphate or nucleotide. Polynucleotides are synthesized when adjacent nucleotides are joined by covalent bonds called phosphodiester linkages that form between the —OH group on the 3’ of one nucleotide and the phosphate on the 5’ carbon of the next. This creates a repeating backbone of sugar-phosphate units, with appendages consisting of the nitrogenous bases. The two free ends of the polymer are distinct. One end has a phosphate attached to a 5’ carbon; this is the 5’ end. The other end has a hydroxyl group on a 3’ carbon; this is the 3’ end. The sequence of bases along a DNA or mRNA polymer is unique for each gene. Because genes are normally hundreds to thousands of nucleotides long, the number of possible base combinations is virtually limitless. The linear order of bases in a gene specifies the order of amino acids—the primary structure—of a protein, which in turn determines three-dimensional conformation and function. 3. Inheritance is based on replication of the DNA double helix. An RNA molecule is a single polynucleotide chain. DNA molecules have two polynucleotide strands that spiral around an imaginary axis to form a double helix. The double helix was first proposed as the structure of DNA in 1953 by James Watson and Francis Crick. The sugar-phosphate backbones of the two polynucleotides are on the outside of the helix. The two backbones run in opposite 5’ -> 3’ directions from each other, an arrangement referred to as antiparallel. Pairs of nitrogenous bases, one from each strand, connect the polynucleotide chains with hydrogen bonds. Most DNA molecules have thousands to millions of base pairs. Reference: Guide for Biology, Campbell, Reece and Mitchell 42 IG Lecture Outlines 1- Because of their shapes, only some bases are compatible with each other. Adenine (A) always pairs with thymine (T) and guanine (G) with cytosine (C). With these base-pairing rules, if we know the sequence of bases on one strand, we know the sequence on the opposite strand. The two strands are complementary. Prior to cell division, each of the strands serves as a template to order nucleotides into a new complementary strand. This results in two identical copies of the original double-stranded DNA molecule, which are then distributed to the daughter cells. This mechanism ensures that a full set of genetic information is transmitted whenever a cell reproduces. 4. We can use DNA and proteins as tape measures of evolution. Genes (DNA) and their products (proteins) document the hereditary background of an organism. Because DNA molecules are passed from parents to offspring, siblings have greater similarity in their DNA and protein than do unrelated individuals of the same species. This argument can be extended to develop a “molecular genealogy” to relationships between species. Two species that appear to be closely related based on fossil and molecular evidence should also be more similar in DNA and protein sequences than are more distantly related species. In fact, that is so. For example, if we compare the sequence of 146 amino acids in a hemoglobin polypeptide, we find that humans and gorillas differ in just 1 amino acid. Humans and gibbons differ in 2 amino acids. Humans and rhesus monkeys differ in 8 amino acids. More distantly related species have more differences. Humans and mice differ in 27 amino acids. Humans and frogs differ in 67 amino acids. Molecular biology can be used to assess evolutionary kinship. Reference: Guide for Biology, Campbell, Reece and Mitchell Reference: Guide for Biology, Campbell, Reece and Mitchell 43 IG Lecture Outlines 1- Chapter 6 A Tour of the Cell Lecture Outline Overview: The Importance of Cells All organisms are made of cells. Many organisms are single-celled. Even in multicellular organisms, the cell is the basic unit of structure and function. The cell is the simplest collection of matter that can live. All cells are related by their descent from earlier cells. A. How We Study Cells 1. Microscopes provide windows to the world of the cell. The discovery and early study of cells progressed with the invention of microscopes in 1590 and their improvement in the 17th century. In a light microscope (LM), visible light passes through the specimen and then through glass lenses. The lenses refract light such that the image is magnified into the eye or onto a video screen. Microscopes vary in magnification and resolving power. Magnification is the ratio of an object’s image to its real size. Resolving power is a measure of image clarity. It is the minimum distance two points can be separated and still be distinguished as two separate points. Resolution is limited by the shortest wavelength of the radiation used for imaging. The minimum resolution of a light microscope is about 200 nanometers (nm), the size of a small bacterium. Light microscopes can magnify effectively to about 1,000 times the size of the actual specimen. At higher magnifications, the image blurs. Techniques developed in the 20th century have enhanced contrast and enabled particular cell components to be stained or labeled so they stand out. While a light microscope can resolve individual cells, it cannot resolve much of the internal anatomy, especially the organelles. To resolve smaller structures, we use an electron microscope (EM), which focuses a beam of electrons through the specimen or onto its surface. Because resolution is inversely related to wavelength used, electron microscopes (whose electron beams have shorter wavelengths than visible light) have finer resolution. Theoretically, the resolution of a modern EM could reach 0.002 nanometer (nm), but the practical limit is closer to about 2 nm. Reference: Guide for Biology, Campbell, Reece and Mitchell 44 IG Lecture Outlines 1- Transmission electron microscopes (TEMs) are used mainly to study the internal ultrastructure of cells. A TEM aims an electron beam through a thin section of the specimen. The image is focused and magnified by electromagnets. To enhance contrast, the thin sections are stained with atoms of heavy metals. Scanning electron microscopes (SEMs) are useful for studying surface structures. The sample surface is covered with a thin film of gold. The beam excites electrons on the surface of the sample. These secondary electrons are collected and focused on a screen. The result is an image of the topography of the specimen. The SEM has great depth of field, resulting in an image that seems three-dimensional. Electron microscopes reveal organelles that are impossible to resolve with the light microscope. However, electron microscopes can only be used on dead cells. Light microscopes do not have as high a resolution, but they can be used to study live cells. Microscopes are major tools in cytology, the study of cell structures. Cytology combined with biochemistry, the study of molecules and chemical processes in metabolism, to produce modern cell biology. 