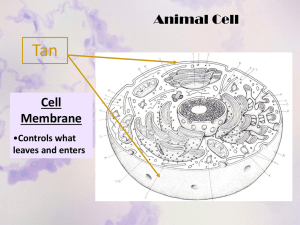
113_supp1 - CMGM Stanford

4/15/20
Supplemental Material for “Dissecting Eukaryotic Translation and Its
Control by Ribosome Density Mapping”
This file includes:
Supplemental Methods.
Supplemental Results.
Supplemental Figure legends 1 and 2. Note that Supplemental Figures 1 and 2 are on a separate file because of their size.
Supplemental Figures 3 and 4.
Supplemental Methods
A. Comparing the predicted densities for mRNAs of different lengths with the experimentally measured values
Previously we observed by global analysis that longer ORFs have lower overall ribosomal density ( Arava et al.
2003). We proposed three models to explain this inverse correlation: (1) mRNAs, regardless of length, have a certain level of incomplete elongation processivity. Longer mRNAs would then have lower overall ribosomal density than shorter mRNAs because they have more elongation steps, giving the ribosomes more chances to dissociate prior to completing translation. (2) mRNAs, regardless of length, have slow termination relative to elongation. Shorter mRNAs would then have higher ribosomal density than longer mRNA because the incremental effect on overall density from queuing of the additional ribosomes at the termination site is higher for short mRNAs compared to long mRNAs. (3) Longer mRNAs have slower initiation rates, with similar elongation and termination rates for all mRNAs, thereby leading to overall lower ribosomal density.
These three models predict different local densities of ribosomes along mRNAs
(Supplemental Figure 3): (1) With imperfect elongation there will be a gradual decrease in ribosome density towards the 3'-end of the mRNA (red line). The extent of decrease in ribosomal density (i.e., the steepness of the red line in Figure 3) will be directly related to the processivity level; lower processivity leads to steeper decrease and higher processivity will lead to a shallower decrease. (2) With termination slow relative to elongation,
1
4/15/20 ribosomes are queuing at maximal density upstream to the stop codon (green line). The length of the termination queue is dependent on how slow termination is compared to elongation. (3) With slower initiation for increasing ORF length but fast termination and highly processive elongation, the local ribosomal density is constant along the mRNA
(black dashed line).
Although the local density is different in each of these models, our previous analysis (1) was limited to identification of the overall (or average) ribosome density of the mRNA. The following section describes the derivation of the overall ribosomal density from the local density according to each of these models. This allows comparison between the models and the results from the global analysis of ribosomal density. Best fits to the density values obtained by the global analysis are presented in Figure 7 of the main text.
1. Ribosome density along mRNAs according to the incomplete processivity model
Total ORF length (nts)
Initial (5'-most) ribosome density (ribosomes/100 nts)
Probability of completing an elongation step
Ribosome density at position x (nts) d c r cr x/3
(RD) overall
= Ribosome density (ribosomes/ 100 nts) for ORF with length d
=
0 d cr x / 3 dx d
=
3 c ( r d / 3 r
0
) d ln r
= c for 0 < r < 1 for r = 1 (S1)
Equation S1 was used to calculate the expected ribosome density values for mRNA with different ORF lengths and various processivity levels. The best fit to the experimental data (minimal root mean square) is found with a processivity level of 99.3% per elongation step and an initial ribosome density ( c ) of 1.5 ribosomes/ 100 nts (Figure
7A, solid line).
2. Ribosome density along mRNAs according to the slow termination model
Total ORF length (nts) d
Initial ribosome density (ribosomes/100 nts) c
2
4/15/20
Length of the termination queue (nts)
Ribosome density at position x (nts) t c if x < d – t
2.5 ribosomes / 100 nts if x > d-t
(RD) overall
=
2 .
5 t
d
t
c d
(S2)
Density values for different ORF length were calculated from equation S2 for various values of termination queues (i.e., varying t ). The best fit to the experimental data
(minimal root mean square) occurs with a termination queue of ~ three ribosomes and a
5'-ribosome density ( c ) of 0.18 ribosomes/100 nts.
