CHAPTER SUMMARY
advertisement

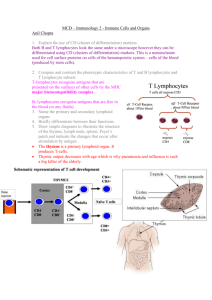
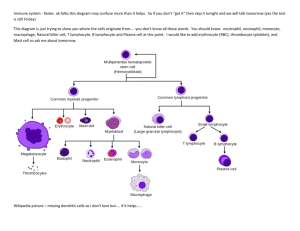
CHAPTER 10 SUMMARY Foreign invaders and newly arisen mutant cells are immediately confronted with multiple interrelated defense mechanisms aimed at destroying and eliminating anything that is not part of the normal self. These mechanisms, collectively referred to as immunity, include both innate and adaptive or acquired immune responses. The most common invaders are multicellular parasites, bacteria and viruses. In addition to defending against these microbes and mutant cells, the immune cells also clean up cellular debris, preparing the way for tissue repair. •Innate immune responses are nonspecific responses that nonselectively defend against foreign material even upon initial exposure to it. All animals have innate defenses. •Adaptive or acquired immune responses are specific responses selectively targeted against particular invaders for which the body has been specially prepared after a prior exposure. Only vertebrates with jaws are known to have adaptive immunity •Hemocytes in many invertebrates and leukocytes and their derivatives in vertebrates are the major effector cells of the immune system and are reinforced by a number of different defensive proteins. Leukocytes are produced in the bone marrow, and then circulate transiently in the blood. They spend most of their time, however, on defense missions in the tissues. Some leukocyte derivatives are also produced and differentiated and perform their defense activities within tissues strategically located at likely points of foreign infiltration. THE VERTEBRATE IMMUNE SYSTEM INNATE IMMUNITY Barrier Tissues are the first line of defense, consisting of body surfaces exposed to the outside environment—the skin and the linings of internal cavities that communicate with the external environment. These serve not only as mechanical barriers to deter would-be pathogenic invaders but also play an active role in thwarting entry of bacteria and other unwanted materials. In mammals, these systems are as follows: •The skin consists of two layers: an outer vascular, keratinized epidermis and an inner, connective tissue dermis. The epidermis contains four cell types: melanocytes, keratinocytes, Langerhans or dendritic cells, and Granstein cells. Melanocytes produce a pigment, melanin, which protects the skin by absorbing harmful ultraviolet radiation. The most abundant cells are the keratinocytes, producers of the tough keratin that forms the outer protective layer of the skin. This physical barrier discourages bacteria and other harmful environmental agents from entering the body and prevents water 10-1 and other valuable body substances from escaping. Keratinocytes further serve immunologically by secreting interleukin 1, which enhances T-cell maturation within the skin, and cathelicidins, which are antimicrobial peptides. Dendritic cells and Granstein cells also function in specific immunity by presenting antigen to helper T-cells. •The digestive system is defended by an antimicrobial salivary enzyme, destructive acidic gastric secretions, gut-associated lymphoid tissue, and harmless colonic resident flora. •The genitourinary system is protected by destructive acidic and particle-entrapping mucus secretions. •The respiratory system is defended by alveolar macrophage activity and on secretion of sticky mucus that traps debris, which is subsequently swept out by ciliary action. Other respiratory defenses include nasal hairs, which filter out large inspired particles; reflex cough and sneeze mechanisms, which expel irritant materials from the trachea and nose, respectively; and the tonsils and adenoids, which defend immunologically. Innate immune responses, which form a second line of defense against non-self cells (foreign, mutant, or injured cells) even upon first exposure to them, include (in mammals) inflammation, interferon, natural killer cells, and the complement system. •Inflammation is a nonspecific response to foreign invasion or tissue damage mediated largely by the specialized phagocytes (neutrophils and monocytes-turned-macrophages) and their secretions. The phagocytic cells destroy foreign and damaged cells both by phagocytosis and by the release of lethal chemicals. Histamine-induced vasodilation and increased permeability of local vessels at the site of invasion or injury permit enhanced delivery of more phagocytic leukocytes and inactive plasma protein precursors crucial to the inflammatory process, such as clotting factors and components of the complement system. These vascular changes are also largely responsible for the observable local manifestations of inflammation—swelling, redness, heat, and pain. •Interferon is nonspecifically released by virus-infected cells and transiently inhibits viral multiplication in other cells to which it binds. Interferon further exerts anticancer effects by slowing division and growth of tumor cells as well as by enhancing the power of killer cells. •Natural killer cells nonspecifically lyse and destroy virus-infected cells and cancer cells on first exposure to them. •Upon being activated by locally released factors or microbes themselves at the site of invasion, the complement protein system directly destroys the foreign invaders by lysing their membranes and also augments other aspects of the inflammatory process. ACQUIRED (ADAPTIVE) IMMUNITY 10-2 Following initial exposure to a microbial invader, specific components of