2. Cell biologists can isolate organelles to study their functions. The goal of cell fractionation is to separate the major organelles of the cells so their individual functions can be studied. This process is driven by an ultracentrifuge, a machine that can spin at up to 130,000 revolutions per minute and apply forces of more than 1 million times gravity (1,000,000 g). Fractionation begins with homogenization, gently disrupting the cell. The homogenate is spun in a centrifuge to separate heavier pieces into the pellet while lighter particles remain in the supernatant. As the process is repeated at higher speeds and for longer durations, smaller and smaller organelles can be collected in subsequent pellets. Cell fractionation prepares isolates of specific cell components. This enables the functions of these organelles to be determined, especially by the reactions or processes catalyzed by their proteins. For example, one cellular fraction was enriched in enzymes that function in cellular respiration. Electron microscopy revealed that this fraction is rich in mitochondria. This evidence helped cell biologists determine that mitochondria are the site of cellular respiration. Cytology and biochemistry complement each other in correlating cellular structure and function. B. A Panoramic View of the Cell 1. Prokaryotic and eukaryotic cells differ in size and complexity. All cells are surrounded by a plasma membrane. The semifluid substance within the membrane is the cytosol, containing the organelles. All cells contain chromosomes that have genes in the form of DNA. Reference: Guide for Biology, Campbell, Reece and Mitchell 45 IG Lecture Outlines 1- All cells also have ribosomes, tiny organelles that make proteins using the instructions contained in genes. A major difference between prokaryotic and eukaryotic cells is the location of chromosomes. In a eukaryotic cell, chromosomes are contained in a membrane-enclosed organelle, the nucleus. In a prokaryotic cell, the DNA is concentrated in the nucleoid without a membrane separating it from the rest of the cell. In eukaryote cells, the chromosomes are contained within a membranous nuclear envelope. The region between the nucleus and the plasma membrane is the cytoplasm. All the material within the plasma membrane of a prokaryotic cell is cytoplasm. Within the cytoplasm of a eukaryotic cell are a variety of membrane-bound organelles of specialized form and function. These membrane-bound organelles are absent in prokaryotes. Eukaryotic cells are generally much bigger than prokaryotic cells. The logistics of carrying out metabolism set limits on cell size. At the lower limit, the smallest bacteria, mycoplasmas, are between 0.1 to 1.0 micron. Most bacteria are 1–10 microns in diameter. Eukaryotic cells are typically 10–100 microns in diameter. Metabolic requirements also set an upper limit to the size of a single cell. As a cell increases in size, its volume increases faster than its surface area. Smaller objects have a greater ratio of surface area to volume. The plasma membrane functions as a selective barrier that allows the passage of oxygen, nutrients, and wastes for the whole volume of the cell. The volume of cytoplasm determines the need for this exchange. Rates of chemical exchange across the plasma membrane may be inadequate to maintain a cell with a very large cytoplasm. The need for a surface sufficiently large to accommodate the volume explains the microscopic size of most cells. Larger organisms do not generally have larger cells than smaller organisms—simply more cells. Cells that exchange a lot of material with their surroundings, such as intestinal cells, may have long, thin projections from the cell surface called microvilli. Microvilli increase surface area without significantly increasing cell volume. 2. Internal membranes compartmentalize the functions of a eukaryotic cell. A eukaryotic cell has extensive and elaborate internal membranes, which partition the cell into compartments. These membranes also participate directly in metabolism, as many enzymes are built into membranes. The compartments created by membranes provide different local environments that facilitate specific metabolic functions, allowing several incompatible processes to go on simultaneously in a cell. The general structure of a biological membrane is a double layer of phospholipids. Other lipids and diverse proteins are embedded in the lipid bilayer or attached to its surface. Each type of membrane has a unique combination of lipids and proteins for its specific functions. Reference: Guide for Biology, Campbell, Reece and Mitchell 46 IG Lecture Outlines 1- For example, enzymes embedded in the membranes of mitochondria function in cellular respiration. C. The Nucleus and Ribosomes 1. The nucleus contains a eukaryotic cell’s genetic library. The nucleus contains most of the genes in a eukaryotic cell. Additional genes are located in mitochondria and chloroplasts. The nucleus averages about 5 microns in diameter. The nucleus is separated from the cytoplasm by a double membrane called the nuclear envelope. The two membranes of the nuclear envelope are separated by 20–40 nm. The envelope is perforated by pores that are about 100 nm in diameter. At the lip of each pore, the inner and outer membranes of the nuclear envelope are fused to form a continuous membrane. A protein structure called a pore complex lines each pore, regulating the passage of certain large macromolecules and particles. The nuclear side of the envelope is lined by the nuclear lamina, a network of protein filaments that maintains the shape of the nucleus. There is evidence that a framework of fibers called the nuclear matrix extends through the nuclear interior. Within the nucleus, the DNA and associated proteins are organized into discrete units called chromosomes, structures that carry the genetic information. Each chromosome is made up of fibrous material called chromatin, a complex of proteins and DNA. Stained chromatin appears through light microscopes and electron microscopes as a diffuse mass. As the cell prepares to divide, the chromatin fibers coil up and condense, becoming thick enough to be recognized as the familiar chromosomes. Each eukaryotic species has a characteristic number of chromosomes. A typical human cell has 46 chromosomes. A human sex cell (egg or sperm) has only 23 chromosomes. In the nucleus is a region of densely stained fibers and granules adjoining chromatin, the nucleolus. In the nucleolus, ribosomal RNA (rRNA) is synthesized and assembled with proteins from the cytoplasm to form ribosomal subunits. The subunits pass through the nuclear pores to the cytoplasm, where they combine to form ribosomes. The nucleus directs protein synthesis by synthesizing messenger RNA (mRNA). The mRNA travels to the cytoplasm through the nuclear pores and combines with ribosomes to translate its genetic message into the primary structure of a specific polypeptide. 2. Ribosomes build a cell’s proteins. Ribosomes, containing rRNA and protein, are the organelles that carry out protein synthesis. Cell types that synthesize large quantities of proteins (e.g., pancreas cells) have large numbers of ribosomes and prominent nucleoli. Some ribosomes, free ribosomes, are suspended in the cytosol and synthesize proteins that function within the cytosol. Reference: Guide for Biology, Campbell, Reece and Mitchell 47 IG Lecture Outlines 1- Other ribosomes, bound ribosomes, are attached to the outside of the endoplasmic reticulum or nuclear envelope. These synthesize proteins that are either included in membranes or exported from the cell. Ribosomes can shift between roles depending on the polypeptides they are synthesizing. D. The Endomembrane System Many of the internal membranes in a eukaryotic cell are part of the endomembrane system. These membranes are either directly continuous or connected via transfer of vesicles, sacs of membrane. In spite of these connections, these membranes are diverse in function and structure. The thickness, molecular composition and types of chemical reactions carried out by proteins in a given membrane may be modified several times during a membrane’s life. The endomembrane system includes the nuclear envelope, endoplasmic reticulum, Golgi apparatus, lysosomes, vacuoles, and the plasma membrane. 