3. Density on mRNA for the slow initiation model
Total ORF length (nts)
Initial ribosome density (ribosomes/100 nts) d c
Local ribosome density is constant along the coding region and equal to the initial ribosome density. Thus the overall (or average) ribosome density for mRNA, irrespective of its length, is uniform across its coding region and equal to c .
Assuming an exponential decrease in initiation rates with length, the density of
ORF with length d (RD d
) is:
RD d
RD
0 e
-ad
(S3)
Where:
RD
0
is the density for shortest mRNA (200 nts); and a is parameter representing the magnitude of the length dependence.
The best fit of Equation S3 to the experimental data is achieved with initiation decreasing by half for every 1200 nts.
B. Estimates of processivity and termination queue from the RDM data.
3
4/15/20
The results presented in Figures 5 and 6 of the main text indicate that there is at most limited variation in ribosomal density along the coding region. This is most consistent with the last of the three models presented in section A. However, the experimental data could agree with models 1 or 2 if very high processivity levels ( r ) or very low ribosomal accumulation ( t ), respectively, were employed. Indeed, model 1 is actually the same as model 3 at 100% processivity (i.e., r = 1), and model 2 is the same as model 3 if there is no ribosomal queue at termination (i.e., t = 0).
We attempted to identify the limiting values for r and t that our experimental data can exclude in order to determine whether or not models 1 or 2 could account for the observed global inverse correlation between overall ribosome density and ORF length (1).
To accomplish this, we simulated the sedimentation profiles of fragments corresponding in sizes and position to those shown in Figures 5 and 6 using varying r (model 1) or t
(model 2), and compared the results of the simulations to the experimental results
(Supplemental Figures 1 and 2, respectively).
Simulation method
In a velocity sedimentation experiment, the 5'- and 3'-fragments are distributed among several fractions rather than just one fraction (Figures 5 and 6), presumably reflecting stochastic variation in the numbers of ribosomes bound to individual mRNA molecules. The distribution of the numbers of ribosomes on individual mRNA fragments is fit well by a Poisson distribution (see examples in Supplemental Figures 1 and 2), consistent with a random and independent positioning of ribosomes along the mRNA
(Murphy 1982). We therefore used a Poisson model to calculate predicted distributions of ribosomes on each fragment for the models of the previous section with various processivity levels ( r ) and termination queues ( t ).
The Poisson distribution for a parameter is fully determined by its average parameter (
) (Murphy 1982). We selected average values for the number of bound ribosomes on each fragment such that the sum of averages for the 5' and 3' pair of simulated fragments was equal to the sum of averages obtained experimentally.
1
In the
1 The average number of ribosomes on the fragments analyzed experimentally was calculated from the sedimentation position of the centroid of the radioactive signal and the
4
4/15/20 examples in Supplemental Figure 4, each of the two fragments analyzed experimentally
(black squares or triangles) had an average of five ribosomes, corresponding to a total of ten ribosomes on the full length mRNA. The averages for the simulations were then selected to obtain the same sum: 7.5 for the blue distribution and 2.5 for the red distribution in (A) or 5 ribosomes for both the blue and red distributions in (B).
The ratio between the averages of two simulated distributions from either ends of an mRNA is related to the processivity level ( r ) or termination queue size ( t ) (Equations
S4 and S5, respectively). average number of ribosomes on 5'-fragmen t of length a average number of ribosomes on 3'-fragmen t of length b
3 c ( r a / 3 r
0
)
100 ln r
3 c ( r d / 3 r
( d
b ) / 3
)
100 ln r
= r d / 3 r a
/ 3 r
( d
1
b ) / 3
This equation does not have an algebraic solution but can be solved numerically.