the adaptive or acquired immune system become specially prepared to selectively attack the particular foreigner. Not only is this immune system able to recognize foreign molecules as different from self-molecules, but it can also distinguish between millions of different foreign molecules. •The cells of the adaptive immune system, the lymphocytes, are each uniquely equipped with surface membrane receptors that are able to bind lock-and-key fashion with only one specific complex foreign molecule, which is known as an antigen. The tremendous variation in antigen-detecting ability between different lymphocytes arises from the shuffling around of a few different gene segments, coupled with a high incidence of somatic mutation, during lymphocyte development. •Those lymphocytes produced by chance that are able to attack the bodies own antigen-bearing cells are eliminated or suppressed so that they are prevented from functioning. In this way, the body is able to “tolerate” (not attack) its own antigens. The major surface antigens of all nucleated cells are known as MHC molecules, which are coded for by the major histocompatibility complex (MHC), a group of genes with DNA sequences unique for each individual. There are two broad classes of adaptive immune responses: antibody-mediated immunity and cell-mediated immunity. In both instances, the ultimate outcome of a particular lymphocyte binding with a specific antigen is destruction of the antigen, but the effector cells, stimuli, and tactics involved are different. Both responses appear to be coordinated by Helper T-cells, the integrators of immune regulatory loops. •Plasma cells derived from B lymphocytes (B-cells) are responsible for antibody-mediated immunity, whereas T lymphocytes (T-cells) accomplish cell-mediated immunity. B-cells develop from a lineage of lymphocytes that originally matured within the bone marrow. The T-cell lineage arises from lymphocytes that migrated from the bone marrow to the thymus to complete their maturation. •After being activated by antigen associated with a foreign invader, a lymphocyte (either a B-cell or a T-cell, depending on the characteristics of the antigen) rapidly proliferates, producing a clonal lineage of its own kind that can specifically wage battle against the invader. •Some of the newly developed lymphocytes do not participate in the attack but become memory cells that lie in waiting, ready to launch a swifter and more forceful attack should the same foreigner ever invade the body again. •Helper T-cells activate both B-cells and effector T-cells, but these have different targets because their requirements for antigen recognition differ. •Each B-cell recognizes specific free extracellular antigen that is not associated with cell-bound self-antigens, such as that found on the surface of bacteria. Appropriately, the B-cell activated by a helper T-cell differentiates into a plasma cell, 10-3 which is specialized to secrete freely circulating antibodies that besiege the freely existing invading bacteria (or other foreign substance) that induced their production. Antibodies intensify lethal nonspecific immune mechanisms already called into play by the foreign invasion. Antibodies activate the complement system, enhance phagocytosis by acting as opsonins (tags signaling object to be destroyed by phagocytes), and stimulate killer cells. They may also kill directly via hydrogen peroxide production. •T-cells, in contrast, have a dual binding requirement of foreign antigen in association with MHC molecules on the surface of one of the body’s own cells. Two different types of host cells meet this requirement: (1) virally invaded host cells and (2) other immune cells with foreign antigen attached on their surface. •The presence of different classes of self-antigens on the surface of these foreign antigen-bearing host cells causes two different types of T-cells to differentially interact with them: (1) Cytotoxic T-cells are able to bind only with virus-infected host cells, whereupon they release toxic substances that kill the infected cell. (2) Helper T-cells can only bind with other Tcells, B-cells, and macrophages that have encountered foreign antigen. Subsequently, helper T-cells enhance the immune powers of these other effector cells by secreting specific chemical mediators. •Such differential activation of the various types of lymphocytes assures that the appropriate specific immune response ensues to dispose of the particular enemy efficiently. Moreover, B-cells, the various T-cells, and macrophages reinforce each other’s defense strategies, primarily by releasing a number of important secretory products. •In a process known as immune surveillance, natural killer cells, cytotoxic T-cells, macrophages, and the interferon that they collectively secrete normally eradicate newly arisen cancer cells before they have a chance to spread. OTHER ANIMALS Other jawed vertebrates have innate and acquired immune systems similar to those in mammals, though the genetic mechanisms of generating antibody diversity differ somewhat. Jawless vertebrates and most other animals have only innate immune defenses, which consist of: •Potent antimicrobial peptides in barrier tissues, hemolymph, and hemocytes •Opsonin proteins, which bind to foreign proteins and tag them for destruction •Phagocytes to destroy the invaders •Encapsulation enzymes and hemocytes to wall off invaders There is evidence suggesting that some type of acquired immunity exists in some arthropods. 10-4