1. The endoplasmic reticulum manufactures membranes and performs many other biosynthetic functions. The endoplasmic reticulum (ER) accounts for half the membranes in a eukaryotic cell. The ER includes membranous tubules and internal, fluid-filled spaces called cisternae. The ER membrane is continuous with the nuclear envelope, and the cisternal space of the ER is continuous with the space between the two membranes of the nuclear envelope. There are two connected regions of ER that differ in structure and function. Smooth ER looks smooth because it lacks ribosomes. Rough ER looks rough because ribosomes (bound ribosomes) are attached to the outside, including the outside of the nuclear envelope. The smooth ER is rich in enzymes and plays a role in a variety of metabolic processes. Enzymes of smooth ER synthesize lipids, including oils, phospholipids, and steroids. These include the sex hormones of vertebrates and adrenal steroids. In the smooth ER of the liver, enzymes help detoxify poisons and drugs such as alcohol and barbiturates. Frequent use of these drugs leads to the proliferation of smooth ER in liver cells, increasing the rate of detoxification. This increases tolerance to the target and other drugs, so higher doses are required to achieve the same effect. Smooth ER stores calcium ions. Muscle cells have a specialized smooth ER that pumps calcium ions from the cytosol and stores them in its cisternal space. When a nerve impulse stimulates a muscle cell, calcium ions rush from the ER into the cytosol, triggering contraction. Enzymes then pump the calcium back, readying the cell for the next stimulation. Rough ER is especially abundant in cells that secrete proteins. As a polypeptide is synthesized on a ribosome attached to rough ER, it is threaded into the cisternal space through a pore formed by a protein complex in the ER membrane. As it enters the cisternal space, the new protein folds into its native conformation. Most secretory polypeptides are glycoproteins, proteins to which a carbohydrate is attached. Reference: Guide for Biology, Campbell, Reece and Mitchell 48 IG Lecture Outlines 1- Secretory proteins are packaged in transport vesicles that carry them to their next stage. Rough ER is also a membrane factory. Membrane-bound proteins are synthesized directly into the membrane. Enzymes in the rough ER also synthesize phospholipids from precursors in the cytosol. As the ER membrane expands, membrane can be transferred as transport vesicles to other components of the endomembrane system. 2. The Golgi apparatus is the shipping and receiving center for cell products. Many transport vesicles from the ER travel to the Golgi apparatus for modification of their contents. The Golgi is a center of manufacturing, warehousing, sorting, and shipping. The Golgi apparatus is especially extensive in cells specialized for secretion. The Golgi apparatus consists of flattened membranous sacs—cisternae—looking like a stack of pita bread. The membrane of each cisterna separates its internal space from the cytosol. One side of the Golgi, the cis side, is located near the ER. The cis face receives material by fusing with transport vesicles from the ER. The other side, the trans side, buds off vesicles that travel to other sites. During their transit from the cis to the trans side, products from the ER are usually modified. The Golgi can also manufacture its own macromolecules, including pectin and other noncellulose polysaccharides. The Golgi apparatus is a very dynamic structure. According to the cisternal maturation model, the cisternae of the Golgi progress from the cis to the trans face, carrying and modifying their protein cargo as they move. Finally, the Golgi sorts and packages materials into transport vesicles. Molecular identification tags are added to products to aid in sorting. Products are tagged with identifiers such as phosphate groups. These act like ZIP codes on mailing labels to identify the product’s final destination. 3. Lysosomes are digestive compartments. A lysosome is a membrane-bound sac of hydrolytic enzymes that an animal cell uses to digest macromolecules. Lysosomal enzymes can hydrolyze proteins, fats, polysaccharides, and nucleic acids. These enzymes work best at pH 5. Proteins in the lysosomal membrane pump hydrogen ions from the cytosol into the lumen of the lysosomes. Rupture of one or a few lysosomes has little impact on a cell because the lysosomal enzymes are not very active at the neutral pH of the cytosol. However, massive rupture of many lysosomes can destroy a cell by autodigestion. Lysosomal enzymes and membrane are synthesized by rough ER and then transferred to the Golgi apparatus for further modification. Proteins on the inner surface of the lysosomal membrane are spared by digestion by their threedimensional conformations, which protect vulnerable bonds from hydrolysis. Lysosomes carry out intracellular digestion in a variety of circumstances. Reference: Guide for Biology, Campbell, Reece and Mitchell 49 IG Lecture Outlines 1- Amoebas eat by engulfing smaller organisms by phagocytosis. The food vacuole formed by phagocytosis fuses with a lysosome, whose enzymes digest the food. As the polymers are digested, monomers pass to the cytosol to become nutrients for the cell. Lysosomes can play a role in recycling of the cell’s organelles and macromolecules. This recycling, or autophagy, renews the cell. During autophagy, a damaged organelle or region of cytosol becomes surrounded by membrane. A lysosome fuses with the resulting vesicle, digesting the macromolecules and returning the organic monomers to the cytosol for reuse. The lysosomes play a critical role in the programmed destruction of cells in multicellular organisms. This process plays an important role in development. The hands of human embryos are webbed until lysosomes digest the cells in the tissue between the fingers. This important process is called programmed cell death, or apoptosis. 4. Vacuoles have diverse functions in cell maintenance. Vesicles and vacuoles (larger versions) are membrane-bound sacs with varied functions. Food vacuoles are formed by phagocytosis and fuse with lysosomes. Contractile vacuoles, found in freshwater protists, pump excess water out of the cell to maintain the appropriate concentration of salts. A large central vacuole is found in many mature plant cells. The membrane surrounding the central vacuole, the tonoplast, is selective in its transport of solutes into the central vacuole. The functions of the central vacuole include stockpiling proteins or inorganic ions, disposing of metabolic byproducts, holding pigments, and storing defensive compounds that defend the plant against herbivores. Because of the large vacuole, the cytosol occupies only a thin layer between the plasma membrane and the tonoplast. The presence of a large vacuole increases surface area to volume ratio for the cell. E. Other Membranous Organelles 1. Mitochondria and chloroplasts are the main energy transformers of cells. Mitochondria and chloroplasts are the organelles that convert energy to forms that cells can use for work. Mitochondria are the sites of cellular respiration, generating ATP from the catabolism of sugars, fats, and other fuels in the presence of oxygen. Chloroplasts, found in plants and algae, are the sites of photosynthesis. They convert solar energy to chemical energy and synthesize new organic compounds such as sugars from CO2 and H2O. Mitochondria and chloroplasts are not part of the endomembrane system. In contrast to organelles of the endomembrane system, each mitochondrion or chloroplast has two membranes separating the innermost space from the cytosol. Their membrane proteins are not made by the ER, but rather by free ribosomes in the cytosol and by ribosomes within the organelles themselves. Both organelles have small quantities of DNA that direct the synthesis of the polypeptides produced by these internal ribosomes. Reference: Guide for Biology, Campbell, Reece and Mitchell 50 IG Lecture Outlines 1- Mitochondria and chloroplasts grow and reproduce as semiautonomous organelles. Almost all eukaryotic cells have mitochondria. There may be one very large mitochondrion or hundreds to thousands of individual mitochondria. The number of mitochondria is correlated with aerobic metabolic activity. A typical mitochondrion is 1–10 microns long. Mitochondria are quite dynamic: moving, changing shape, and dividing. Mitochondria have a