(S4) average number of ribosomes on 5'-fragmen t of length average number of ribosomes on 3'-fragmen t of length a b
=
2 .
c
5 t
100
( a b
t ) c
100
(S5)
Thus, from the ratio of the averages of the 5'- to 3'-fragments we could identify the processivity level or termination queue predicted to give that value. Vice versa, for a particular assumed processivity level or termination queue, we could identify the ratio of the averages of the 5'- and 3'-fragments and simulate their sedimentation accordingly.
Simulations were performed for various levels of processivity ( r ) or various numbers of ribosomes accumulating upstream to the stop codon, i.e. the “queue size” ( t ).
Representative simulations are shown in Supplemental Figures 1 and 2 and are described in the Supplemental Results. number of ribosomes sedimenting at that position of the gradient (determined by the
OD
254
of sedimentation profiles such as that in Figure 3B, top panel).
5
4/15/20
Supplemental Results
Processivity levels
The simulations in Supplementary Figure 1 demonstrate that relatively low processivity (99.3%, r = 0.993) gives only small overlap between the distributions of 5'- and 3'-fragments. As processivity increases the overlap between the distributions increases
(blue and red curves in Supplemental Figure 1, panels i-vi). The degree of overlap between the distributions can be measured by calculating the ratio of the amount of the 5'fragment to the amount of the 3'-fragment for each number of ribosomes (i.e., for each point along the x-axis). Plots of the logarithms of these ratios as a function of the number of ribosomes (Supplemental Figure 1, panel vii) reveal a linear relationship, which is a property of the Poisson distribution (Equation S6).
Where: ln p ( p ( x x ,
,
1
)
2
)
ln
1
2
x
(
2
1
) slope = ln
1
2
; and intercept =
2
1
(S6)
The slope of the line corresponds to the degree of overlap: small overlap of the ratios gives steep slopes, and as the overlap increases the slope become shallower. We used this measure of overlap to compare the processivity model levels to the experimental data. In panel vii of Supplemental Figure 1A-M the black squares correspond to the 5' to 3' ratios obtained experimentally. The results indicate that a processivity level of 99.8% or greater is required in all cases to obtain a reasonable agreement with the experimental data.
Termination queue
The simulations demonstrate that when many (e.g., five) ribosomes accumulate upstream to the stop codon, there is little overlap in the distributions of the 5'- and 3'fragments; as the number of ribosomes in the queue decreases (i.e. as t becomes smaller), the overlap increases (Supplemental Figure 2 panels i-v, blue and red curves). As for processivity, the degree of overlap can be determined from the slope of the logarithms of the ratios between the 5'- and 3'-fragments for each number of bound ribosomes (i.e. for
6
4/15/20 every point along the x-axis). A steep slope indicates a high accumulation upstream to the stop codon and a shallower slope indicates less accumulation of ribosomes (panel vi in
Supplemental Figure 2A-F). Comparing the 5' to 3' ratios of the experimental measurements and the simulations indicate that the RDM results are most consistent with one or less ribosomes accumulating at the termination site.
Supplemental References
Arava, Y., Y. Wang, J. D. Storey, C. L. Liu, P. O. Brown , et al.
(2003). Genome-wide analysis of mRNA translation profiles in Saccharomyces cerevisiae . Proc Natl
Acad Sci U S A 100 (7): 3889-94
Murphy, E. A. (1982) Biostatistics in Medicine (The John Hopkins University Press,
Baltimore)
Wolin S.L. and Walter P. (1988) Ribosome pausing and stacking during translation of a eukaryotic mRNA. EMBO J. 7, 3559-69
7
4/15/20
Supplemental Figure 1
Lower limits for processivity estimated by simulations of sedimentation profiles.