smooth outer membrane and a convoluted inner membrane with infoldings called cristae. The inner membrane divides the mitochondrion into two internal compartments. The first is the intermembrane space, a narrow region between the inner and outer membranes. The inner membrane encloses the mitochondrial matrix, a fluid-filled space with DNA, ribosomes, and enzymes. Some of the metabolic steps of cellular respiration are catalyzed by enzymes in the matrix. The cristae present a large surface area for the enzymes that synthesize ATP. The chloroplast is one of several members of a generalized class of plant structures called plastids. Amyloplasts are colorless plastids that store starch in roots and tubers. Chromoplasts store pigments for fruits and flowers. Chloroplasts contain the green pigment chlorophyll as well as enzymes and other molecules that function in the photosynthetic production of sugar. Chloroplasts measure about 2 microns × 5 microns and are found in leaves and other green organs of plants and algae. The contents of the chloroplast are separated from the cytosol by an envelope consisting of two membranes separated by a narrow intermembrane space. Inside the innermost membrane is a fluid-filled space, the stroma, in which float membranous sacs, the thylakoids. The stroma contains DNA, ribosomes, and enzymes. The thylakoids are flattened sacs that play a critical role in converting light to chemical energy. In some regions, thylakoids are stacked like poker chips into grana. The membranes of the chloroplast divide the chloroplast into three compartments: the intermembrane space, the stroma, and the thylakoid space. Like mitochondria, chloroplasts are dynamic structures. Their shape is plastic, and they can reproduce themselves by pinching in two. Mitochondria and chloroplasts are mobile and move around the cell along tracks of the cytoskeleton. 2. Peroxisomes generate and degrade H2O2 in performing various metabolic functions. Peroxisomes contain enzymes that transfer hydrogen from various substrates to oxygen. An intermediate product of this process is hydrogen peroxide (H2O2), a poison. The peroxisome contains an enzyme that converts H2O2 to water. Some peroxisomes break fatty acids down to smaller molecules that are transported to mitochondria as fuel for cellular respiration. Peroxisomes in the liver detoxify alcohol and other harmful compounds. Specialized peroxisomes, glyoxysomes, convert the fatty acids in seeds to sugars, which the seedling can use as a source of energy and carbon until it is capable of photosynthesis. Peroxisomes are bound by a single membrane. Reference: Guide for Biology, Campbell, Reece and Mitchell 51 IG Lecture Outlines 1- They form not from the endomembrane system, but by incorporation of proteins and lipids from the cytosol. They split in two when they reach a certain size. F. The Cytoskeleton The cytoskeleton is a network of fibers extending throughout the cytoplasm. The cytoskeleton organizes the structures and activities of the cell. 1. The cytoskeleton provides support, motility, and regulation. The cytoskeleton provides mechanical support and maintains cell shape. The cytoskeleton provides anchorage for many organelles and cytosolic enzymes. The cytoskeleton is dynamic and can be dismantled in one part and reassembled in another to change the shape of the cell. The cytoskeleton also plays a major role in cell motility, including changes in cell location and limited movements of parts of the cell. The cytoskeleton interacts with motor proteins to produce motility. Cytoskeleton elements and motor proteins work together with plasma membrane molecules to move the whole cell along fibers outside the cell. Motor proteins bring about movements of cilia and flagella by gripping cytoskeletal components such as microtubules and moving them past each other. The same mechanism causes muscle cells to contract. Inside the cell, vesicles can travel along “monorails” provided by the cytoskeleton. The cytoskeleton manipulates the plasma membrane to form food vacuoles during phagocytosis. Cytoplasmic streaming in plant cells is caused by the cytoskeleton. Recently, evidence suggests that the cytoskeleton may play a role in the regulation of biochemical activities in the cell. There are three main types of fibers making up the cytoskeleton: microtubules, microfilaments, and intermediate filaments. Microtubules, the thickest fibers, are hollow rods about 25 microns in diameter and 200 nm to 25 microns in length. Microtubule fibers are constructed of the globular protein tubulin. Each tubulin molecule is a dimer consisting of two subunits. A microtubule changes in length by adding or removing tubulin dimers. Microtubules shape and support the cell and serve as tracks to guide motor proteins carrying organelles to their destination. Microtubules are also responsible for the separation of chromosomes during cell division. In many cells, microtubules grow out from a centrosome near the nucleus. These microtubules resist compression to the cell. In animal cells, the centrosome has a pair of centrioles, each with nine triplets of microtubules arranged in a ring. Before a cell divides, the centrioles replicate. A specialized arrangement of microtubules is responsible for the beating of cilia and flagella. Reference: Guide for Biology, Campbell, Reece and Mitchell 52 IG Lecture Outlines 1- Many unicellular eukaryotic organisms are propelled through water by cilia and flagella. Cilia or flagella can extend from cells within a tissue layer, beating to move fluid over the surface of the tissue. For example, cilia lining the windpipe sweep mucus carrying trapped debris out of the lungs. Cilia usually occur in large numbers on the cell surface. They are about 0.25 microns in diameter and 2–20 microns long. There are usually just one or a few flagella per cell. Flagella are the same width as cilia, but 10–200 microns long. Cilia and flagella differ in their beating patterns. A flagellum has an undulatory movement that generates force in the same direction as the flagellum’s axis. Cilia move more like oars with alternating power and recovery strokes that generate force perpendicular to the cilium’s axis. In spite of their differences, both cilia and flagella have the same ultrastructure. Both have a core of microtubules sheathed by the plasma membrane. Nine doublets of microtubules are arranged in a ring around a pair at the center. This “9 + 2” pattern is found in nearly all eukaryotic cilia and flagella. Flexible “wheels” of proteins connect outer doublets to each other and to the two central microtubules. The outer doublets are also connected by motor proteins. The cilium or flagellum is anchored in the cell by a basal body, whose structure is identical to a centriole. The bending of cilia and flagella is driven by the arms of a motor protein, dynein. Addition and removal of a phosphate group causes conformation changes in dynein. Dynein arms alternately grab, move, and release the outer microtubules. Protein cross-links limit sliding. As a result, the forces exerted by the dynein arms cause the doublets to curve, bending the cilium or flagellum. Microfilaments are solid rods about 7 nm in diameter. Each microfilament is built as a twisted double chain of actin subunits. Microfilaments can form structural networks due to their ability to branch. The structural role of microfilaments in the cytoskeleton is to bear tension, resisting pulling forces within the cell. They form a three-dimensional network just inside the plasma membrane to help support the cell’s shape, giving the cell cortex the semisolid consistency of a gel. Microfilaments are important in cell motility, especially as part of the contractile apparatus of muscle cells. In muscle cells, thousands of actin filaments are arranged parallel to one another. Thicker filaments composed of myosin interdigitate with the thinner actin fibers. Myosin molecules act as motor proteins, walking along the actin filaments to shorten the cell. In other cells, actin-myosin aggregates are less organized but still cause localized contraction. A contracting belt of microfilaments divides the cytoplasm of animal cells during cell division. Localized contraction brought about by actin and myosin also drives amoeboid movement. Reference: Guide for Biology, Campbell, Reece and Mitchell 53 IG Lecture Outlines 1- Pseudopodia, cellular extensions, extend and contract through the reversible assembly and contraction of actin subunits into microfilaments. Microfilaments assemble into networks that convert sol to gel. According to a widely accepted model, filaments near the cell’s trailing edge interact with myosin, causing contraction. The contraction forces the interior fluid into the pseudopodium, where the actin network has been weakened. The pseudopodium extends until the actin reassembles into a network. In plant cells, actin-myosin interactions and sol-gel transformations drive cytoplasmic streaming. This creates a circular flow of cytoplasm in the cell, speeding the distribution of materials within the cell. Intermediate filaments range in diameter from 8–12 nanometers, larger than microfilaments but smaller than microtubules. Intermediate filaments are a diverse class of cytoskeletal units, built from a family of proteins called keratins. Intermediate filaments are specialized for bearing tension. Intermediate filaments are more permanent fixtures of the cytoskeleton than are the other two classes. They reinforce cell shape and fix organelle location. G. Cell Surfaces and Junctions 1. Plant cells are encased by cell walls. The cell wall, found in prokaryotes, fungi, and some protists, has multiple functions. In plants, the cell wall protects the cell, maintains its shape, and prevents excessive uptake of water. It also supports the plant against the force of gravity. The thickness and chemical composition of cell walls differs from species to species and among cell types within a plant. The basic design consists of microfibrils of cellulose embedded in a matrix of proteins and other polysaccharides. This is the basic design of steel-reinforced concrete or fiberglass. A mature cell wall consists of a primary cell wall, a middle lamella with sticky polysaccharides that holds cells together, and layers of secondary cell wall. Plant cell walls are perforated by channels between adjacent cells called plasmodesmata. 2. The extracellular matrix (ECM) of animal cells functions in support, adhesion, movement, and regulation. Though lacking cell walls, animal cells do have an elaborate extracellular matrix (ECM). The primary constituents of the extracellular matrix are glycoproteins, especially collagen fibers, embedded in a network of glycoprotein proteoglycans. In many cells, fibronectins in the ECM connect to integrins, intrinsic membrane proteins that span the membrane and bind on their cytoplasmic side to proteins attached to microfilaments of the cytoskeleton. The interconnections from the ECM to the cytoskeleton via the fibronectin-integrin link permit the integration of changes inside and outside the cell. The ECM can regulate cell behavior. Reference: Guide for Biology, Campbell, Reece and Mitchell 54 IG Lecture Outlines 1- Embryonic cells migrate along specific pathways by matching the orientation of their microfilaments to the “grain” of fibers in the extracellular matrix. The extracellular matrix can influence the activity of genes in the nucleus via a combination of chemical and mechanical signaling pathways. This may coordinate the behavior of all the cells within a tissue. 3. Intercellular junctions help integrate cells into higher levels of structure and function. <BL1>Neighboring cells in tissues, organs, or organ systems often adhere, interact, and communicate through direct physical contact. Plant cells are perforated with plasmodesmata, channels allowing cytosol to pass between cells. Water and small solutes can pass freely from cell to cell. In certain circumstances, proteins and RNA can be exchanged. Animals have 3 main types of intercellular links: tight junctions, desmosomes, and gap junctions. In tight junctions, membranes of adjacent cells are fused, forming continuous belts around cells. This prevents leakage of extracellular fluid. Desmosomes (or anchoring junctions) fasten cells together into strong sheets, much like rivets. Intermediate filaments of keratin reinforce desmosomes. Gap junctions (or communicating junctions) provide cytoplasmic channels between adjacent cells. Special membrane proteins surround these pores. Ions, sugars, amino acids, and other small molecules can pass. In embryos, gap junctions facilitate chemical communication during development. 4. A cell is a living unit greater than the sum of its parts. While the cell has many structures with specific functions, all these structures must work together. For example, macrophages use actin filaments to move and extend pseudopodia to capture their bacterial prey. Food vacuoles are digested by lysosomes, a product of the endomembrane system of ER and Golgi. The enzymes of the lysosomes and proteins of the cytoskeleton are synthesized on the ribosomes. The information for the proteins comes from genetic messages sent by DNA in the nucleus. All of these processes require energy in the form of ATP, most of which is supplied by the mitochondria. A cell is a living unit greater than the sum of its parts. Reference: Guide for Biology, Campbell, Reece and Mitchell Reference: Guide for Biology, Campbell, Reece and Mitchell 55 IG Lecture Outlines 1- Chapter 7 Membrane Structure and Function Lecture Outline Overview The plasma membrane separates the living cell from its nonliving surroundings. This thin barrier, 8 nm thick, controls traffic into and out of the cell. Like all biological membranes, the plasma membrane is selectively permeable, allowing some substances to cross more easily than others. A. Membrane Structure The main macromolecules in membranes are lipids and proteins, but carbohydrates are also important. The most abundant lipids are phospholipids. Phospholipids and most other membrane constituents are amphipathic molecules. Amphipathic molecules have both hydrophobic regions and hydrophilic regions. The arrangement of phospholipids and proteins in biological membranes is described by the fluid mosaic model. 1. Membrane models have evolved to fit new data. Models of membranes were developed long before membranes were first seen with electron microscopes in the 1950s. In 1915, membranes isolated from red blood cells were chemically analyzed and found to be composed of lipids and proteins. In 1925, E. Gorter and F. Grendel reasoned that cell membranes must be a phospholipid bilayer two molecules thick. The molecules in the bilayer are arranged such that the hydrophobic fatty acid tails are sheltered from water while the hydrophilic phosphate groups interact with water. Actual membranes adhere more strongly to water than do artificial membranes composed only of phospholipids. One suggestion was that proteins on the surface of the membrane increased adhesion. In 1935, H. Davson and J. Danielli proposed a sandwich model in which the phospholipid bilayer lies between two layers of globular proteins. Early images from electron microscopes seemed to support the Davson-Danielli model, and until the 1960s, it was widely accepted as the structure of the plasma membrane and internal membranes. Further investigation revealed two problems. First, not all membranes were alike. Membranes differ in thickness, appearance when stained, and percentage of proteins. Reference: Guide for Biology, Campbell, Reece and Mitchell 56 IG Lecture Outlines 1- Membranes with different functions differ in chemical composition and structure. Second, measurements showed that membrane proteins are not very soluble in water. Membrane proteins are amphipathic, with hydrophobic and hydrophilic regions. If membrane proteins were at the membrane surface, their hydrophobic regions would be in contact with water. In 1972, S. J. Singer and G. Nicolson presented a revised model that proposed that the membrane proteins are dispersed and individually inserted into the phospholipid bilayer. In this fluid mosaic model, the hydrophilic regions of proteins and phospholipids are in maximum contact with water, and the hydrophobic regions are in a nonaqueous environment within the membrane. A specialized preparation technique, freeze-fracture, splits a membrane along the middle of the phospholipid bilayer. When a freeze-fracture preparation is viewed with an electron microscope, protein particles are interspersed in a smooth matrix, supporting the fluid mosaic model. 