The sets of Panels (A-M) demonstrate graphically the predicted sedimentation profiles for individual mRNAs at various processivity levels for 5'- and 3'-fragments. The simulations are described in the Methods Supplement. The length of the fragments and their position along the coding region is depicted schematically at the top of each panel, with the 5'-fragment labeled in blue and the 3'-fragment labeled in red. Each panel includes seven graphs: i-vi) Simulated sedimentation profiles based on a Poisson model of
5'-fragment (blue line) and 3'-fragment (red line) at increasing processivity levels: (i)
99.3%;(ii) 99.6%; (iii) 99.8%; (iv) 99.9%; (v) 99.99%; and (vi) 100%. The experimental results from quantitation of Northern blots across the sedimentation gradients are shown for the 5'-fragment (black triangles) and the 3'-fragment (black squares). vii) Ratio of amount of the 5'- and 3'-fragment at each sedimentation position. For each value of number of bound ribosomes (x-axis), the ratio between the amount of 5'-fragment and 3'fragment was calculated (i.e., ratio of the blue to red values from panels i-vi). The actual ratios from experimental data are presented as black squares. Experimental measurements that were too close to background (fraction of fragment lower than 0.025) were excluded from the plot. To account for uncertainty in quantitation of the radioactive signal in the
Northern blot error bars of +50% are shown for each ratio.
8
4/15/20
Supplemental Figure 2
Length of termination queue estimated by simulations of sedimentation profiles.
The sets of panels (A-F) demonstrate graphically the expected sedimentation profiles for
5'- and 3'-fragments of a series of mRNAs when various numbers of ribosomes accumulate at maximal density (one ribosome / 40 nts) directly upstream of the stop codon. The simulations are described in the Methods Supplement. The sizes of the simulated fragments and their position along the ORF are indicated at the top of each panel. Each panel includes six graphs: i-v) Simulated sedimentation profiles based on a
Poisson model of a 5'-fragment (blue line) and a 3'-fragment (red line) with the following termination queue sizes: (i) five; (ii) three; (iii) two; (iv) one and (v) zero ribosomes. The experimental results from quantitation of Northern blots across the sedimentation gradients for the 5'-fragment (black triangles) and 3'-fragment (black squares) are also shown. vi) Ratio of amount of the 5'- and 3'-fragment at each sedimentation position. For each value of number of bound ribosomes (x-axis), the ratio between the amount of 5'fragment and 3'-fragment was calculated (i.e. ratio between the blue and red values from panels i-v). Dark green, light green, pink and dark blue lines represent the ratio when five ribosomes, three, two ribosomes and one ribosome, respectively, are at maximal density upstream to the stop codon, and the light blue line represents the ratio when there is no termination queue upstream to the stop codon. The ratios for the experimental results are presented as black squares. Error estimates and exclusion of data too close to background were set as in Supplemental Figure 1.
9
2.5
4/15/20 t
Slow termination c Slow initiation
0
0 x
Position along the mRNA d
Incomplete processivity
Supplemental Figure 3: Differences between the expected ribosomal densities along an mRNA predicted for the three models described in the text.
Each colored line represents the ribosomal density at each position ( x ) along an mRNA in the following models: Slow initiation (black dashed line): initiation is slow relative to elongation, elongation is fast and constant along the coding region with no significant dissociation, and termination is similar in rate to elongation. Incomplete processivity (red line): Rates as in slow initiation, except that ribosomes dissociate during elongation steps due to incomplete processivity. Slow termination (green line): Rates as in slow initiation except that termination is much slower than elongation, leading to a termination queue of ribosomes ( t ). The ribosome density in the termination queue is based on the ~40 nts ribosome footprint (Wolin and Walter 1988).
10
4/15/20
B)
A)
0.24
0.16
0.08
0
0
0.24
0.16
0.08
0
0
5 10 15
Number of ribosomes
5 10 15
Number of ribosomes
20
20
Supplemental Figure 4: Examples of simulated and experimental distributions of ribosomes.
The fraction of mRNA associated with each number of ribosomes is plotted for (A) 5'- and 3'-fragments with average of 7.5 ribosomes (blue line) and with average of
2.5 ribosomes (red line) and (B) fragments with average of ~5 ribosomes on each (blue and red lines almost overlap). Distributions were simulated assuming Poisson model
(Supplemental Methods). The distributions observed experimentally are plotted as black triangles and squares for 5'- and 3'-fragment, respectively, each with an average of ~5 ribosomes.
11