2. Membranes are fluid. Membrane molecules are held in place by relatively weak hydrophobic interactions. Most of the lipids and some proteins drift laterally in the plane of the membrane, but rarely flip-flop from one phospholipid layer to the other. The lateral movements of phospholipids are rapid, about 2 microns per second. A phospholipid can travel the length of a typical bacterial cell in 1 second. Many larger membrane proteins drift within the phospholipid bilayer, although they move more slowly than the phospholipids. Some proteins move in a very directed manner, perhaps guided or driven by motor proteins attached to the cytoskeleton. Other proteins never move and are anchored to the cytoskeleton. Membrane fluidity is influenced by temperature. As temperatures cool, membranes switch from a fluid state to a solid state as the phospholipids pack more closely. Membrane fluidity is also influenced by its components. Membranes rich in unsaturated fatty acids are more fluid that those dominated by saturated fatty acids because the kinks in the unsaturated fatty acid tails at the locations of the double bonds prevent tight packing. The steroid cholesterol is wedged between phospholipid molecules in the plasma membrane of animal cells. At warm temperatures (such as 37°C), cholesterol restrains the movement of phospholipids and reduces fluidity. At cool temperatures, it maintains fluidity by preventing tight packing. Thus, cholesterol acts as a “temperature buffer” for the membrane, resisting changes in membrane fluidity as temperature changes. To work properly with active enzymes and appropriate permeability, membranes must be about as fluid as salad oil. Cells can alter the lipid composition of membranes to compensate for changes in fluidity caused by changing temperatures. For example, cold-adapted organisms such as winter wheat increase the percentage of unsaturated phospholipids in their membranes in the autumn. Reference: Guide for Biology, Campbell, Reece and Mitchell 57 IG Lecture Outlines 1- This prevents membranes from solidifying during winter. 3. Membranes are mosaics of structure and function. A membrane is a collage of different proteins embedded in the fluid matrix of the lipid bilayer. Proteins determine most of the membrane’s specific functions. The plasma membrane and the membranes of the various organelles each have unique collections of proteins. There are two major populations of membrane proteins. Peripheral proteins are not embedded in the lipid bilayer at all. Instead, they are loosely bound to the surface of the protein, often connected to integral proteins. Integral proteins penetrate the hydrophobic core of the lipid bilayer, often completely spanning the membrane (as transmembrane proteins). The hydrophobic regions embedded in the membrane’s core consist of stretches of nonpolar amino acids, often coiled into alpha helices. Where integral proteins are in contact with the aqueous environment, they have hydrophilic regions of amino acids. On the cytoplasmic side of the membrane, some membrane proteins connect to the cytoskeleton. On the exterior side of the membrane, some membrane proteins attach to the fibers of the extracellular matrix. The proteins of the plasma membrane have six major functions: 4. Transport of specific solutes into or out of cells. 5. Enzymatic activity, sometimes catalyzing one of a number of steps of a metabolic pathway. 6. Signal transduction, relaying hormonal messages to the cell. 7. Cell-cell recognition, allowing other proteins to attach two adjacent cells together. 8. Intercellular joining of adjacent cells with gap or tight junctions. 9. Attachment to the cytoskeleton and extracellular matrix, maintaining cell shape and stabilizing the location of certain membrane proteins. 4. Membrane carbohydrates are important for cell-cell recognition. The plasma membrane plays the key role in cell-cell recognition. Cell-cell recognition, the ability of a cell to distinguish one type of neighboring cell from another, is crucial to the functioning of an organism. This attribute is important in the sorting and organization of cells into tissues and organs during development. It is also the basis for rejection of foreign cells by the immune system. Cells recognize other cells by binding to surface molecules, often carbohydrates, on the plasma membrane. Membrane carbohydrates are usually branched oligosaccharides with fewer than 15 sugar units. They may be covalently bonded to lipids, forming glycolipids, or more commonly to proteins, forming glycoproteins. The oligosaccharides on the external side of the plasma membrane vary from species to species, from individual to individual, and even from cell type to cell type within the same individual. This variation distinguishes each cell type. Reference: Guide for Biology, Campbell, Reece and Mitchell 58 IG Lecture Outlines 1- The four human blood groups (A, B, AB, and O) differ in the external carbohydrates on red blood cells. 5. Membranes have distinctive inside and outside faces. Membranes have distinct inside and outside faces. The two layers may differ in lipid composition. Each protein in the membrane has a directional orientation in the membrane. The asymmetrical orientation of proteins, lipids and associated carbohydrates begins during the synthesis of membrane in the ER and Golgi apparatus. Membrane lipids and proteins are synthesized in the endoplasmic reticulum. Carbohydrates are added to proteins in the ER, and the resulting glycoproteins are further modified in the Golgi apparatus. Glycolipids are also produced in the Golgi apparatus. When a vesicle fuses with the plasma membrane, the outside layer of the vesicle becomes continuous with the inside layer of the plasma membrane. In that way, molecules that originate on the inside face of the ER end up on the outside face of the plasma membrane. B. Traffic across Membranes 1. A membrane’s molecular organization results in selective permeability. A steady traffic of small molecules and ions moves across the plasma membrane in both directions. For example, sugars, amino acids, and other nutrients enter a muscle cell, and metabolic waste products leave. The cell absorbs oxygen and expels carbon dioxide. It also regulates concentrations of inorganic ions, such as Na+, K+, Ca2+, and Cl−, by shuttling them across the membrane. However, substances do not move across the barrier indiscriminately; membranes are selectively permeable. The plasma membrane allows the cell to take up many varieties of small molecules and ions and exclude others. Substances that move through the membrane do so at different rates. Movement of a molecule through a membrane depends on the interaction of the molecule with the hydrophobic core of the membrane. Hydrophobic molecules, such as hydrocarbons, CO2, and O2, can dissolve in the lipid bilayer and cross easily. The hydrophobic core of the membrane impedes the direct passage of ions and polar molecules, which cross the membrane with difficulty. This includes small molecules, such as water, and larger molecules, such as glucose and other sugars. An ion, whether a charged atom or molecule, and its surrounding shell of water also has difficulty penetrating the hydrophobic core. Proteins assist and regulate the transport of ions and polar molecules. Specific ions and polar molecules can cross the lipid bilayer by passing through transport proteins that span the membrane. Some transport proteins, called channel proteins, have a hydrophilic channel that certain molecules or ions can use as a tunnel through the membrane. For example, the passage of water through the membrane can be greatly facilitated by channel proteins known as aquaporins. Other transport proteins, called carrier proteins, bind to molecules and change shape to shuttle them across the membrane. Reference: Guide for Biology, Campbell, Reece and Mitchell 59 IG Lecture Outlines 1- Each transport protein is specific as to the substances that it will translocate. For example, the glucose transport protein in the liver will carry glucose into the cell but will not transport fructose, its structural isomer. 2. Passive transport is diffusion across a membrane with no energy expenditure. Diffusion is the tendency of molecules of any substance to spread out in the available space. Diffusion is driven by the intrinsic kinetic energy (thermal motion or heat) of molecules. Movements of individual molecules are random. However, movement of a population of molecules may be directional. Imagine a permeable membrane separating a solution with dye molecules from pure water. If the membrane has microscopic pores that are large enough, dye molecules will cross the barrier randomly. The net movement of dye molecules across the membrane will continue until both sides have equal concentrations of the dye. At this dynamic equilibrium, as many molecules cross one way as cross in the other direction. In the absence of other forces, a substance will diffuse from where it is more concentrated to where it is less concentrated, down its concentration gradient. No work must be done to move substances down the concentration gradient. Diffusion is a spontaneous process that decreases free energy and increases entropy by creating a randomized mixture. Each substance diffuses down its own concentration gradient, independent of the concentration gradients of other substances. The diffusion of a substance across a biological membrane is passive transport because it requires no energy from the cell to make it happen. The concentration gradient itself represents potential energy and drives diffusion. Because membranes are selectively permeable, the interactions of the molecules with the membrane play a role in the diffusion rate. Diffusion of molecules of limited permeability through the lipid bilayer may be assisted by transport proteins. 3. Osmosis is the passive transport of water. Differences in the relative concentration of dissolved materials in two solutions can lead to the movement of ions from one to the other. The solution with the higher concentration of solutes is hypertonic relative to the other solution. The solution with the lower concentration of solutes is hypotonic relative to the other solution. These are comparative terms. Tap water is hypertonic compared to distilled water but hypotonic compared to seawater. Solutions with equal solute concentrations are isotonic. Imagine that two sugar solutions differing in concentration are separated by a membrane that will allow water through, but not sugar. The hypertonic solution has a lower water concentration than the hypotonic solution. More of the water molecules in the hypertonic solution are bound up in hydration shells around the sugar molecules, leaving fewer unbound water molecules. Reference: Guide for Biology, Campbell, Reece and Mitchell 60 IG Lecture Outlines 1- Unbound water molecules will move from the hypotonic solution, where they are abundant, to the hypertonic solution, where they are rarer. Net movement of water continues until the solutions are isotonic. The diffusion of water across a selectively permeable membrane is called osmosis. The direction of osmosis is determined only by a difference in total solute concentration. The kinds of solutes in the solutions do not matter. This makes sense because the total solute concentration is an indicator of the abundance of bound water molecules (and, therefore, of free water molecules). When two solutions are isotonic, water molecules move at equal rates from one to the other, with no net osmosis. The movement of water by osmosis is crucial to living organisms. 4. Cell survival depends on balancing water uptake and loss. An animal cell (or other cell without a cell wall) immersed in an isotonic environment experiences no net movement of water across its plasma membrane. Water molecules move across the membrane but at the same rate in both directions. The volume of the cell is stable. The same cell in a hypertonic environment will lose water, shrivel, and probably die. A cell in a hypotonic solution will gain water, swell, and burst. For organisms living in an isotonic environment (for example, many marine invertebrates), osmosis is not a problem. The cells of most land animals are bathed in extracellular fluid that is isotonic to the cells. Organisms without rigid walls have osmotic problems in either a hypertonic or hypotonic environment and must have adaptations for osmoregulation, the control of water balance, to maintain their internal environment. For example, Paramecium, a protist, is hypertonic to the pond water in which it lives. In spite of a cell membrane that is less permeable to water than other cells, water still continually enters the Paramecium cell. To solve this problem, Paramecium cells have a specialized organelle, the contractile vacuole, which functions as a bilge pump to force water out of the cell. The cells of plants, prokaryotes, fungi, and some protists have walls that contribute to the cell’s water balance. A plant cell in a hypotonic solution will swell until the elastic cell wall opposes further uptake. At this point the cell is turgid (very firm), a healthy state for most plant cells. Turgid cells contribute to the mechanical support of the plant. If a plant cell and its surroundings are isotonic, there is no movement of water into the cell. The cell becomes flaccid (limp), and the plant may wilt. The cell wall provides no advantages when a plant cell is immersed in a hypertonic solution. As the plant cell loses water, its volume shrinks. Eventually, the plasma membrane pulls away from the wall. This plasmolysis is usually lethal. 5. Specific proteins facilitate passive transport of water and selected solutes. Many polar molecules and ions that are normally impeded by the lipid bilayer of the membrane diffuse passively with the help of transport proteins that span the membrane. Reference: Guide for Biology, Campbell, Reece and Mitchell 61 IG Lecture Outlines 1- The passive movement of molecules down their concentration gradient via transport proteins is called facilitated diffusion. Two types of transport proteins facilitate the movement of molecules or ions across membranes: channel proteins and carrier proteins. Some channel proteins simply provide hydrophilic corridors for the passage of specific molecules or ions. For example, water channel proteins, aquaporins, greatly facilitate the diffusion of water. Many ion channels function as gated channels. These channels open or close depending on the presence or absence of a chemical or physical stimulus. If chemical, the stimulus is a substance other than the one to be transported. For example, stimulation of a receiving neuron by specific neurotransmitters opens gated channels to allow sodium ions into the cell. When the neurotransmitters are not present, the channels are closed. Some transport proteins do not provide channels but appear to actually translocate the solute-binding site and solute across the membrane as the transport protein changes shape. These shape changes may be triggered by the binding and release of the transported molecule. In certain inherited diseases, specific transport systems may be defective or absent. Cystinuria is a human disease characterized by the absence of a protein that transports cysteine and other amino acids across the membranes of kidney cells. An individual with cystinuria develops painful kidney stones as amino acids accumulate and crystallize in the kidneys. 6. Active transport uses energy to move solutes against their gradients. Some transport proteins can move solutes across membranes against their concentration gradient, from the side where they are less concentrated to the side where they are more concentrated. This active transport requires the cell to expend metabolic energy. Active transport enables a cell to maintain its internal concentrations of small molecules that would otherwise diffuse across the membrane. Active transport is performed by specific proteins embedded in the membranes. ATP supplies the energy for most active transport. ATP can power active transport by transferring a phosphate group from ATP (forming ADP) to the transport protein. This may induce a conformational change in the transport protein, translocating the solute across the membrane. The sodium-potassium pump actively maintains the gradient of sodium ions (Na+) and potassium ions (K+) across the plasma membrane of animal cells. Typically, K+ concentration is low outside an animal cell and high inside the cell, while Na+ concentration is high outside an animal cell and low inside the cell. The sodium-potassium pump maintains these concentration gradients, using the energy of one ATP to pump three Na+ out and two K+ in. 7. Some ion pumps generate voltage across membranes. All cells maintain a voltage across their plasma membranes. Voltage is electrical potential energy due to the separation of opposite charges. Reference: Guide for Biology, Campbell, Reece and Mitchell 62 IG Lecture Outlines 1- The cytoplasm of a cell is negative in charge compared to the extracellular fluid because of an unequal distribution of cations and anions on opposite sides of the membrane. The voltage across a membrane is called a membrane potential, and ranges from −50 to −200 millivolts (mV). The inside of the cell is negative compared to the outside. The membrane potential acts like a battery. The membrane potential favors the passive transport of cations into the cell and anions out of the cell. Two combined forces, collectively called the electrochemical gradient, drive the diffusion of ions across a membrane. One is a chemical force based on an ion’s concentration gradient. The other is ann electrical force based on the effect of the membrane potential on the ion’s movement. An ion does not simply diffuse down its concentration gradient but diffuses down its electrochemical gradient. For example, there is a higher concentration of Na+ outside a resting nerve cell than inside. When the neuron is stimulated, a gated channel opens and Na+ diffuse into the cell down their electrochemical gradient. The diffusion of Na+ is driven by their concentration gradient and by the attraction of cations to the negative side of the membrane. Special transport proteins, electrogenic pumps, generate the voltage gradient across a membrane. The sodium-potassium pump in animals restores the electrochemical gradient not only by the active transport of Na+ and K+, setting up a concentration gradient, but because it pumps two K+ inside for every three Na+ that it moves out, setting up a voltage across the membrane. The sodium-potassium pump is the major electrogenic pump of animal cells. In plants, bacteria, and fungi, a proton pump is the major electrogenic pump, actively transporting H+ out of the cell. Proton pumps in the cristae of mitochondria and the thylakoids of chloroplasts concentrate H+ behind membranes. These electrogenic pumps store energy that can be accessed for cellular work. 8. In cotransport, a membrane protein couples the transport of two solutes. A single ATP-powered pump that transports one solute can indirectly drive the active transport of several other solutes in a mechanism called cotransport. As the solute that has been actively transported diffuses back passively through a transport protein, its movement can be coupled with the active transport of another substance against its concentration gradient. Plants commonly use the gradient of hydrogen ions generated by proton pumps to drive the active transport of amino acids, sugars, and other nutrients into the cell. One specific transport protein couples the diffusion of protons out of the cell and the transport of sucrose into the cell. Plants use the mechanism of sucrose-proton cotransport to load sucrose into specialized cells in the veins of leaves for distribution to nonphotosynthetic organs such as roots. 9. Exocytosis and endocytosis transport large molecules across membranes. Small molecules and water enter or leave the cell through the lipid bilayer or by transport proteins. Large molecules, such as polysaccharides and proteins, cross the membrane via vesicles. During exocytosis, a transport vesicle budded from the Golgi apparatus is moved by the cytoskeleton to the plasma membrane. Reference: Guide for Biology, Campbell, Reece and Mitchell 63 IG Lecture Outlines 1- When the two membranes come in contact, the bilayers fuse and spill the contents to the outside. Many secretory cells use exocytosis to export their products. During endocytosis, a cell brings in macromolecules and particulate matter by forming new vesicles from the plasma membrane. Endocytosis is a reversal of exocytosis, although different proteins are involved in the two processes. A small area of the plasma membrane sinks inward to form a pocket. As the pocket deepens, it pinches in to form a vesicle containing the material that had been outside the cell. There are three types of endocytosis: phagocytosis (“cellular eating”), pinocytosis (“cellular drinking”), and receptor-mediated endocytosis. In phagocytosis, the cell engulfs a particle by extending pseudopodia around it and packaging it in a large vacuole. The contents of the vacuole are digested when the vacuole fuses with a lysosome. In pinocytosis, a cell creates a vesicle around a droplet of extracellular fluid. All included solutes are taken into the cell in this nonspecific process. Receptor-mediated endocytosis allows greater specificity, transporting only certain substances. This process is triggered when extracellular substances, or ligands, bind to special receptors on the membrane surface. The receptor proteins are clustered in regions of the membrane called coated pits, which are lined on their cytoplasmic side by a layer of coat proteins. Binding of ligands to receptors triggers the formation of a vesicle by the coated pit, bringing the bound substances into the cell. Receptor-mediated endocytosis enables a cell to acquire bulk quantities of specific materials that may be in low concentrations in the environment. Human cells use this process to take in cholesterol for use in the synthesis of membranes and as a precursor for the synthesis of steroids. Cholesterol travels in the blood in low-density lipoproteins (LDL), complexes of protein and lipid. These lipoproteins act as ligands to bind to LDL receptors and enter the cell by endocytosis. In an inherited disease called familial hypercholesterolemia, the LDL receptors are defective, leading to an accumulation of LDL and cholesterol in the blood. This contributes to early atherosclerosis. Reference: Guide for Biology, Campbell, Reece and Mitchell Reference: Guide for Biology, Campbell, Reece and Mitchell 64 IG Lecture Outlines 1-